
水稻种子休眠调控技术研究进展
2021-03-04 08:02:26邹文雄关亚静曹栋栋卞晓波施德云丁丽玲
浙江农业学报 2021年2期
邹文雄,吴 伟,关亚静,曹栋栋,卞晓波,施德云,丁丽玲
(1.浙江省种子管理总站,浙江 杭州 310020;2.浙江大学 农业与生物技术学院,浙江 杭州 310058;3.浙江省农业科学院 作物与核技术利用研究所,浙江 杭州 310021;4.金华市农业科学研究院,浙江 金华 321017;5.缙云县农业农村局,浙江 缙云321400)
种子休眠是指在一定时期内有生活力的种子在适宜发芽条件下不能发芽的现象[1]。种子休眠是植物生命周期中的一个阶段,在植物后代繁衍与进化中具有重要意义,休眠种子能够避免在恶劣环境下发芽,有助于种族延续。同时,由于休眠程度或所处环境不同,导致不同品种的种子萌发和出苗在时间和空间上存在差异,增加了同一物种不同品种生态类型的分化。
种子休眠对植物本身是一种有益的生物学特性。在温暖多湿的热带地区,种子具有易发芽的特性,休眠期很短或不存在;在干湿冷热交替且气候条件多变的温带地区,种子往往需要经过一定的休眠期才能遇到适合萌发生长的环境条件而萌发[2]。因此,种子休眠是植物适应外界环境条件以确保物种生存、繁衍和进化的自我保护方式。我国是世界上最大的稻米生产国,水稻生产地域分布广泛,生产条件和气候环境存在较大差异[3]。自1981年开始就有学者通过发芽试验测定种子休眠率和休眠期,开展水稻种子休眠特性的研究,发现水稻的遗传特性、种子内源激素含量与比值、淀粉酶活性、种子成熟期的环境条件和收获后的贮藏条件等都会影响种子的休眠特性。通过喷施不同激素、去除种子稃壳、HNO3或强氯精等溶液浸种、日晒或高温烘干等休眠调控技术可不同程度地解除水稻种子休眠,从而有效提高种子出苗率、稻谷产量和品质,更好地服务于农业生产。
1 种子休眠特性
种子休眠特性是种胚自身所固有的形态休眠(又叫形态后熟)和生理休眠(又叫生理后熟)的特性,是植物在长期的自然选择中为适应环境和自身的繁衍进化而来的,对减少种子在不利环境下萌发,确保在适宜季节和环境条件下种子的正常发芽和植株正常生长,维持种族繁衍具有重要意义。种子的遗传特性、淀粉酶活性、种被障碍、生理后熟、成熟期的环境条件和收获后的贮藏条件都会影响种子的休眠特性。
1.1 检测方法
目前一般采用原始休眠率描述种子休眠特性。水稻原始休眠率检测的方法主要有2种,一种检测方法由Wan等[4]提出:水稻抽穗35 d后取种子保存于4 ℃冰箱以保持其休眠性;将休眠性分为5级:种子出芽率(萌发种子数量占供试种子总数的百分比)在80%以上为无休眠性,发芽率在50%~80%为轻度休眠,30%~50%为中度休眠,5%~30%为强休眠,5%以下为极强休眠。该检测方法简单易行,但有的水稻品种成熟期较长,取样时种子并未完全成熟,存在检测休眠性大于实际休眠性的现象。另一种检测方法是在水稻种子收获后迅速干燥处理,测定种子发芽率;发芽率计算方法参照《农作物种子检验规程》(GB/T3453—1995)中种子生活力的测定方法,首先采用四唑染色法鉴定该批种子的生活力,然后测定发芽种子数,计算发芽率,以此判断种子休眠程度[5]。该方法能准确地反映不同生育期水稻品种种子的原始休眠特性,目前大部分人采用该方法测定种子休眠特性。笔者认为,为了更好判断种子休眠程度,应将出芽率的计算方法修订为出芽种子数占供检活种子的百分比。笔者在水稻种子休眠性调控技术研究工作中,将水稻种子休眠性分为强休眠(初次出芽率<40%)、较强休眠(40%≤初次出芽率<60%)、较弱休眠(60%≤初次出芽率<80%)、弱休眠(80%≤初次出芽率<95%)和无休眠(95%≤初次出芽率)5个等级;种子休眠性测定每隔10 d进行一次,直至达到该批种子的最高出芽率为止(出芽率与该批种子本身的生活力基本相一致),种子休眠持续的时间即为该水稻品种的休眠期。该检测方法进一步量化了水稻休眠程度,有利于充分反映水稻品种休眠特性的真实情况。
1.2 遗传特性
种子的休眠特性是由多基因决定的,种子的休眠现象普遍存在,但不同作物休眠程度不同,例如大麦种子的休眠性通常比水稻和玉米强[2]。同一作物不同类型和不同品种的种子休眠性也会相差悬殊。如大部分籼稻休眠性很弱,充分成熟的籼稻种子,经收获干燥后,发芽率即可达80%以上;多数早粳稻的休眠性较弱,其休眠期约为1~4周;多数晚粳稻品种休眠性较强,休眠期长的可达数月;水稻恢复系的休眠性一般强于不育系。随着人们对稻米食用品质和产量的更加重视,籼粳杂交稻发展迅速,但籼粳杂交稻品种存在较严重的穗发芽问题[5]。曹雅君等[6]研究表明,不同休眠性水稻品种杂交后代具有不同程度的休眠性,不同休眠性基因位点互作会影响F1代种子休眠特性;另外,强休眠性水稻品种与无休眠性品种的杂交后代休眠性介于两亲本之间,为中度休眠;轻度休眠性品种与无休眠性品种杂交后代,有保持轻度休眠性的,也有表现强休眠性的,这可能是不同休眠性基因位点间的互作引起的。同一品种不同成熟度种子的休眠性强弱不同,随着种子后熟,DNA和RNA的活性逐渐提高,种子休眠程度减轻。常志远等[7]发现,秀水121等26个中度休眠性水稻品种的休眠期均在60 d以上,可见中度休眠品种因休眠解除速率低也会具有较长休眠期[8]。有研究发现,籽粒大小、颖壳厚度和种皮颜色等遗传特性均对种子的休眠性有不同程度的影响[9]。因此,在杂交水稻制种工作中,选用抗穗萌发或穗萌发程度较轻的不育系是从遗传上解决水稻制种发生穗发芽的根本措施。
1.3 内源激素
近年来,越来越多的研究表明,种子休眠可能取决于赤霉素(GA)和脱落酸(ABA)生物合成和分解代谢的内在平衡,而不取决于这2种激素的绝对含量[10]。周述波等[11]研究表明,ABA在抑制种子萌发过程中α-淀粉酶活性、可溶性糖、可溶性蛋白含量和ABA含量增加,从而延缓种子萌发。细胞分裂素(CTK)、吲哚乙酸(IAA)等激素在调控种子休眠方面也起非常重要的作用。刘杨[12]研究表明,内源CTK和IAA参与水稻种子萌发与休眠的转换,其中CTK是直接因子,IAA是间接因子,休眠调控不仅受到CTK和IAA绝对含量的影响,还受到CTK与IAA相对含量的影响。此外,CTK还可通过调控水稻分蘖芽中的OsD3(水稻抑制分蘖基因)、OsD10(水稻抑制分蘖基因)和OsD27(水稻控制分蘖基因)等基因表达,调控水稻植株分蘖芽生长。因此,可以通过测定内源激素含量来预测水稻种子休眠特性强弱。
1.4 淀粉酶活性
淀粉酶是种子萌发初期降解淀粉的关键酶,胚乳中的淀粉在淀粉酶的作用下转化为糖,然后运往种胚的生长部位,为胚提供构建新组织的底物和能量。淀粉酶活性的提高是种子尤其淀粉类种子发芽的主要生理特征之一。秦岭等[13]研究结果表明,板栗种子萌发前淀粉酶活性上升,淀粉水解加速。张桂莲等[14]指出无休眠或弱休眠水稻种子中的淀粉酶含量显著高于强休眠水稻种子。王丽萍等[15]研究表明,水稻淀粉酶活性与种子的休眠特性强弱呈负相关。廖泳祥等[16]研究表明,内源GA相对含量和淀粉酶活性较高是穗发芽的主要生理特征。因此,淀粉酶活性可作为衡量水稻种子休眠特性强弱的一个重要生理指标。
2 水稻种子休眠机理
2.1 遗传学基础
国际上,水稻种子休眠调控研究、抗穗发芽的种质资源筛选工作始于20世纪50年代[17]。胡伟民等[18]认为,水稻种子休眠是数量性状的遗传特性,受微效多基因控制。通过定位克隆休眠特性的数量性状基因座(QTL),培育具有不同休眠性的水稻新品种,能有效避免水稻生产中穗发芽现象的发生。学者们通过构建水稻分子标记和遗传连锁图谱,利用水稻染色体重组自交系(RIL)、染色体片段代换系、双单倍体(DH)、F2世代等分离群体对种子休眠相关性状进行QTL遗传分析[19]。由于种子休眠的复杂性,种子休眠遗传特性研究进展非常缓慢,目前共定位了200多个与水稻种子休眠性状和穗发芽性状相关的QTL基因位点。这些QTL广泛分布于水稻的12条染色体上。在1~8、11号染色体上检测到的QTL数量相对较多,在9、10、12号染色体上分布的QTL数量相对较少(表1)。在这些检测到的QTLS中,已精细定位的只有3个,分别是位于第3号染色体的sdr1、第7号染色体的sdr4、第12号染体的qsd12。
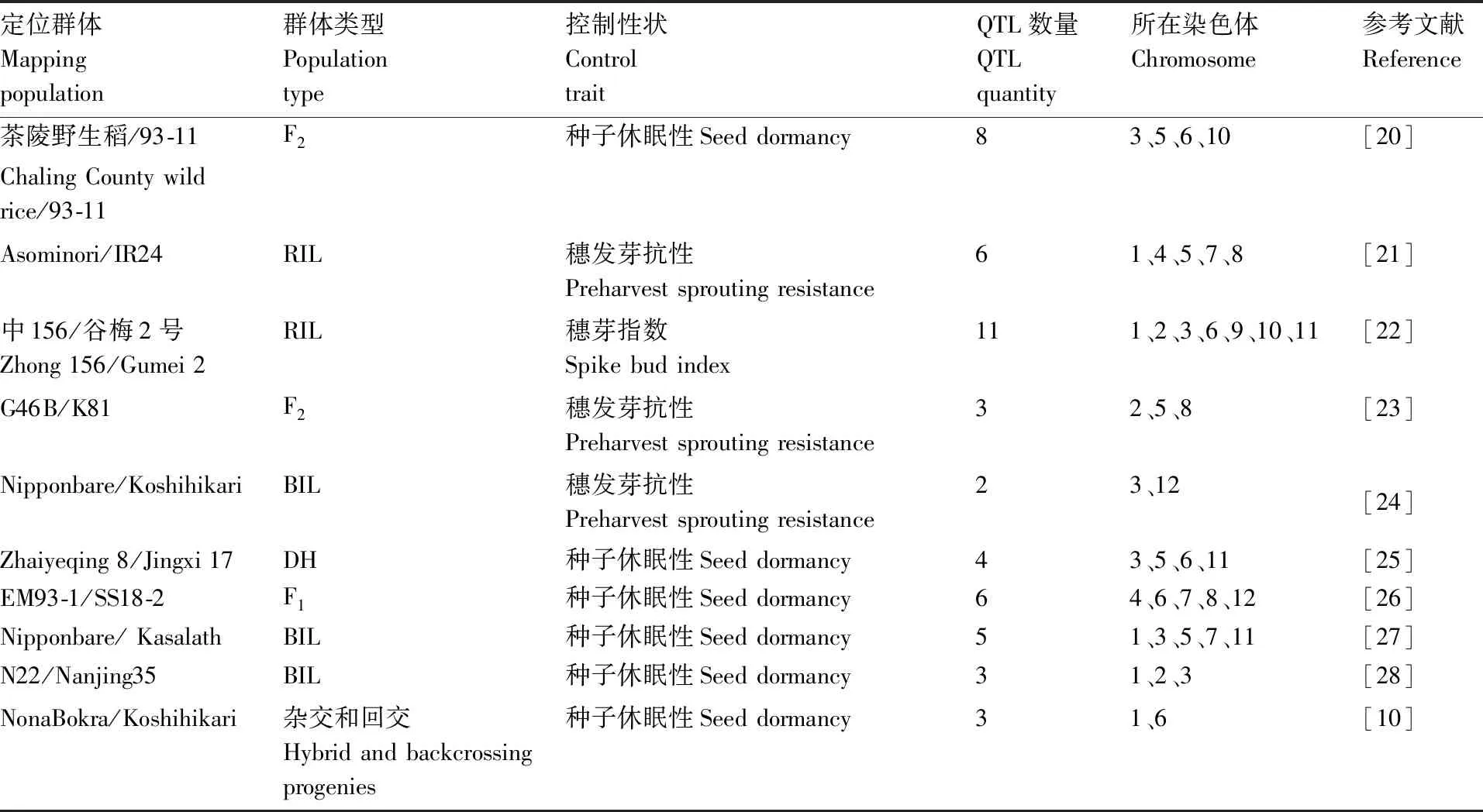
表1 部分与水稻种子休眠或抗穗发芽性状相关的QTL基因位点Table 1 Some QTLs for traits associated with seed dormancy or pre-harvest sprouting resistance in rice
Lin等[29]于1998年在第8条染色体末端检测到控制种子休眠性状的QTL,与Dong等[21]于2003年检测到控制穗发芽的QTL位置相同。唐九友等[30]于2004年利用DV85/KinmazeRIL群体检测到第5条染色体上的休眠性QTL与Lin等[29]于1998年检测到的QTL位点相一致。在不同年份、不同定位群体中检测到位置基本一致的控制休眠性状的QTL,表明这些QTL在调控水稻种子休眠性状方面是稳定表达的。孙爱伶等[31]研究表明,水稻种子携带的休眠性QTL位点越多,其休眠性越强。Miao等[32]研究发现,将产量调节因子miR156敲除,可去除其对靶基因IPA1(ideal plant architecture 1)的抑制作用,从而影响GA代谢通路,增强种子休眠性。Jung等[33]敲除调节水稻种子发育和ABA信号传导的关键转录因子viviparous-1(OsVP1)后发现,水稻种子休眠性下降。Chen等[34]认为,表型遗传调控因子Polycomb复合物蛋白(PcG)基因OsEMF2b可直接与OsVP1的启动子结合,间接或直接调控OsVP1表达,参与ABA信号通路,在种子休眠和幼苗生长过程中发挥关键作用。研究穗发芽和种子休眠相关的QTL,可以更深层次地了解水稻种子休眠的调控途径,为农业生产中调控种子休眠提供理论依据和技术支撑。
2.2 代谢途径
糖酵解(EMP)、三羧酸循环(TCA)和磷酸戊糖(PPP)等呼吸代谢途径都与种子发芽有关。Major等[35]首次提出PPP途径是解除种子休眠的主要途径。糖代谢的PPP途径可以产生DNA和RNA的前体物质,这些物质是种子发芽所必需的。Ogawa等[2]认为,种子休眠的破除是从EMP途径(包括TCA)转向PPP途径的结果,并指出休眠的破除需要伴随氧化反应,还原型辅酶(NADPH)从还原态转化为氧化型辅酶(NADP)。Han等[36]研究水稻种子萌发过程中种胚组织磷酸化调控代谢发现,磷酸化可促进细胞壁前体合成,加快水稻种子形态后熟;信号转导蛋白的磷酸化主要在萌发早期被激活,而胁迫反应和贮藏蛋白的磷酸化则在种子萌发后期被增强;酶活性测定结果发现,果糖激酶、丙酮酸激酶、苹果酸脱氢酶、抗坏血酸过氧化物酶和谷胱甘肽S-转移酶的磷酸化均能持续提高种胚组织活性。PPP途径参与调控水稻种胚组织的淀粉降解,为水稻种子萌发提供所需能量(图1)。Ye等[37]研究表明,与水稻胚乳休眠和株高相关的基因qSD1-2可能通过GA调节的脱水机制控制种胚的初级休眠,种胚组织通过磷酸化进行的蛋白质修饰在控制种子休眠中起关键作用。
王丽萍等[15]分析28个核不育系组配的水稻品种得出,α-淀粉酶活性和可溶性蛋白含量皆与种子休眠性呈负相关,可作为衡量种子休眠特性强弱的重要生理指标。淀粉酶促使籽粒内部贮藏的淀粉提前水解,导致可溶性糖等与呼吸有关的物质含量增加,利于种胚萌发。张桂莲等[14]发现,水稻种子中α-淀粉酶活性提高是造成种子穗发芽的重要原因。Xu等[38]研究发现,东乡野生稻(DXWR)种子的休眠和胚乳贮藏蛋白的降解密切相关,DXWR种子萌发时胚胎中参与EMP、TCA、细胞生长和分裂、蛋白质合成与信号转导的蛋白质相对含量比萌发前高,胚乳中蛋白质相对含量则保持不变或减少。
2.3 激素调控
GA、细胞分裂素(CK)和ABA的相互作用可以调节种子的萌发和休眠[39]。ABA维持种子休眠、抑制种子萌发,而GA则促进种子萌发[40]。
CK可减轻或消除ABA对种子萌发的抑制作用。胡晋[41]认为,在低温条件下,GA处理能打破种子休眠,提高其发芽能力。Tylewicz等[42]发现,脱落酸通过胞间连丝阻止细胞间通讯,抑制分生组织生长,促进水稻初生芽休眠。在水稻种子发育后期,一些胚胎晚期发育LEA蛋白基因特异性表达,其中LEA33蛋白基因的突变可能通过减少油菜素内酯(BR)的积累和增强GA的生物合成来影响种子发芽[43]。
外施GA(50~110 mg·L-1)可解除种子休眠,促进种子萌发。如果GA与100 mg·L-1激动素(BA)配合使用,促进种子萌发效果更加显著。周述波等[11]发现,110 mg·L-1GA破除水稻种子休眠效果不如80 mg·L-1GA。在杂交水稻种子生产过程中,田间喷施GA对解除不育系抽穗包颈现象、改善父母本异交态势具有显著效果,并能改良不育系开花习性,提高柱头外露率,增强柱头活力,大幅度提高水稻制种产量;但GA浓度过高易引发水稻穗发芽[44]。随着穗发芽机理研究的不断深入,利用外源ABA来控制穗发芽越来越被人们所了解[45]。但是,由于采用外施ABA控制田间稻穗发芽成本较高,在农业生产实际中仍以使用可代替ABA的、有效的、绿色的穗萌抑制剂为主。
2.4 环境因素
田间温度、湿度、光照、氧气等都会影响种子休眠。刘英等[46]研究表明,低温层积处理(2 ℃,15~35 d)能够提高野慈姑种子的发芽率,是因为低温层积处理后的种子种皮被软化,种子透水通气性增加,使种子萌发所需的酶、激素、可溶代谢物质和其他化合物达到足够水平,促使种子逐渐由休眠态转向萌发状态。大部分禾谷类作物种子播前晒种可解除休眠,其主要原因是高温晒种可提高种被的通气性。张会杰[47]对161个晚粳稻品种进行耐穗发芽初筛试验,发现水稻种子成熟时期温度越高休眠期越短。在缺氧的情况下,即使种子处于能够萌发的状态下,也会被迫进入休眠而难以萌发,这种现象称为二次休眠。光照主要影响种子内光信号受体的光敏色素,远红光吸收型(Pfr)和红光吸收型(Pr)是光敏色素的2种存在形式。对拟南芥进行光诱导后发现,光照使Pfr与Pr相互转换,Pfr与Pr比值低时可以抑制种子发芽,反之则会促进种子发芽[48]。光温敏核不育系培矮64S海南春季繁殖种子的休眠期较湖南夏季繁殖的休眠期长15 d,表明光照和温度均会影响种子休眠性[48]。种子生长发育过程中的环境因素和收获后贮藏条件会对种子休眠产生影响,了解种子休眠与环境因子的内在联系,可以有效地调控种子休眠。
3 水稻种子休眠调控技术
3.1 物理调控技术
3.1.1 种植管理
顾德兴等[49]研究表明,种子的休眠特性既有遗传基因的作用又受环境因素的影响,不同群体在同一遗传背景下由于所处生态环境不同导致休眠程度也存在差异。在水稻种子灌浆成熟期,调控田间温湿度是防止或减轻穗发芽的重要途径之一。休眠性较弱的水稻品种需适当稀植,并在灌浆成熟时期关注田间温湿度,湿度尤为重要。在选择水稻种子生产基地和安排季节时尽量将种子抽穗扬花期、灌浆成熟期与当地多雨季节避开,同时增施磷钾肥和硅锌肥,防止增施氮肥导致水稻贪青、倒伏。在水稻授粉结束后应及时进行开沟排水,降低水稻田间湿度。杂交稻制繁种授粉结束后尽早将父本割除,减少制种田种植密度,增加制种田通风透光性,从而有效降低制种田水稻穗层湿度。水稻进入成熟期后一旦发现有穗发芽现象,要在85%谷粒成熟情况下,抢晴天收割有穗发芽的田块,收获后及时干燥[50]。在现有的技术条件下,通过栽培技术预防穗发芽是有效的措施之一,且成本相对较低。
3.1.2 其他物理调控技术
去除水稻种子稃壳也是解除其休眠的有效措施。水稻属于典型的颖果,休眠的主要原因是糙米粒被稃壳包裹,种子发芽时缺乏氧气。胡晋[41]认为,水稻去除稃壳能改变种皮对水和气的通透性,从而使种子快速发芽。王丽萍等[15]研究发现,去掉稃壳后,水稻种子的吸水速率和呼吸速率都显著升高,种子发芽率也相应提高。许旭明[51]认为,水稻新鲜颖壳内表面存在种子发芽抑制物质,包括ABA、有机酸类、脂类、酚类、苯类、醛类、醇类和酮类等,会抑制种子正常发芽;这些抑制物质在种子后熟过程中含量逐渐降低,故可去除颖壳解除休眠。但在农业生产实际中,批量水稻种子采用机械去壳,极易损伤种胚而造成种子发芽率等指标下降。同时,去壳的水稻种子在发芽过程中容易感染病菌,一方面可能是由于稻谷稃壳中存在抑菌物质,另一方面稻谷在机械去壳时造成病菌的交叉感染,引起种子感病率大幅增加。
对于休眠性较强的水稻种子,目前最常用的破除种子休眠的技术是高温干燥法,即在36~41 ℃烘干水稻种子,使粳稻种子含水量降至14.0%左右,籼稻种子含水量降至12.5%左右。高温干燥能破除种子休眠,可能是因为增加了水稻种被的透气性。戴剑等[52]认为日晒可提高种子发芽率,晒种2 d以上,可有效破除种子休眠;同时发现,部分水稻品种需要高温催芽,提高种子发芽时的温度能显著提高发芽率。白嵩等[53]则认为,30℃/20℃昼夜处理可提高淀粉酶类和脱氢酶类物质的活性,有利于打破种子休眠,并显著增强水稻秧苗素质,促进水稻秧苗根系发育。
3.2 化学调控技术
3.2.1 抗穗发芽药剂
杂交水稻的穗发芽问题一直是水稻制种工作中的难题,杂交水稻由于母本种子裂颖性、南繁北育对浅休眠品种的人工选择、籽粒灌浆期遭遇高温多雨、田间喷施赤霉素等因素导致长江中下游地区杂交水稻制种穗发芽现象经常出现,水稻种子生产因穗发芽导致质量不合格,对种子企业造成较大的经济损失。植物激素在种子休眠的形成、维持和解除过程中都起着重要的作用。ABA调控种子休眠的作用与GA相反,ABA促进种子储藏蛋白和脂肪的合成,导致种子休眠,抑制种子发芽;GA启动和促进种子萌发,80 mg·L-1GA破休眠效果最佳。周述波等[11]研究表明,杂交水稻在制种过程中存在GA残留的问题,因此,在杂交稻制种过程中应尽可能减少GA的用量。另外,有研究表明,种子休眠调控与GA、ABA的绝对含量关系不大,但与两者的相对含量密切相关。郭根源[54]对环己胺、脱落酸、烯效唑、丁香酚、马来酰肼、烯效唑、多效唑等不同激素和药剂组合进行筛选,筛选出3种效果较好的水稻穗芽抑制剂组合;在水稻灌浆结实期喷施3种抑制剂组合发现,3种抑制剂组合分别使测试品种的种子穗芽率降低了46%、47%和54%,且对测试水稻种子千粒重均无显著影响。Bewley[1]认为,外施高浓度GA(10-4mol·L-1)可解除水稻种子休眠。综上可知,合理运用ABA和GA等植物激素可以有效调控水稻种子休眠。
3.2.2 破休眠药剂
种子休眠对加速育种进程和早翻早(正季早稻秋繁)种子不利,使种子检验工作也难以取得正确结论[51]。采用强氯精、H2O2、硝酸等稀释溶液对水稻种子进行浸种处理,可以有效地解除种子休眠。彭丁文等[55]通过混合液1号(H2O2+强氯精)和混合液2号(GA+强氯精)破除种子休眠,发现混合液1号和2号解除休眠的效果与单独使用H2O2溶液效果相似,可能是因为H2O2溶液、GA与强氯精存在一定的相互削弱作用。刘浪等[56]研究发现,5%的H2O2溶液和0.25g·L-1的GA均能打破水稻种子休眠,且效果较其他浓度好;此外,强氯精溶液对解除种子的休眠有极显著作用,且以稀释浓度1∶125处理效果最佳。丁显萍等[57]通过试验证明,0.1%的GA溶液处理水稻种子,打破种子休眠效果较好。GA可诱导水稻种胚的盾片和糊粉层组织合成和分泌α淀粉酶,诱导水稻种胚周围组织软化,克服种皮对种胚的束缚,促进种胚生长,使水稻种子出苗整齐、秧苗长势强壮。
3.3 化学调控技术的应用
3.3.1 抗穗发芽
胡晋[41]发现,在水稻乳熟末期至黄熟初期喷施1~2 g·kg-1的丁香酚可有效抑制穗萌发,且该方法对环境无污染。此外,多效唑也能抑制水稻种子穗上发芽,随着多效唑用量的增加,田间稻穗上的种子穗发芽情况明显降低;一般抽穗后15 d叶面喷施750 g·hm-2多效唑1~2次,抗穗发芽效果显著。王熹等[58]发现,外源ABA与马来酰肼对杂交稻和常规稻均有抑制穗发芽的效果。雍太文等[59]研究表明,在种子发育各时期喷施ABA均能显著降低冈46A的穗芽率,同时提高结实率和千粒重,其中,以乳熟末期喷施50 mg·L-1ABA效果最佳。丁香酚、多效唑、ABA、马来酰肼等属于无毒的水稻穗发芽抑制剂,选用无毒的穗发芽抑制剂是解决水稻种子生产穗发芽问题的有效途径之一。
3.3.2 破除休眠
徐伟东等[60]研究表明,休眠水稻种子采用稀释400倍的强氯精溶液浸种6 h可最大幅度地破除其休眠,但浸种时间不宜超过12 h;强氯精同时具有打破种子休眠和杀灭种传病菌作用,适合在农业生产中大批量水稻种子破休眠时应用。许旭明[51]研究表明,休眠性深的水稻品种采用29% H2O2溶液浸种2 h,休眠浅的品种采用3% H2O2溶液浸种24 h,可有效解除水稻种子休眠。硝酸对破除水稻种子休眠也有良好的效果,硝酸可以腐蚀水稻种皮,使种皮通透性增加,且硝酸价格低廉。广西省农业科学院热带作物品种资源研究所采用0.1 mg·L-1硝酸或1.2×10-4mg·L-1赤霉素溶液浸种24 h,可以将具有强休眠性的野生稻种子发芽率从5%提高到80%以上。但低浓度的硝酸依然具有很强的氧化性,在使用过程中应格外小心[51]。农业生产上大批量处理具有休眠性的水稻种子适宜采用强氯精溶液浸种处理,科研育种、原种繁殖所需小批量水稻种子破休眠处理可采用H2O2或硝酸溶液;但H2O2和硝酸溶液均具有腐蚀性,需由专业人员操作,注意人身安全。
4 展望
水稻种子休眠是在生命周期中一个十分复杂的现象,受多基因调控和环境因子的影响。目前,利用数量遗传学方法和基因编辑等手段对其休眠特性进行了较为深入的研究,但由于使用的供体品种不同,如中156/谷梅2号等定位群体控制穗芽指数的QTL及其染色体分布存在明显差异,并且由于不同的研究者所使用的分离群体不同,使得对已有水稻种子休眠性QTL的定位及其效应表达情况的有效性难以进行比较[22]。此外,种子成熟环境与贮藏条件等的差异会导致同一品种休眠特性的改变,这对种子休眠机制的探索造成一定的困难;同时,需要在农业生产中根据实际情况采用相应的休眠调控技术措施,但这方面还缺乏系统性研究。通过对水稻QTL与环境互作效应分析,探讨水稻抽穗期温度对种子休眠性的影响,筛选出优异材料和QTL基因位点,可为今后利用分子标记辅助选择(MAS)技术选育抗穗发芽品种提供基础。
水稻是我国的主要粮食作物之一,确保水稻种子质量是我国粮食安全的重要保障。对水稻种子休眠机制与调控机理进行研究在较大程度上可为减少穗发芽、提高制繁种的种子质量,或破除种子休眠促进田间出苗快速整齐、确保不误农时、保证粮食作物产品产量与品质等提供理论指导与技术支撑。随着分子生物学技术的快速发展,以及水稻休眠相关基因的深入挖掘,水稻休眠特性的精准检测和优异种质资源的创新将进一步加快。对水稻种子休眠性调控将不再只关注某个单一技术,而是更多地强调多种技术相结合的系统性精准调控,比如水稻种子休眠机理的深入研究,抗穗发芽水稻品种的选育,栽培管理措施的科学研究,水稻品种休眠特性数据库的建立,高效绿色低成本易施用的水稻穗萌抑制剂研发,快速有效水稻种子休眠破除技术体系构建等等,以满足我国乃至世界水稻生产的发展需要,而这些都需要大量的系统性科学研究支撑。本文作者所在研究团队初步建立了浙江省水稻主栽品种种子休眠特性数据库,并且综合休眠率和休眠期将水稻品种的休眠特性划分为5个等级;根据不同的水稻品种休眠特性,提出了破除休眠的适宜处理方法,以提高田间出苗整齐度。此外,经多年研究筛选,开发出绿色有效的水稻抗穗萌药剂及其配套施用技术,以尽可能降低水稻种子生产基地出现穗上发芽现象,提高水稻种子生产质量;同时还积极探索采用种植栽培技术预防水稻制繁种穗发芽。但是这方面研究还处于初级阶段,尚需要更为深入的系统性研究。
猜你喜欢
防护林科技(2020年11期)2020-12-30 03:55:36
学苑创造·A版(2020年12期)2020-01-07 14:07:23
中国外汇(2019年15期)2019-10-14 01:00:34
中国粮油学报(2019年4期)2019-07-12 09:06:32
中国食品学报(2019年3期)2019-01-13 01:47:39
作文教学研究(2016年1期)2016-07-05 12:22:47
蔬菜(2016年10期)2016-03-27 12:35:11
武夷学院学报(2015年3期)2015-07-18 11:03:47
医学研究杂志(2015年8期)2015-06-22 14:00:57
食品工业科技(2014年15期)2014-03-11 18:17:52
