
山药不同种植模式对土壤线虫群落结构和土壤理化性质的影响
2021-03-04 08:02范琳娟刘子荣徐雪亮王奋山彭德良姚英娟
浙江农业学报 2021年2期
范琳娟,刘子荣,徐雪亮,王奋山,彭德良,姚英娟,*
(1.江西省农业科学院 农业应用微生物研究所,江西 南昌330200;2.中国农业科学院 植物保护研究所,北京 100094)
山药是重要的药食同源优势作物,综合种植效益远高于其他粮食作物和经济作物[1]。但由于单一的种植结构和种植特点,以及长期不变的种植茬口和品种,长期种植山药的土壤环境逐渐劣变,土传病害发生逐年加重[2],尤其以植物寄生线虫的发生最为严重。研究发现,为害山药的病原线虫主要为短体线虫和根结线虫[3-6]。据调查,其在江西省的田间发生率一般为10%~50%,重者可达100%,可致减产20%左右。作者团队于2019年对江西省山药主产区植物寄生线虫的发生程度进行调查,结果发现,大部分山药块茎和土样中都检出了根结线虫或短体线虫,山药出现明显的连作障碍。山药病原线虫的繁殖能力和抗逆性都很强,化学防治比较困难。为此,农户常采用新开垦荒地栽培或两茬间撂荒3~10 a方才再次栽培的方式,这严重制约了山药种植面积的扩大和山药产业的发展。
土壤线虫是土壤生态系统的重要构成部分,按照食性的不同,可分为自由生活线虫和植物寄生线虫2类。自由生活线虫对有机物分解、养分循环和土壤肥力保持等具有重要作用[7-9];植物寄生线虫主要寄生于植物的各种组织,对植物产生危害。由于土壤线虫具有数量多、营养类群多样、易于鉴别、世代周期短、对环境变化敏感等特点,能及时反映土壤环境的变化,因此也常被用作土壤生态系统受干扰的指示生物[10-12]。
杜峰等[13]研究表明,不同撂荒年限会对土壤生态环境,尤其是对土壤养分产生影响。但在本研究检索范围内,并未见到以土壤线虫为指示生物研究山药两茬间不同撂荒年限对土壤生态环境影响的报道,而且,关于撂荒对控制山药病原线虫种群数量的效果也未得到有效研究。为此,特选择山药线虫病发生较重的瑞昌山药栽培试验区,采取两茬间隔0、3、6、10 a等4种种植模式,通过研究其对土壤线虫群落结构的影响,评价不同种植模式下山药田的土壤状况,以期为通过改变种植模式来防治山药线虫病提供依据。
1 材料与方法
1.1 供试材料和试验设计
供试土壤采自瑞昌市高丰镇青丰村瑞昌山药田(29°35′N,115°33′E)。试验共设计4个处理:YPI-0,两茬不间隔,2018年和2019年连续种植山药2 a;YPI-3,两茬间隔3 a,2016年和2019年各种植山药1 a,2017年和2018年无作物种植;YPI-6,两茬间隔6 a,2013年和2019年各种植山药1 a,2014—2018年间连续5 a无作物种植;YPI-10,两茬间隔10 a,2009年和2019年各种植山药1 a,2010—2018年间连续9 a无作物种植。试验田面积为20 m2(4 m×5 m),株行距(20~30) cm×(45~55) cm,每小区种8行,每行20株,即每小区160株,每处理重复3次。不同处理的田间管理方式等基本一致。
1.2 试验方法
分别在山药苗期(2019-07-18)和成熟期(2019-11-20)2次在田间采样。第一次采样是在降雨后一周进行的,土壤含水量在20%左右;第二次采样与第一次采样期间降雨极少,至第二次采样时土壤含水量在10%左右。采用随机五点取样法,取5~20 cm深度土样,每个取样点取约200 g,混匀后取约500 g土样带回实验室。待测土样一部分用于分离土壤线虫,一部分自然风干后,用于测定含水量和土壤理化性状。
1.2.1 土壤理化性状测定
土壤pH值,采用水土体积质量比2.5∶1浸提-酸度计法测定;有机质含量,采用重铬酸钾容量法测定;铵态氮含量,采用靛酚蓝比色法测定;速效钾含量,采用火焰光度法测定;速效磷含量,采用钼锑抗比色法测定[14]。
1.2.2 土壤线虫的分离与鉴定
采用贝尔曼漏斗法分离线虫48 h,60 ℃杀死分离到的线虫后,在显微镜下观察线虫的形态特征,依据谢辉[15]的《植物线虫分类学》进行属水平的鉴定,分别统计各属线虫的数量,以及4大营养类群——食细菌(bacterivores)、食真菌(fungivores)、杂食/捕食(omnivores/predators)和植物寄生(plant-parasites)线虫的数量[16]。
1.3 数据处理
(1)多样性指数。参照文献[17-18]的方法,计算Shannon多样性指数(H’)和优势度指数(λ)。
(2)生态功能指数。参照文献[19]的方法计算线虫通路比值(NCR)和瓦斯乐斯卡指数(WI)。
(3)成熟度指数。参照文献[20]的方法,计算植物寄生线虫成熟度指数(PPI)和自由生活线虫成熟度指数(MI)。
参考Bongers[20],计算c-p值,根据线虫不同的生活史策略,将陆地和淡水生活的线虫划分为r-对策者向K-对策者过渡的5个类群,分别对应于1~5。
所有数据用Microsoft Excel 2013进行整理,用SPSS 20.0做统计分析。采用冗余分析(RDA)研究不同处理的土壤线虫群落组成与环境变量间的关系,变量的显著性需经过499次的蒙特卡罗(Monte Carlo)检验,用Canoco 4.5软件制作RDA排序图。
2 结果与分析
2.1 不同种植模式对土壤理化性质的影响
土壤理化性质与山药的生长状况密切相关。由表1可知,不同种植模式对山药苗期和成熟期的土壤pH值、有机质和速效钾含量,以及成熟期的土壤铵态氮含量无显著影响。苗期时,YPI-10处理的土壤铵态氮含量显著(P<0.05)高于其他处理。YPI-0和YPI-3处理的土壤速效磷含量在山药苗期和成熟期均显著(P<0.05)高于YPI-6和YPI-10处理。与苗期时相比,成熟期时不同处理的土壤pH值,以及土壤有机质、铵态氮和速效磷含量均降低,而速效钾含量大幅提高。
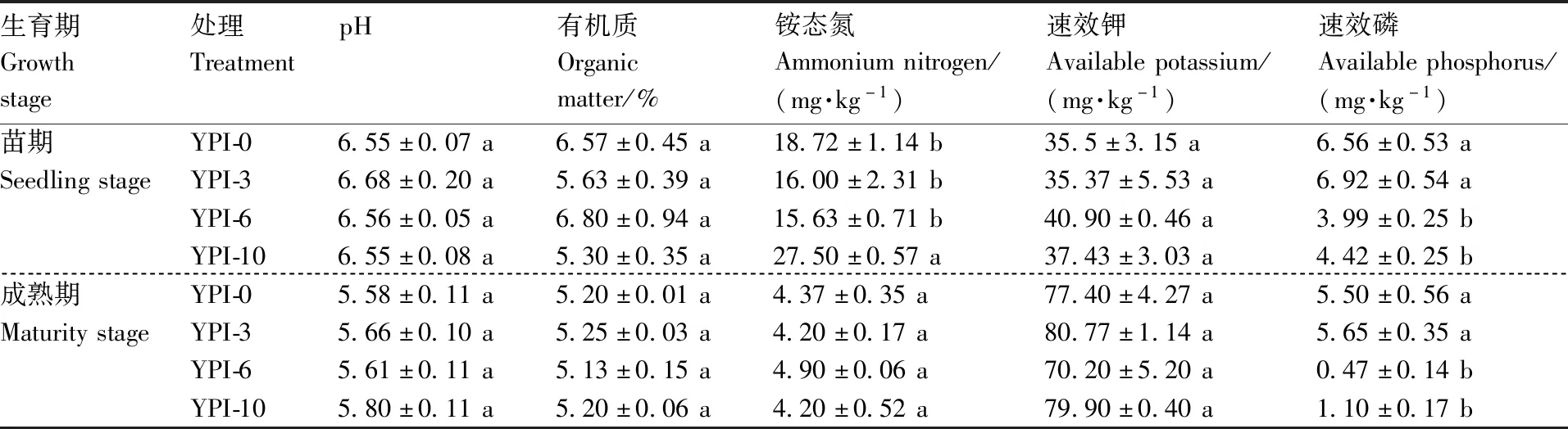
表1 不同种植模式的土壤理化性质Table 1 Soil properties under different planting patterns
2.2 不同种植模式对土壤线虫群落组成的影响
本研究共从山药土壤中分离、鉴定出线虫26个属(表2),其中从苗期土壤中分离鉴定到的有23属,每100 g干土中线虫的数量在500~700条(表3),从成熟期土壤中分离鉴定到的有18属,每100 g干土中线虫的数量在200~300条。
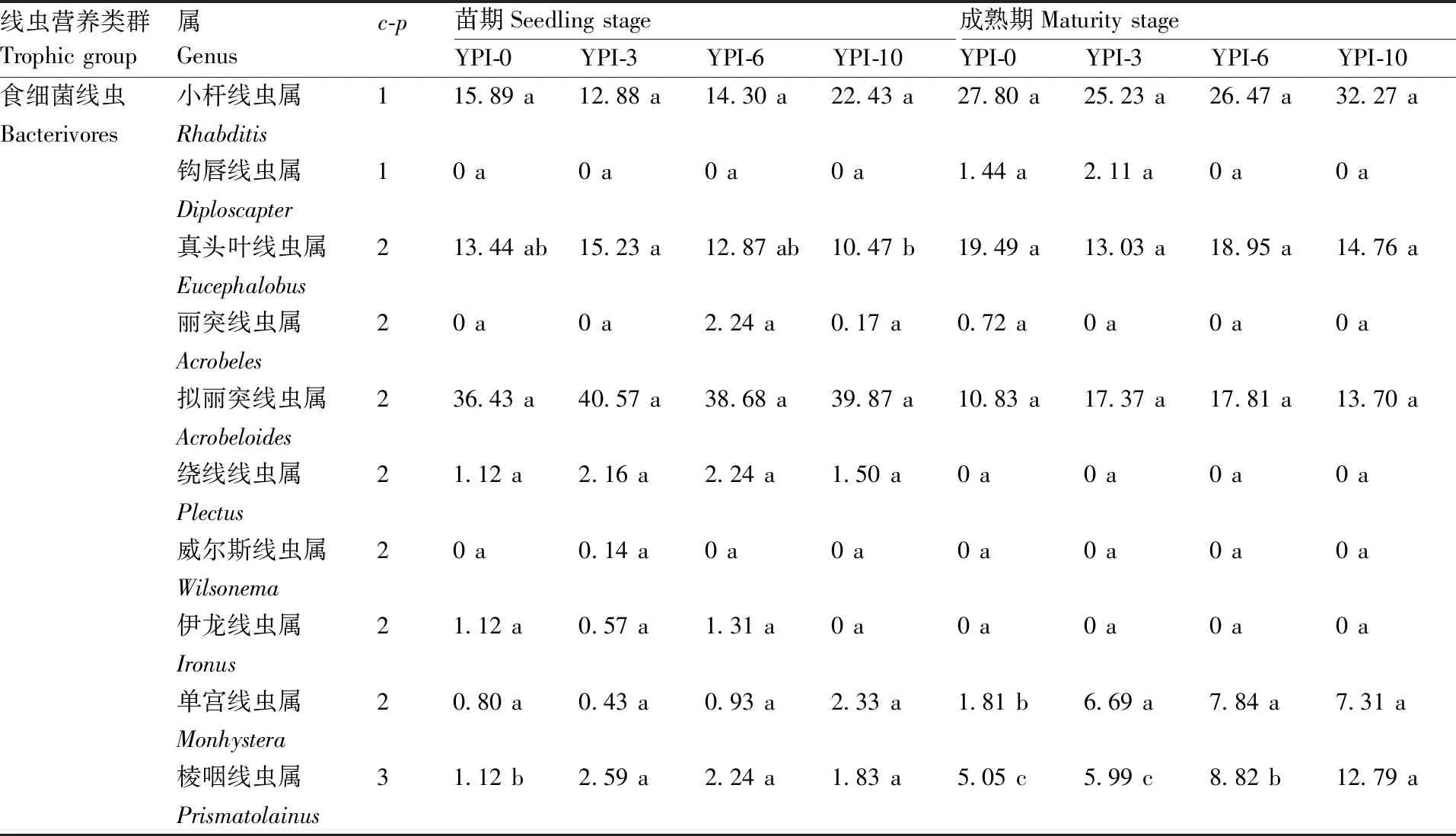
表2 不同种植模式对土壤线虫群落相对丰度的影响Table 2 Effects of different planting patterns on relative abundance of soil nematodes %
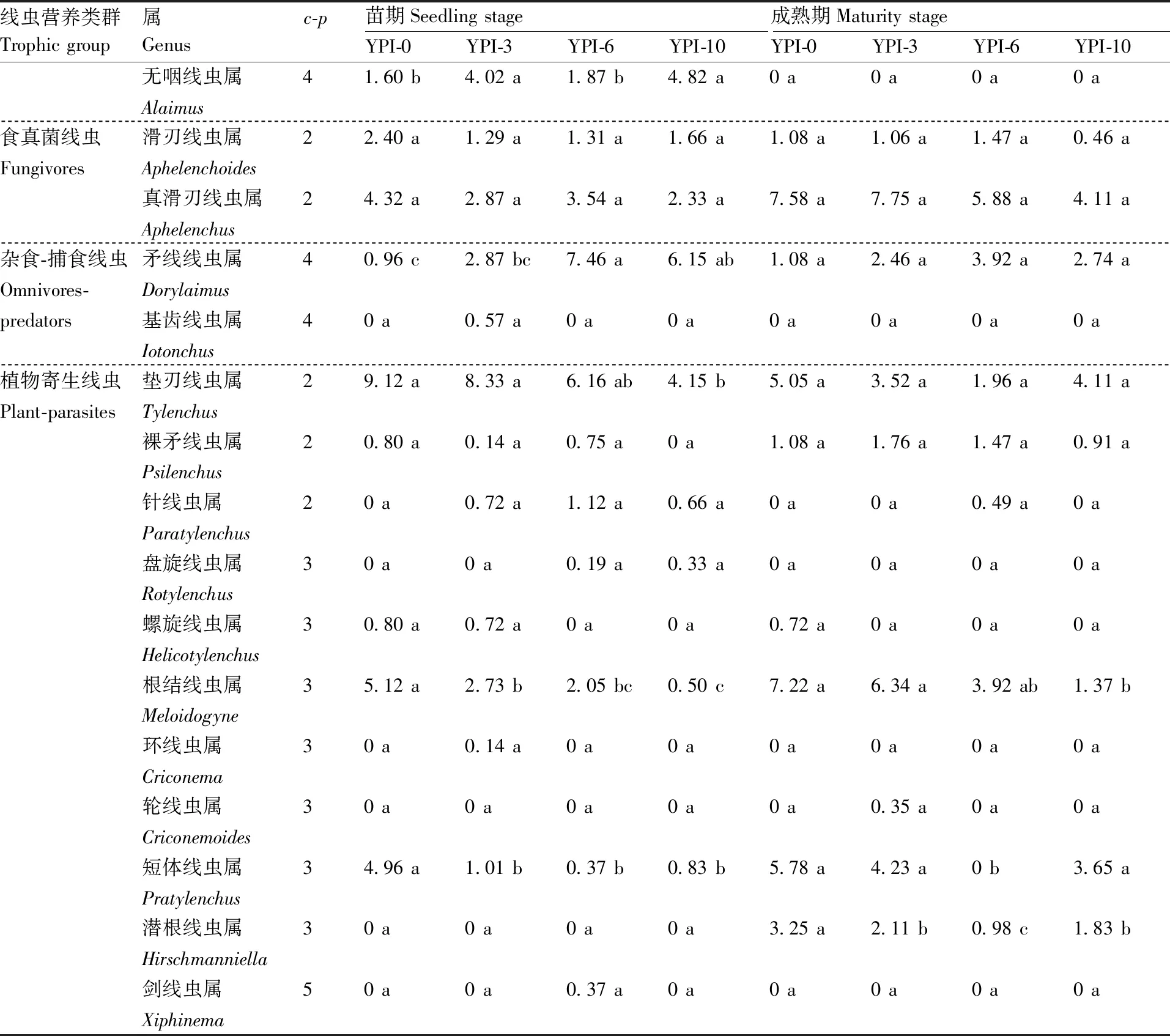
续表2 Continued Table 2
在苗期和成熟期,不同处理土壤中的食细菌线虫均以小杆线虫属(Rhabditis)、真头叶线虫属(Eucephalobus)和拟丽突线虫属(Acrobeloides)为极优势属(相对丰度≥10%[21]),同一时期,不同处理这些属的相对丰度间均无显著差异。成熟期时,小杆线虫属的相对丰度较苗期大幅增加,增幅达50%~100%,而拟丽突线虫属的相对丰度大幅降低,降幅在50%~75%。
食真菌线虫中,真滑刃线虫属(Aphelenchus)的相对丰度最高,且苗期和成熟期不同处理间的相对丰度均无显著差异。苗期不同处理下,依照土壤中真滑刃线虫属的相对丰度,均判定其为次优势属(相对丰度在2%~<5%),至成熟期时,YPI-0、YPI-3和YPI-6处理的真滑刃线虫属相对丰度大幅增加,均达到优势属的水平(相对丰度在5%~<10%),但YP-10处理下仍为次优势属。
杂食/捕食线虫中,YPI-3、YPI-6和YP-10处理下矛线线虫属(Dorylaimus)在苗期和成熟期土壤中的相对丰度达到次优势属或优势属水平,而在YPI-0处理下则为稀有属(相对丰度<1%)或常见属(相对丰度在1%~<2%)水平。
植物寄生线虫中,主要包括垫刃线虫属(Tylenchus)、根结线虫属(Meloidogyne)和短体线虫属(Pratylenchus)等,且其在苗期和成熟期不同处理间的相对丰度随两茬间隔年限的增加总体呈降低趋势。其中,YPI-10处理土壤中垫刃线虫属的相对丰度在苗期和成熟期均为次优势属的水平,且其在苗期时的相对丰度较YPI-0处理显著(P<0.05)降低54.50%。YPI-3、YPI-6和YPI-10处理下苗期时根结线虫属和短体线虫属的相对丰度均较YPI-0处理显著(P<0.05)降低,降幅分别达46.68%、59.96%和90.23%;至成熟期后,YPI-10处理的根结线虫属和YPI-6处理的短体线虫属相对丰度分别较YPI-0处理显著(P<0.05)降低81.02%和100%。由此可见,不同种植模式不同时期的土壤线虫群落种类具有较大差异,两茬间撂荒一定年限可以降低土壤中山药病原线虫的种群数量。
2.3 不同种植模式对土壤线虫营养类群的影响
如图1所示,不同种植模式对山药苗期和成熟期土壤中线虫营养类群的影响不同。在苗期和成熟期,YPI-10处理的土壤食细菌线虫的相对丰度均显著(P<0.05)高于YPI-0处理,而YPI-3和YPI-6处理均与YPI-0处理无显著差异。苗期时,不同处理土壤的食真菌线虫相对丰度无显著差异,但成熟期时YPI-10处理的食真菌线虫相对丰度显著(P<0.05)低于其他处理。苗期时,YPI-6和YPI-10处理的土壤杂食/捕食类线虫的相对丰度显著(P<0.05)高于YPI-0处理,但至成熟期后各处理间并无显著差异。苗期时,不同处理土壤的植物寄生线虫的相对丰度表现为YPI-0>YPI-3>YPI-6>YPI-10,且各处理间差异显著(P<0.05);至成熟期时,YPI-0和YPI-3处理的植物寄生线虫的相对丰度仍显著(P<0.05)高于YPI-6和YPI-10处理。以上结果说明,山药两茬间隔6 a或10 a可改善土壤线虫营养类群,不仅可提高食细菌线虫的相对丰度,还可有效控制植物寄生线虫的种群数量,尤以间隔10 a的作用效果最为明显。
2.4 不同种植模式对土壤线虫生态指数的影响
土壤线虫的多样性指数和生态功能指数分别指示土壤食物网的循环途径和稳定性。由表4可知,不同处理对山药苗期和成熟期土壤线虫的Shannon多样性指数(H’)和优势度指数(λ)均无显著影响。苗期不同处理的土壤线虫通路比值(NCR)无显著差异,但成熟期时YPI-10处理的却显著(P<0.05)高于其他处理。苗期和成熟期不同种植模式下的NCR值均大于0.5,表明不同种植模式对土壤有机质分解途径的影响不大,均以细菌降解为主。苗期不同处理的土壤线虫瓦斯乐斯卡指数(WI)表现为YPI-10>YPI-6>YPI-3>YPI-0,且各处理间差异显著(P<0.05);至成熟期时,YPI-6和YPI-10处理的WI仍显著(P<0.05)高于YPI-0和YPI-3处理。苗期,YPI-0处理的土壤植物寄生线虫成熟度指数(PPI)显著(P<0.05)高于YPI-10处理;成熟期,不同处理间无显著差异。自由生活线虫成熟度指数(MI)在苗期表现为YPI-3、YPI-6和YPI-10处理均显著(P<0.05)高于YPI-0处理,至成熟期YPI-6和YPI-10处理的MI显著(P<0.05)高于YPI-0和YPI-3处理。苗期,YPI-3、YPI-6和YPI-10处理的PPI/MI均显著(P<0.05)低于YPI-0处理;至成熟期,YPI-6和YPI-10处理的PPI/MI仍显著(P<0.05)低于YPI-0和YPI-3处理。由此可知,山药两茬间隔6 a或10 a可在一定程度上影响土壤生态系统的成熟度和稳定性。
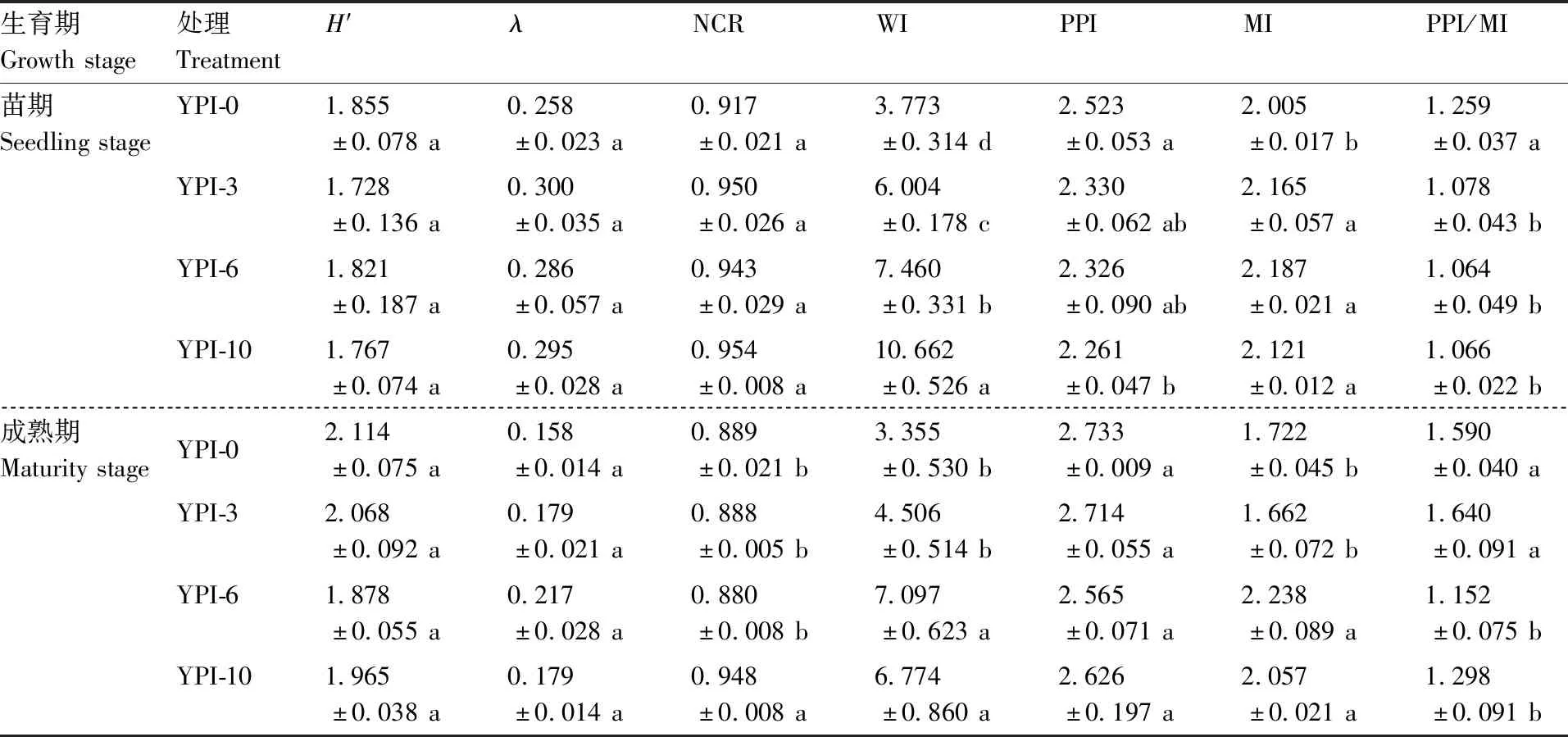
表4 不同种植模式对土壤线虫生态指数的影响Table 4 Effect of different planting patterns on ecological indices of soil nematodes
2.5 土壤线虫群落与土壤理化性质的相关性
经分析,苗期不同种植模式土壤线虫属与环境因子间无显著差异,因此仅通过RDA建立了山药成熟期不同种植模式土壤线虫属组成与土壤环境因子的排序图(图2),轴1和轴2共解释了土壤线虫属与土壤养分因子总体变异的77.7%。从RDA排序图可以看出,不同处理的土壤线虫群落组成和土壤环境因子显著(P<0.05)相关,土壤有机质、速效磷和速效钾对土壤线虫群落种属组成影响较大,而土壤pH值和铵态氮的影响较小。在YPI-0和YPI-3处理土样中,土壤速效磷和速效钾含量与小杆线虫属、根结线虫属、短体线虫属、垫刃线虫属、真滑刃线虫属等数量呈较强正相关,与拟丽突线虫属、棱咽线虫属(Prismatolainus)等少部分线虫属的数量呈较强负相关。YPI-6和YPI-10处理的土壤pH值、铵态氮、有机质与其所对应的大部分线虫属的数量均呈较强的负相关。在YPI-6处理土样中,土壤pH值、铵态氮、有机质与真头叶线虫属、单宫线虫属(Monhystera)、棱咽线虫属等的数量呈较强正相关;在YPI-10处理土样中,土壤pH值和铵态氮与棱咽线虫属、拟丽突线虫属等的数量呈较强正相关。
3 讨论
3.1 山药不同种植模式对土壤理化性质的影响
土壤理化性质是衡量土壤质量的重要指标,对山药的生长发育具有重要作用。本研究结果显示,从山药苗期至成熟期,不同种植模式下土壤速效钾含量大幅增长,而有机质、铵态氮和速效磷含量降低。这表明瑞昌山药在生长过程中对有机质、氮肥和磷肥有较大的需求,与范琳娟等[22]的研究结果基本一致。马祥华等[23]研究表明,土壤速效磷含量会随撂荒年限的增加而降低,具有明显的植被土壤效应;杜峰等[13]研究也表明,随着撂荒年限的增加,土壤速效磷含量会逐渐降低。这些研究结果均与本研究一致。推测本研究中土壤速效钾含量的变化可能与瑞昌山药的生长特性或田间管理习惯(膨大期时追施了大量钾肥)有关。
3.2 山药不同种植模式对土壤线虫群落结构的影响
不同种植模式对土壤理化性质的改变可通过直接或间接影响作物生长或土壤微生物活动而影响线虫的营养类群、群落结构和丰度[24]。Vink等[25]研究表明,休耕会对土壤线虫群落产生影响。李琪等[26]研究发现,撂荒可大幅降低土壤中短体线虫的相对丰度,但根结线虫在土壤中无寄主存在的情况下仍可存活约3 a。本研究结果也显示,随着两茬间撂荒年限的增加,不同种植模式土壤中植物寄生线虫丰度较高的垫刃线虫属、根结线虫属和短体线虫属的相对丰度基本均呈降低的变化趋势,而食细菌线虫,如小杆线虫属等的相对丰度大幅提高。小杆线虫属依据c-p值属于1类群,是典型的机会主义者,生命周期短,但繁殖能力强,耐受外界干扰的能力较强[27]。这与Walker[28]提出的“冗余种”假说基本一致,即为了维持相对稳定,某些线虫种群会大幅增加来替代不易存活的线虫。刘贝贝等[29]、焦加国等[30]研究表明,土壤线虫数量、群落组成与土壤养分、含水量,以及气候均有密切的关系。2019年7月中旬后江西省降雨一直极少,由此推测,成熟期山药土壤线虫数量大幅降低可能与土壤含水量太低有关。食细菌线虫对土壤食物网和营养循环具有重要作用,可作为指示土壤肥力的一个潜在指标[31]。高密度的植物寄生线虫种群数量不仅会对植物造成机械损伤,还有可能使植物感染微生物,对植物造成更严重的危害[32]。综合本研究结果,山药两茬间隔撂荒一定年限可在一定程度上影响土壤线虫数量、群落组成和各营养类群的相对丰度,改善土壤环境,减轻线虫病的发生程度。
本研究表明,山药不同种植模式对Shannon多样性指数和优势度指数无显著影响,这表明其对山药土壤线虫群落中的稀有种多样性和属内线虫的数量无显著影响。这与陈璐等[33]研究的草原植被人工恢复4 a和8 a对土壤线虫群落多样性的影响基本一致。WI值可反映土壤的健康状况,当WI值>1时,值越大,说明土壤健康状况越好[19]。MI值越高,生态系统成熟度越高,抵抗外界干扰的能力越强;PPI值则与扰动的频率呈正相关,PPI/MI值在受扰动的情况下会升高[20]。本研究表明,两茬间隔6 a或10 a的处理下,在山药苗期和成熟期,其WI值显著高于间隔0 a的处理,而PPI/MI值显著低于间隔0 a的处理。这表明撂荒一定年限有利于提高土壤的健康状况,提高土壤生态系统的抗干扰能力。
在农田生态系统中,影响土壤线虫群落多样性的因素有很多,主要包括耕作方式、养分管理、土壤含水量和食物的可利用性等[34-35]。本研究的RDA排序图同样表明,山药成熟期时土壤养分因子与线虫群落具有一定的相关性,土壤中丰度较高的植物寄生线虫,如垫刃线虫属、根结线虫属、短体线虫属等数量与土壤速效磷、速效钾含量呈正相关,与土壤pH值,及有机质和铵态氮含量呈负相关,而大部分食微线虫与土壤pH值,及有机质和铵态氮含量呈正相关。这与苏兰茜等[36]的研究结果基本一致。7月上旬至10月上旬是瑞昌山药茎叶和块茎生长最旺盛的时期,这一时期山药对水肥的要求比较高。不同种植模式下,土壤养分状况有很大的变化,而土壤生物与土壤环境具有密切的关系,据此推测,可能是土壤养分的改变通过影响地上植被生长状况和微生物活动,进而改变了土壤线虫的群落结构[33]。苗期时土壤养分因子与线虫群落无显著相关性,可能是由于瑞昌山药在苗期生长较为缓慢,不同种植模式下山药的生长对土壤环境的影响较小,因此其相关性未达显著水平。
本研究表明,山药两茬间撂荒6 a或10 a,一方面可使土壤速效磷含量显著降低,影响山药对土壤中磷元素的吸收;另一方面可显著降低土壤中植物寄生线虫的相对丰度,增加土壤食细菌线虫相对丰度,提高土壤生态系统的健康状况和抗干扰能力,且以两茬间撂荒10 a的效果更佳。这说明改变种植模式可以在一定程度上控制山药病原线虫的种群数量,是替代化学防治行之有效的办法,但基于对现有耕地的有效利用,建议山药与其他非线虫寄主植物进行轮作或间作。
猜你喜欢
今日农业(2022年14期)2022-09-15
土壤学报(2022年2期)2022-06-09
中国生物防治学报(2022年1期)2022-04-22
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
农药科学与管理(2021年11期)2022-01-12
房地产导刊(2020年9期)2020-10-28
载人航天(2020年3期)2020-07-02
中国计算机报(2017年47期)2018-01-12
电影文学(2016年23期)2017-02-13
江苏农业科学(2016年10期)2017-02-05
