
芽胞杆菌FJAT-2349 脂肽对枇杷采后炭疽病菌抑制作用
2021-03-04 01:11:06陈梅春邓英杰陈燕萍肖荣凤郑雪芳朱育菁王阶平
福建农业学报 2021年12期
陈梅春,邓英杰,陈燕萍,肖荣凤,郑雪芳,朱育菁*,王阶平*,刘 波
(1. 福建省农业科学院农业生物资源研究所,福建 福州 350003;2. 华中农业大学 生命科学技术学院,湖北 武汉 430000)
0 引 言
【研究意义】炭疽病是枇杷生产过程中最严重的病害之一,主要危害枇杷幼苗、叶片和果实,该病会引起苗木死亡和果实腐烂,造成巨大的经济损失[1]。其中,由炭疽病菌(Colletotrichum acutatum)引起的炭疽病是枇杷果实采后贮存、运输中发生的主要病害,该病严重时损失达50%以上[2]。开展枇杷果实采后炭疽病病害防控研究,对于延长枇杷货架期、降低枇杷采后损失具有重要意义。【前人研究进展】枇杷采后炭疽病是一类侵染性病害,病原菌炭疽病菌始于果园,于枇杷果实成熟后侵入果实扩展致其发病,常使得贮藏期果实大量腐烂。枇杷炭疽病发病时,果实表面先产生淡褐色水渍状圆形病斑,而后变得干缩凹陷,且表面密生出许多小黑点,潮湿时表面溢出粉红色黏物,病斑继续发展,致使全果褐变腐烂或干缩呈僵果[3]。目前常用的枇杷果实采后病害的防治手段为低温冷藏和化学杀菌剂[2,4],但因方法自身的特点使得它们在枇杷采后病害防治领域的应用受到了限定,如冷藏温度过低容易使枇杷果实发生冷害,品质下降,失去商品性;长期大量使用化学杀菌剂会使病菌产生抗药性、污染环境及危害人类健康,因此研究安全有效的防治措施势在必行,其中生物防治是最具应用前景措施之一[5]。芽胞杆菌脂肽具有广谱抗菌活性、稳定性好、不易产生耐药性,对人畜毒性低且对环境友好等优点,能够作为生防试剂用于水果采后病原菌的防治,如桃子上施用枯草芽胞杆菌fmbJ 脂肽后,防腐率可达76.5%[6];葡萄、草莓和西红柿上施用贝莱斯芽胞杆菌XT1 脂肽后,疾病的发生率下降了12%~100%[7];Calvo 等[8]将解淀粉芽胞杆菌BUZ-14 脂肽应用于水果保鲜,发现其能够完全抑制桃子褐腐病和苹果蓝腐病的发生;Jin 等[9]研究发现贝莱斯芽胞杆菌HN-2 产生的脂肽bacillomycin D 具有强拮抗芒果炭疽病菌作用,其半抑制浓度仅有阳性杀菌剂的1/3~1/6;Lin 等[10]将Bacillomycin D 和chitosan 混合处理樱桃番茄,降低了果实软腐病和灰霉病的发病率,使樱桃番茄货架期延长15 d。【本研究切入点】关于脂肽在枇杷采后病害防治方面研究还有待深入探讨。【拟解决的关键问题】以枇杷采后炭疽病原菌为靶标,筛选出产脂肽芽胞杆菌生防菌株,探究芽胞杆菌脂肽对枇杷采后炭疽病的抑制作用,为枇杷采后炭疽病生物防治提供新资源,同时,为果蔬采后病害生防机制的深入研究奠定基础。
1 材料与方法
1.1 供试材料
供试菌株 解淀粉芽胞杆菌(Bacillus amyloliquefaciens)FJAT-2349;枇杷采后炭疽病菌(Colletotrichum acutatum)FJAT-30256;香蕉尖孢镰刀菌(Fusarium oxysporum)FJAT-370;甜瓜尖孢镰刀菌(F. oxysporum)FJAT-9230;龙眼采后腐烂菌小新壳梭孢菌(Neofusicoccum parvum)FJAT-3531 由本实验室分离、保存。
供试枇杷品种:解放钟,产地:莆田。
1.2 菌株培养
解淀粉芽胞杆菌FJAT-2349 培养:菌株FJAT-2349 接种至50 mL 马铃薯葡萄糖肉汤液体培养基中,25 ℃、170 r·min−1振荡培养48 h。
真菌培养:供试真菌于马铃薯葡萄糖肉汤固体培养基上培养5 d 后,将菌株FJAT-370、FJAT-9230和FJAT-3531 接种至50 mL 的马铃薯葡萄糖肉汤液体培养基中,于30 ℃、170 r·min−1下培养3 d 后,调整含量为107CFU·mL−1,备用;用无菌水将固体平板上的枇杷炭疽病菌FJAT-30256 孢子洗脱下来,制备孢子悬浮液,含量调整为107CFU·mL−1。
1.3 解淀粉芽胞杆菌菌株对枇杷炭疽病菌的抑菌检测
采用平板对峙法探究解淀粉芽胞杆菌对枇杷炭疽病菌的抑菌作用。用1 mL 无菌枪头于新鲜长满枇杷炭疽病菌的平板边缘打孔钻取菌饼,将长菌一面朝下置于PDA 平板中央作为指示菌,从活化好的待筛选芽胞杆菌平板上挑取单菌落,在距离菌饼3 cm 处划线。以只接种枇杷炭疽病菌菌饼作为空白对照。随后置于25 ℃培养箱,培养5−7 d,计算菌株对枇杷炭疽病菌的抑菌率。抑菌率/%=[(A−B)−(C−B)]/(A−B)×100。式中A 为对照菌落直径(mm);B 为菌饼直径(mm),C 为处理组菌落直径(mm)。
1.4 解淀粉芽胞杆菌脂肽的制备
解淀粉芽胞杆菌FJAT-2349 于PDA 液体培养基中培养48 h 后,将发酵液离心弃去菌体得上清液。于上清液中加入2 mol·L−1盐酸至pH<2,将其置于4 ℃下静置24 h 后,离心获得沉淀。上述沉淀经低温冷冻干燥后即为粗脂肽粉末。
1.5 解淀粉芽胞杆菌脂肽对真菌抑制活性检测
吸取枇杷采后炭疽病菌孢子悬浮液0.5 mL,加入到50 ℃ 80 mL 的PDA 半固体培养基(0.9%琼脂)中,混匀后倾覆在预先已凝固的PDA 下层固体培养基上(1.8%琼脂)。待上层培养基凝固冷却后,在平板中打孔(直径7 mm),分别注入不同含量脂肽的溶液80 μL。以无菌甲醇为空白对照, 28 ℃下培养3~5 d 后测抑菌圈直径。每个处理3 次重复。
1.6 解淀粉芽胞杆菌脂肽对枇杷采后炭疽病菌活体抑制试验
挑选果形端正、无机械伤、无裂果,无病虫斑的枇杷作为试材。枇杷经酒精表面消毒后,用无菌水洗涤,晾干待用。试验设置4 个处理组,分别为治愈性对照组、治愈性处理组、预防性对照组和预防性处理组。
治愈性对照组:每个枇杷果实腰部用消毒的枪头刺伤一个点,深度为1.5 mm。每个刺伤点的枇杷炭疽病菌孢子悬浮液用量5 μL,孢子含量为1.0×104CFU·mL−1。24 h 后,刺伤口加入5 μL 的清水。
治愈性处理组:每个枇杷果实腰部用消毒的枪头刺伤一个点,深度为1.5 mm。每个刺伤点的枇杷炭疽病菌孢子悬浮液用量5 μL,孢子含量为1.0×104CFU·mL−1。24 h 后,刺伤口加入50 mg·mL−1的脂肽溶液5 μL。
预防性对照组:每个枇杷果实腰部用消毒的枪头刺伤一个点,深度为1.5 mm。每个刺伤点加入清水5 μL,24 h 后加入枇杷炭疽病菌孢子悬浮液5 μL,孢子含量为1.0×104CFU·mL−1。
预防性处理组:每个枇杷果实腰部用消毒的枪头刺伤一个点,深度为1.5 mm。每个刺伤点加入50 mg·mL−1的脂肽溶液5 μL,24 h 后加入枇杷炭疽病菌孢子悬浮液5 μL,孢子含量为1.0×104CFU·mL−1。
每个试验组枇杷个数为24 个,为了加快发病速度,枇杷储存温度为28~30 ℃,湿度为75%~80%;每天观察发病情况,统计发病率和病斑直径。
1.7 扫描电镜和透射电镜
样品处理:从枇杷炭疽病菌与解淀粉芽胞杆菌FJAT-2349 脂肽抑菌圈边缘靠近病原菌一侧切取培养基块,每个处理切取—块。
扫描电镜:样品采用2.5%戊二醛进行样品固定,经PBS 缓冲液、乙醇、叔丁醇处理风干后喷金,利用扫描电镜观察,以未处理正常生长的枇杷炭疽病菌菌块为对照。
透射电镜:样品采用戊二醛、锇酸双重固定法固定,在透射电镜下观察,以未处理正常生长的枇杷炭疽病菌菌块为对照。
1.8 数据处理
采用DPS 9.5 对数据进行显著性检验。
2 结果与分析
2.1 解淀粉芽胞杆菌FJAT-2349 对枇杷炭疽病菌的抑制作用
通过平板对峙法筛选到1 株对枇杷炭疽病菌FJAT-30265 具有强抑制作用的菌株FJAT-2349(图1),该菌株鉴定为解淀粉芽胞杆菌[11],其对枇杷炭疽病菌的抑菌带直径为(17.64±0.25) mm,抑菌率可达87.8%。解淀粉芽胞杆菌FJAT-2349 能够产生脂肽,该脂肽由iturin、 fengycin 和surfactin 三大家族组成。菌株发酵上清液中的脂肽总含量为(207.21±15.46) mg·L−1,其中fengycin 占脂肽总量的87.8%[11]。
2.2 解淀粉芽胞杆菌脂肽对枇杷炭疽病菌离体抑制作用
采用抑菌圈法测定解淀粉芽胞杆菌FJAT-2349 脂肽对枇杷采后炭疽病菌的抑制活性,结果如表1所示。研究发现该菌产生的脂肽对枇杷炭疽病菌具有很好的抑制作用,脂肽含量为30 mg·mL−1时,其抑菌圈直径可达(22.75±0.31) mm,且抑制效果呈剂量效应,浓度越高,抑菌效果越好。同时,本研究还探究了FJAT-2349 脂肽的抑菌谱,发现该菌产生的脂肽对香蕉和甜瓜尖孢镰刀菌及龙眼致腐菌小新壳梭孢菌均具有强抑制作用,当脂肽含量为10~30 mg·mL−1时,抑菌圈直径为17.43~25.46 mm。
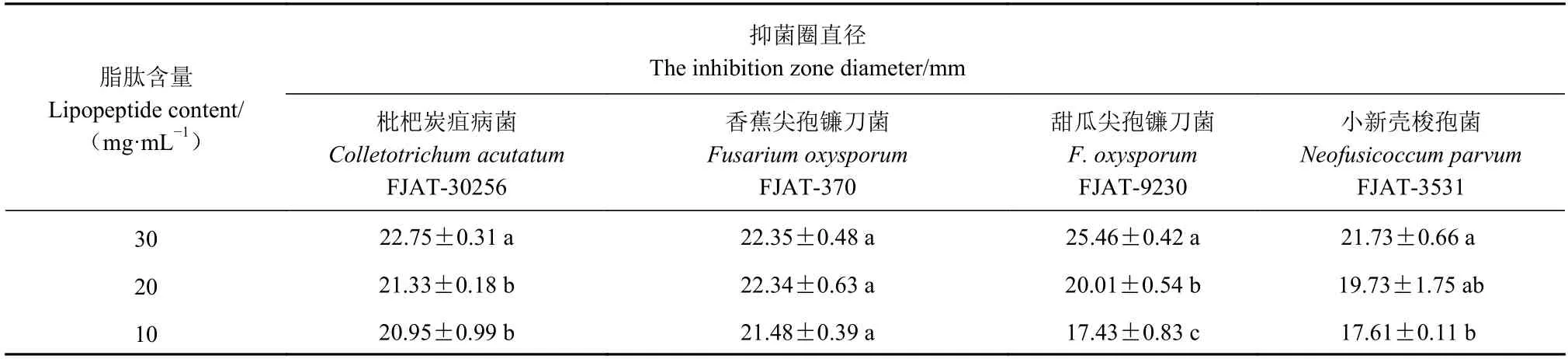
表1 不同含量脂肽对真菌的抑制作用Table 1 Antifungal effect of lipopeptides
2.3 解淀粉芽胞杆菌脂肽对枇杷炭疽病菌活体抑制作用
通过人工刺伤接种的方法探究脂肽对枇杷采后炭疽病菌活体抑制作用,先接种病原菌后接种清水/脂肽的治愈性组和先接种清水/脂肽后接种病原菌的预防性组的发病率和病斑直径如表2、3 所示。
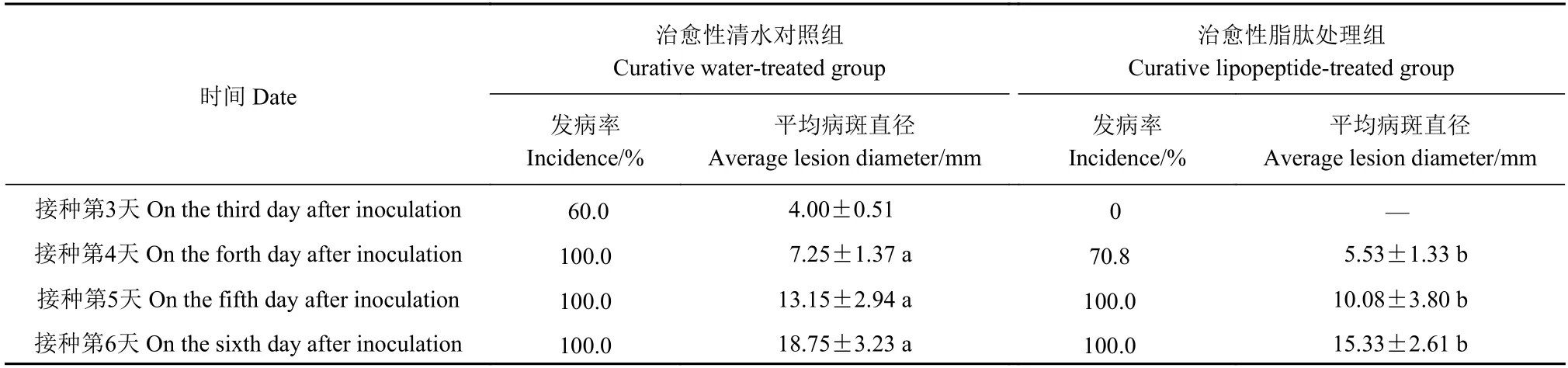
表2 脂肽对枇杷炭疽病害治愈性试验结果Table 2 Effect of lipopeptides in curing anthracnose on postharvest loquats
从表2 中可以看出,治愈性清水对照组于接种第3 天开始发病,发病率达60.0%,平均病斑直径为4.00 mm,而治愈性脂肽处理组还未发病。当治愈性清水对照组发病率达100.0%时,平均病斑直径7.25 mm;治愈性脂肽处理组发病率为70.8%,平均病斑直径为5.00 mm,其发病率和平均病斑直径均低于对照组,计算相对防效为29.2%;随着存储时间的延长,治愈性脂肽处理组平均病斑直径仍低于对照组。上述结果说明脂肽能够推迟枇杷炭疽病的发病时间,并在一定程度上减轻枇杷炭疽病的发病程度。
从表3 中可以看出,预防性清水对照组于接种第3 天开始发病,发病率达27.8%,平均病斑直径为5.00 mm,而预防性脂肽处理组还未发病。当预防性清水对照组发病率达100.0%时,平均病斑直径6.89 mm;预防性脂肽处理组发病率仅为27.3%,平均病斑直径为4.67 mm,其发病率和平均病斑直径均低于对照组,相对防效为72.7%;随着存储时间的延长,预防性脂肽处理组发病率缓慢上升,其平均病斑直径小于对照组。上述结果说明脂肽能够推迟枇杷炭疽病的发病时间,降低枇杷炭疽病的发病率,并在一定程度减轻枇杷炭疽病的发病程度。与治愈性脂肽处理组相比,预防性脂肽处理组发病速度慢,发病率低,这说明先用脂肽处理枇杷,能更有效地降低枇杷炭疽病的发生率,减轻炭疽病的发病程度。
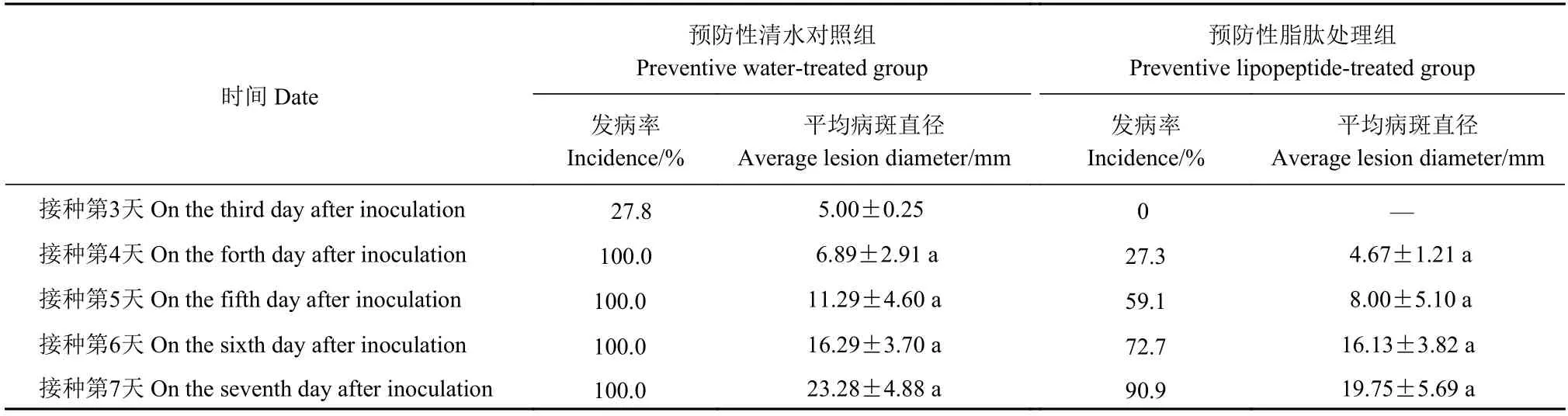
表3 脂肽对枇杷炭疽病害预防性试验结果Table 3 Effect of lipopeptides in preventing anthracnose on postharvest loquats
2.4 解淀粉芽胞杆菌脂肽对枇杷炭疽病菌形态结构的影响
利用电镜观察正常枇杷采后炭疽病菌菌丝(对照组)和抑菌圈边缘的枇杷炭疽菌丝形态(处理组)。扫描电镜结果如图2 所示,结果显示正常的枇杷采后炭疽病菌菌丝表面光滑、结构完整饱满、粗细均匀;抑菌圈边缘的枇杷炭疽病菌菌丝发生异常,呈不规则的肿大和皱缩,明显畸形,这表明抑菌脂肽会致使菌丝体发育畸形,影响炭疽病菌的正常生长。
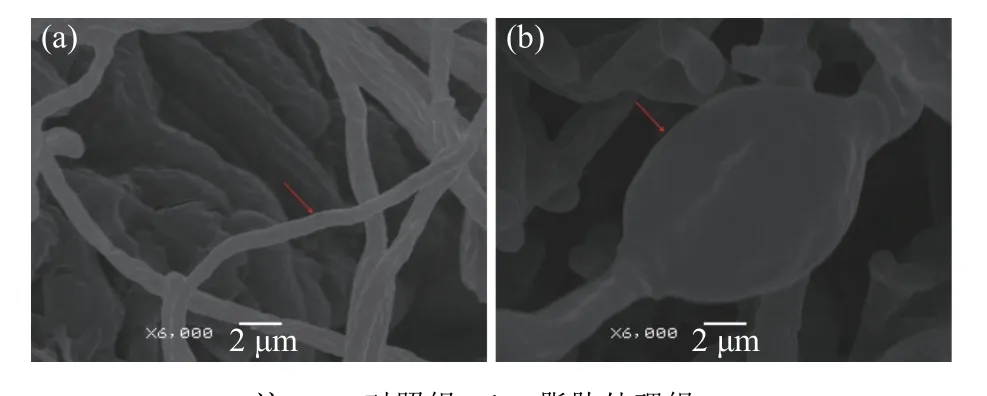
图2 枇杷炭疽病菌菌丝形态扫描电镜观察Fig. 2 Scanning electron micrograph of C. acutatum hyphae
透射电镜结果如图3 所示,结果显示正常的枇杷炭疽病菌菌丝细胞壁厚度均一,细胞内原生质均匀,细胞结构完好无损;脂肽处理后,菌丝细胞壁变薄或消失,细胞膜被破坏,细胞内容物减少,内部组成紊乱,从而导致导致菌丝细胞解体死亡。这说明脂肽通过破坏枇杷炭疽病菌丝体细胞的完整性,从而导致细胞死亡。
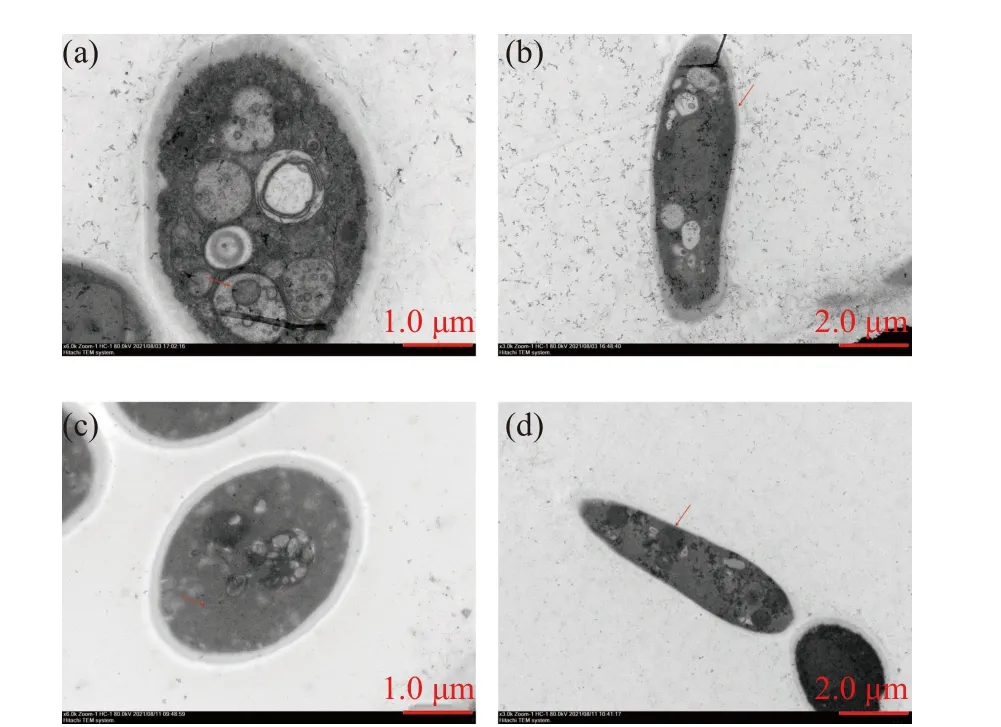
图3 枇杷炭疽病菌菌丝形态透射电镜观察Fig. 3 Transmission electron micrograph of C. acutatum hyphae
3 讨论与结论
芽胞杆菌产生的脂肽包括iturin、fengycin 和surfactin 等,脂肽的环肽是由羟基/β-氨基脂肪酸与7/10 个氨基酸以酞氨键连接形成的[12−13]。不同菌株产生的脂肽组成不同,这与其合成调控基因相关。例如,解淀粉芽胞杆菌S76-3 产生的脂肽由C14−C15iturinA、C13−C16surfactin 和 C14−C17PlipastatinA、 C16−C17plipastatin B 组成[14];解淀粉芽胞杆菌 SYBC H47 产生的脂肽由C13−C15Bacillomycin L、C16−C17fengycin A/B 和C13−C15surfactin 组成[15]。本研究中解淀粉芽胞杆菌FJAT-2349 产生的脂肽由iturin(C14–C17iturin A)、fengycin(C14−C15fengycin A、C14fengycin B2、C16fengycin A2/B2、C16−C17fengycin B、C14fengycin A 衍生物和C15fengycin B 衍生物)和surfactin(C12−C16surfactin A)组成[11]。脂肽组成不同,其抑菌能力不同,如iturin 能够强烈抑制酵母菌和真菌的生长,但对细菌抑菌作用较弱;fengycin不仅能抑制丝状真菌,对细菌也有较好的拮抗作用;surfactin 主要作用于细菌和有脂膜包被的病毒,其对真菌的抗性不佳[16]。研究报道,芽胞杆菌iturin和fengycin 家族脂肽具有较强拮抗炭疽病菌的作用,如枯草芽胞杆菌KS03 脂肽iturins[17]、解淀粉芽孢杆菌RC-2 脂肽iturinA2[18]、贝莱斯芽胞杆菌HN-2 脂肽bacillomycin D[9]、苏云金芽胞杆菌CMB26 类fengycin脂肽[19]等。本研究中的解淀粉芽胞杆菌FJAT-2349脂肽能抑制枇杷采后炭疽病菌的生长,该脂肽处理枇杷果实后,推迟了枇杷采后炭疽病的发病时间,降低了枇杷采后炭疽病的发生率,有效地减轻枇杷采后炭疽病的发病程度。菌株FJAT-2349 产生脂肽中fengycin 占总量的87.8%,推测其为抑制枇杷采后炭疽病菌的主要拮抗物质。
芽胞杆菌脂肽主要通过破坏病菌细胞膜来发挥其抗真菌作用,如苏云金芽胞杆菌CMB26 产生的类fengycin 脂肽作用于炭疽病菌表面后,使其菌丝发育畸形,从而影响菌丝正常生长[19];贝莱斯芽胞杆菌HN-2 脂肽bacillomycin D 会破坏芒果炭疽病菌细胞壁和细胞膜,使细胞内细胞质和细胞器渗出后形成空腔,进而导致病菌死亡[9]。本研究中通过扫描电镜和透射电镜观察发现,被解淀粉芽胞杆菌FJAT-2349 脂肽抑制的枇杷炭疽病菌丝肿胀变形,细胞壁变薄或消失,细胞内容物减少,内部组成紊乱,这表明菌株FJAT-2349 产生的抑菌脂肽会使枇杷炭疽病菌丝体发育畸形,影响枇杷炭疽病菌的正常生长。本研究明确了解淀粉芽胞杆菌发酵生产的脂肽对枇杷炭疽病菌的抑制作用,这为脂肽的后续研究及在水果采后病害防治中的推广应用奠定了基础。
猜你喜欢
中国生物防治学报(2022年3期)2022-07-09 10:00:22
微生物学杂志(2021年2期)2021-07-01 11:01:06
小学生必读(中年级版)(2021年12期)2021-03-03 07:43:44
小学生必读(中年级版)(2021年11期)2021-02-22 05:19:06
宁夏医学杂志(2020年4期)2021-01-21 08:25:16
微生物学杂志(2020年2期)2020-12-31 07:17:13
山东化工(2020年4期)2020-03-30 09:18:18
分析科学学报(2019年2期)2019-04-25 03:36:56
衡阳师范学院学报(2016年3期)2016-07-10 07:16:27
物理化学学报(2015年7期)2015-12-30 12:13:08
