
Therapeutic potential of trehalose in neurodegenerative diseases: the knowns and unknowns
2021-03-03 15:04MasoomehKhalifehGeorgeBarretoAmirhosseinSahebkar
中国神经再生研究(英文版) 2021年10期
Masoomeh Khalifeh, George E. Barreto, Amirhossein Sahebkar
Neurodegenerative diseases (NDs) are a growing health problem associated with a high burden due to destructive and longterm clinical effects. Cellular aggregations of misfolded proteins are the most common pathological hallmark of many late-onset NDs called proteinopathies including Parkinson’s disease (PD), Alzheimer’s disease (AD),tauopathies, amyotrophic lateral sclerosis(ALS), and polyglutamine (polyQ) expansion diseases such as Huntington’s disease (HD)and various spinocerebellar ataxias (SCA)such as SCA3 (Renna et al., 2010). Misfolded proteins can be generated by posttranslational conjugation (e.g., hyperphosphorylated tau in AD), or endoproteolytic cleavage (e.g., amyloid β peptides) or genetic mutations in specific proteins (such as HTT in HD, α-synuclein in PD,PrPC in prion disease and SOD1 and TDP-43 in ALS) leading to the formation of oligomers.
Generally, misfolded proteins do not accumulate in the cells and are cleared through cellular quality control machinery. However,oligomers are partially resistant to all known proteolytic pathways and tend to grow into fibrillar inclusion bodies (Ciechanover and Kwon, 2015). Moreover, β-sheet-enriched aggregates can interact with various cellular molecules and impair the ubiquitin-proteasome system as well as chaperone-mediated autophagy either directly or indirectly.Emerging evidence suggests that autophagy is closely linked to NDs (Ciechanover and Kwon,2015). Biochemical analyses have shown that autophagic vacuoles are abundant in the postmortem brain samples of many NDs, and are co-localized with pathological proteins(Tan et al., 2014). Therefore, the notion that stimulation of autophagy ameliorates neuropathology and neurodegeneration has garnered considerable attention and various agents targeting autophagy pathway have been investigated as potential therapies in experimental studies.
Trehalose is one such candidate, which was reported to have neuroprotective effects in animal models of various NDs, such as PD, AD,and HD. Trehalose, also known as mycose, is a nonreducing disaccharide that is made of two glucose molecules (linked in a α, α-1, 1 configuration) and is abundantly present in microorganisms, plants, and invertebrates.Physical and chemical properties of trehalose arise from its nonreducing property, which leads to chemical stability, high hydrophilicity,and strong resistance to both acid hydrolysis and cleavage by glucosidases (Lee et al., 2018).Trehalose ingestion is generally safe as part of the diet and does not evoke a rapid increase in blood glucose levels. At doses up to 50 g,trehalose is safe for human, and no symptom has been reported in the majority of subjects except trehalose-deficient individuals in whom;gastrointestinal side effects may occur. In animal studies, no deaths were observed following intravenous administration, though diarrhea sporadically occurred. Trehalose is absorbed similarly to that of glucose by either the oral or intravenous administration. However, it is partially hydrolyzed to glucose by the enzyme trehalase present in the intestinal brush border and the proximal tubules of the kidneys, liver,and blood (Emanuele, 2014).
Trehalose can protect biomacromolecules and biological systems using a variety of different mechanisms (Figure 1). It was initially known as a chemical chaperone that has a stabilizing activity on protein structures and prevents protein aggregation. Trehalose was shown to stabilize the partially unfolded Huntingtin protein and inhibit β amyloid (Aβ)and scrapie prion protein (PrPSc) aggregation in cellular models of HD, AD, and prion diseases, respectively (Lee et al., 2018).Moreover, trehalose could exert significant antioxidant and anti-inflammatory effects both in vivo and in vitro by scavenging oxygen free radicals and preventing inducible nitric oxide synthase expression or nitric oxide formation,respectively (Emanuele, 2014).
Recently, autophagy induction has been proposed as the principal theory to explain the mechanism of neuroprotection by trehalose in proteinopathy models (both in vitro and in vivo) (Hosseinpour-Moghaddam et al., 2018). In cell culture experiments, trehalose treatment enhanced the clearance of mutant Huntingtin,α-synuclein, and tau proteins by increasing autophagic flux in a variety of mammalian cells.It was proposed that these protective effects were mediated by intracellular trehalose.Similar observations were made in animal studies in which oral and intraperitoneal administration of trehalose induced autophagy and reduced protein aggregation. Moreover,inflammation and gliosis were decreased,and behavioral and neurobiological defects were partially recovered in response to trehalose (Lee et al., 2018). For example,oral administration of trehalose significantly reduced insoluble SOD1 and ubiquitinated proteins in the spinal cord tissues of ALS mice.In the HD transgenic mouse model, trehalose reduced polyglutamine aggregates in the brain, improved motor dysfunction, reduced brain atrophy, and extended the lifespan compared with glucose-fed control mice,suggesting that the neuroprotective effects were not associated with increased glucose levels following trehalose hydrolysis (Yoon et al., 2017). Trehalose improved cognition and diminished the abnormal Aβ deposition in the hippocampus of AD transgenic mouse model. Additionally, trehalose promoted synaptic activity via upregulation of growth factors and neurogenesis markers in the mouse hippocampus that might be correlated with cognitive improvement. Trehalose treatment also decelerated pathological features in the mouse model of PD (Khalifeh et al., 2019,2020). An oral solution of trehalose decreased the abnormal aggregation of PABPN1 in muscle cells and improved muscle weakness in transgenic model of oculopharyngeal muscular dystrophy (Emanuele, 2014).
Despite the positive direction toward autophagy activation in several models of ND, there is no precise delineation of the mechanism by which trehalose drives autophagy. It was shown that trehalose, as an mTOR-independent autophagy activator, stimulates de novo expression of autophagic proteins including SQSTM1/p62 and MAP1LC3B, and promotes LC3-I to LC3II conversion. It also acts through modulating different molecular chaperones like HSP90 and SigmaR1. It has been reported that trehalose induces autophagy in hepatocytes in an AMPKdependent manner via inhibition of the glucose transporter SLC2A8/GLUT8, which produces glucose starvation and activates autophagy.Nevertheless, to trigger autophagy, trehalose needs to be imported into the cells, and deletion of SLC2A8, which also acts as an importer of trehalose, prevents its ability to stimulate autophagy (Lee et al., 2018; Khalifeh et al., 2020).
A recent study has indicated that trehalose exerts its metabolic effects through changes in lysosomes, causing their rapid enlargement and transient permeabilization. Such lysosomal modifications might be triggered by lysosomal osmotic stress, which leads to lysosomal Ca2+release and PPP3/calcineurin activation. PPP3/calcineurin as a phosphatase specifically targets TFEB and induces its translocation to the nucleus. TFEB-induced autophagic flux stimulates the expression of the genes required for autophagy activation,including ATG10, ATG12, BECN1, SQSTM1/p62 and, MAP1LC3B. Moreover, trehalaseresistant analogues of trehalose, melibiose and lactulose, showed similar effects (Rusmini et al., 2019). Lysosomal breakdown and accumulation of autophagosomes have also been described in PD brains, and TFEB colocalization with α-synuclein was detected in Lewy bodies (Khalifeh et al., 2019). It is noteworthy that autophagosome impairment and defective autophagosome-lysosome fusion have also been found in other NDs. Some studies suggested that the neuroprotective mechanism of trehalose might be indirect in animals. When orally administrated, trehalose might be hydrolyzed in the gut, raising the possibility that the neuroprotective effects of trehalose are through microbiota-gut-brain signaling (Lee et al., 2018). An in vitro study indicated that trehalose blocked the autophagic flux that would be inconsistent with the notion that it is an autophagy inducer. The authors reported that trehalose impaired lysosomal membrane integrity and led to the prevention of autophagosome-lysosome fusion and accumulation of α-synuclein in neuronal cells;however, cell viability did not change (Khalifeh et al., 2019). Remarkably, Martano et al. (2017)showed that trehalose is present in the rodent hippocampus as an endogenous metabolite.They found that trehalose is biosynthesized in primary astrocytes as an endogenous School of Pharmacy Mashhad University of Medical Sciences, Mashhad, Iran (Sahebkar A)
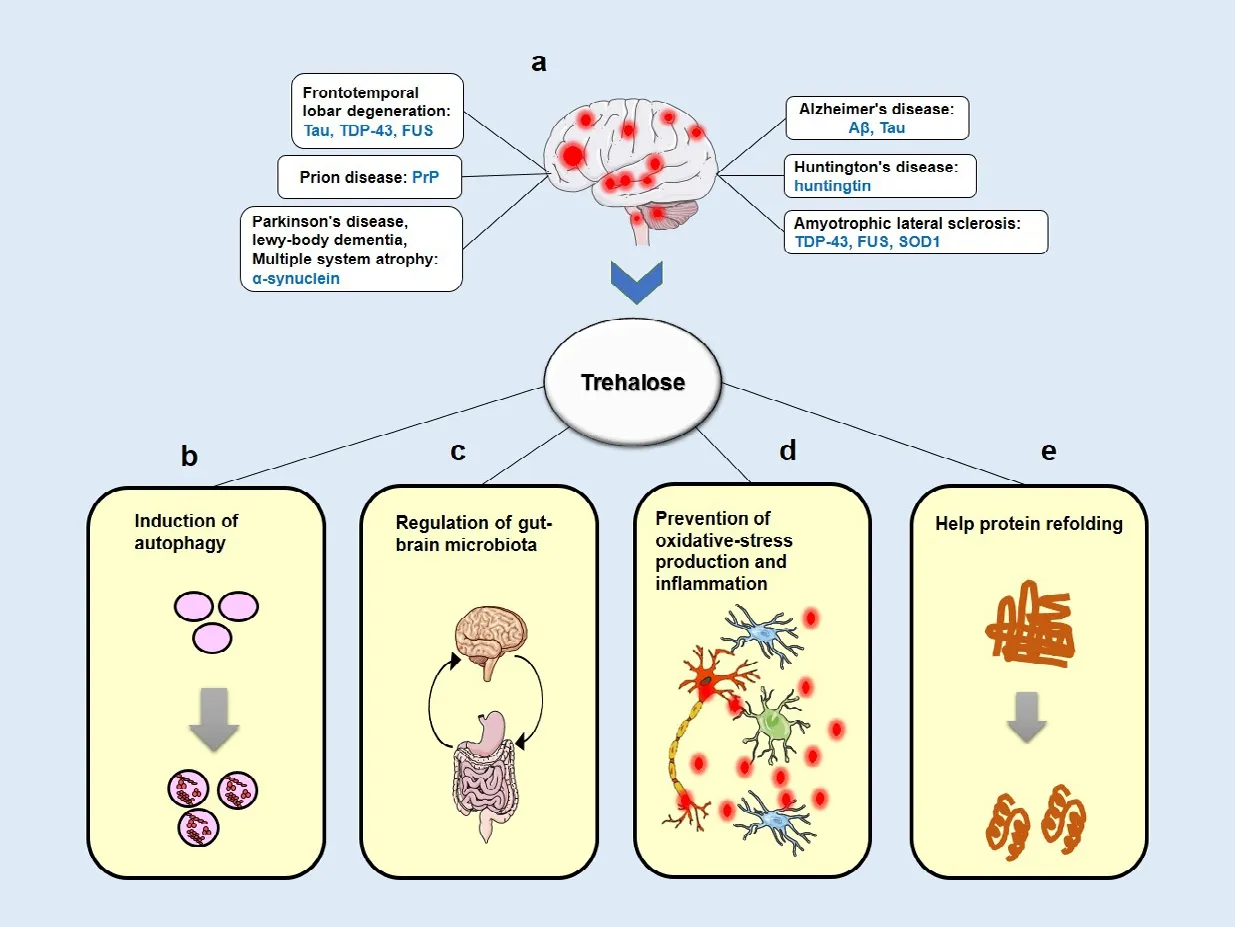
Figure 1|A schematic view of the mainly hypothesized neuroprotective mechanisms by trehalose.
*Correspondence to:Amirhossein Sahebkar,PharmD, PhD, sahebkara@mums.ac.ir or amir_saheb2000@yahoo.com.https://orcid.org/0000-0002-8656-1444(Amirhossein Sahebkar)
Date of submission:September 30, 2020
Date of decision:November 19, 2020
Date of acceptance:January 13, 2021
Date of web publication:February 19, 2021
https://doi.org/10.4103/1673-5374.308085
How to cite this article:Khalifeh M, Barreto GE,Sahebkar A (2021) Therapeutic potential of trehalose in neurodegenerative diseases: the knowns and unknowns. Neural Regen Res 16(10):2026-2027.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.metabolite that is used by neurons and implicated in neuronal maturation by increasing dendritic arborization. Besides, trehalase enzyme in the mouse brain is mainly detected in neurons but not in astrocytes (Martano et al.,2017; Halbe and Rami, 2019). Investigating the interaction between trehalose and trehalase and the mechanism that are responsible for neuronal arborization would be interesting.
The therapeutic potential of trehalose is appealing due to its beneficial effects in different models of neurodegeneration that strongly support its applications in the clinical setting. Till now, several clinical trials have been performed with trehalose, commonly in diseases such as macular edema, depressive disorder, and muscular dystrophies. In a recent open-label study without placebo arm in SCA3 patients, daily administration of 100 g trehalose via oral route exhibited a mild improvement in the severity of the disease.Gastrointestinal side effects were reported in seven patients, and two withdrew due to severe diarrhea. In humans, around 99.5% of oral trehalose is enzymatically metabolized in the small intestinal epithelium and is not readily absorbed as an intact molecule, hence larger doses of trehalose is required to achieve adequate blood concentrations and this might lead to gastrointestinal symptoms such as diarrhea and flatulence. Therefore, intravenous administration has been suggested as an alternative administration approach, which could provide higher tissue concentrations of trehalose. The safety, tolerability and efficacy of intravenous trehalose (13.5 g or 27 g per week) have been recently tested in a phase II trial in patients with SCA3, and no serious adverse event was identified in for up to 1 year(Zaltzman et al., 2020). Moreover, intravenous trehalose solution (90 mg/mL) was observed to be safe and well-tolerated in patients with oculopharyngeal muscular dystrophy, and multiple recorded endpoints improved upon trehalose administration.
In summary, there is mounting evidence on the therapeutic potential of trehalose in NDs.Trehalose is generally recognized as a safe substance and a widely used FDA-approved ingredient in pharmaceutical preparations,including parenteral products. Its safety can facilitate further clinical testing of this promising agent in clinical studies. It appears that trehalose may elicit more significant effects when administered at the presymptomatic stage of NDs before the appearance of widespread neurodegeneration. Besides the timing of treatment, attaining information on the proper dosing and administration route of trehalose is also necessary. It is still unknown if the neuroprotective effects of trehalose follow a dose-dependent pattern. Furthermore, while the intravenous route provides a significantly higher bioavailability of trehalose compared to the oral route, it remains elusive as to whether the gut microbiota mediates a main role in the neuroprotective effects of trehalose. Further studies are also needed to explore whether alterations in brain function occur in the direct or indirect pathway of trehalose.
Masoomeh Khalifeh, George E. Barreto,Amirhossein Sahebkar*
Department of Medical Biotechnology and Nanotechnology, Faculty of Medicine, Mashhad University of Medical Sciences, Mashhad, Iran(Khalifeh M)
Department of Biological Sciences; Health Research Institute, University of Limerick, Limerick,Ireland (Barreto GE)
Applied Biomedical Research Center;
Biotechnology Research Center, Pharmaceutical Technology Institute, Mashhad University of Medical Sciences, Mashhad, Iran (Sahebkar A)Polish Mother’s Memorial Hospital Research Institute (PMMHRI), Lodz, Poland (Sahebkar A)

- 中国神经再生研究(英文版)的其它文章
- Axonal regeneration and sprouting as a potential therapeutic target for nervous system disorders
- The role of gap junctions in cell death and neuromodulation in the retina
- Don’t know what you got till it’s gone: microglial depletion and neurodegeneration
- Low-dose lipopolysaccharide as an immune regulator for homeostasis maintenance in the central nervous system through transformation to neuroprotective microglia
- Protein post-translational modifications after spinal cord injury
- Axonal mRNA localization and local translation in neurodegenerative diseases