
双氢青蒿素对人宫颈癌细胞Hela生物学特性的影响
2021-03-02 00:29张新影高翠红孙翀
广东医学 2021年2期
张新影,高翠红,孙翀
1天津市津南医院妇产科(天津 300350);2石家庄市妇产医院妇产科(河北石家庄 050000);3天津市中心妇产科医院妇产科(天津 300100)
宫颈癌是女性的头号杀手及妇科常见的高危恶性肿瘤疾病,也是全球范围内第三大最常见的癌症[1]。宫颈癌的有效局部控制主要采用放射治疗,放疗敏感性降低导致的放射治疗的临床失败越来越受到人们的关注,尽管如此,肿瘤放射治疗敏感性降低的分子机制尚不完全清楚。临床上药物治疗的一些不良反应和不良预后导致了紧急情况下需要使用新的化学治疗剂。而最近的研究表明[2],抗疟药青蒿素及其衍生物可显著抑制宫颈癌细胞的增殖,已使用多年成功治疗人类疟疾和病毒。双氢青蒿素(DHA)是青蒿素(ART)的活性代谢产物,可对多种人类癌症包括子宫颈癌、乳腺癌、卵巢癌和结直肠癌细胞等有细胞杀伤作用毒性发挥作用。最近的研究表明,其抗癌活性包括DHA在内的ART衍生物主要是通过铁介导的过氧化物桥的裂解来实现[3]。癌细胞膜上转铁蛋白受体数量和胞内Fe2+浓度远高于正常细胞,DHA对癌细胞具有选择性毒性。此外,临床试验已经表明,人体对ART衍生物微量药物具有很好的耐受性,在疟疾治疗中没有发现明显的不良反应,可见DHA在肿瘤学上具有重要的潜力[4-5]。但是,它的作用机制还没有阐明。因此,2018年10月至2019年8月我们开展本实验主要探索DHA对人宫颈癌Hela细胞的影响及相关机制。
1 材料与方法
1.1 材料 Hela细胞株(中国科学院,北京),Trizol试剂盒、胎牛血清、RPMI 1640培养基(北京沃比森科技有限公司)双氢青蒿素、Bax、caspase-3、GAPDH单克隆抗体(sigma公司,美国),Matrigel基质胶(BD生物技术公司,美国),Western blot试剂盒、IP细胞裂解液(碧云天生物技术研究所);PVDF(美国Millipore公司);相关引物(由上海生工生物工程有限公司合成);普通光学倒置显微镜(Nikon)、流式凋亡及细胞周期试剂盒、Annexin V-FITC/PI试剂盒(上海吉凯公司)CO2培养箱(Thermo)。
1.2 细胞培养和分组 人宫颈癌细胞株Hela细胞培养于含10% 小牛血清及100 U/mL左氧氟沙星的RPMI 1640培养基中,于5%CO2及37℃条件下密闭式孵箱内培养,每1~2 d行细胞传代,后续实验均取对数生长期的细胞。按密度为每毫升 2×105个的宫颈癌Hela细胞接种于 6 孔板,每孔 3 mL,于5%CO2及37℃条件下密闭式孵箱内培养 24 h。空白对照组(加入等体积生理盐水)、低浓度DHA组 (50 μmol/L)及高浓度DHA 组(400 μmol/L)。48 h后进行后继检测
1.3 RT-PCR检测 分组及加药情况同1.2,培养48 h后收集三组Hela细胞,加入TRIzol试剂裂解细胞抽提总RNA。根据逆转录说明书合成相应的cDNA并进行扩增。条件如下95℃行预变性60 s,95℃下变性30 s,59℃退火30 s,71℃延伸30 s,共40个循环。Bax上游引物:5′-ACACCTGAGCTGACCTTGGA-3′,下游引物:5′-CCGTGTCCACGTCAGCAATC-3′,引物长度为133 bp;Caspase-3上游引物:5′-AGAGCTGGACTGCGGTATTGAG-3′,下游引物:5′-GAACCATGACCCGTCCCTTG-3′,引物长度为148 bp;内参GAPDH上游引物:5′-CGTGGAAGGACTCATGACCA-3′,下游引物:5′TCCAGGGGTCTTACTCCTTG-3′,引物长度为509 bp。以GAPDH为内参,测定Caspase-3、Bax mRNA相对表达水平。
1.4 Western blot检测 分组及加药情况同1.2,培养48 h后收集三组Hela细胞,加入RIPA蛋白裂解液中,提取总蛋白,通过BCA法测定各组蛋白含量,10%聚丙烯酰氨凝胶(SDS-PAGE)电泳,15 mA电泳3 h转膜至聚偏氟乙稀(PVDF)膜,然后置于4℃条件下进行封闭2 h,Bax、caspase-3及GAPDH一抗(1∶1 000)孵育过夜,二抗(1∶1 500)室 温孵育1 h,暗室加入ECL发光液后曝光后显影,X线胶片照相,行灰度值分析。
1.5 流式细胞仪检测细胞凋亡 分组及加药情况同1.2,培养48 h后收集三组Hela细胞,将细胞制成单细胞悬液,1 000 r/min离心5 min沉淀细胞孵育缓冲液洗1次,用100 μL的Annexin V-PI标记溶液重悬细胞,室温下避光孵育15 min,加入荧光(SA-FLOUS)溶液,避光4℃下振动孵育20 min,上流式细胞仪检测。实验重复至少3次。
1.6 流式细胞仪检测细胞周期 分组及加药情况同1.2,培养48 h后收集三组Hela细胞,于37℃下,胰蛋白酶(浓度为0.25%)消化后,1 500 r/min,离心10 min,离心半径15 cm,除去细胞固定液,通过预冷的70%乙醇固定细胞过夜,PBS洗1次后加入PI的染色液(含RNase A) 0.5 mL,避光于37℃下温浴30 min,冰浴放置保存,流式细胞仪行细胞周期检测,实验重复3次。
1.7 克隆形成实验 分组及加药情况同1.2,培养48 h后收集三组Hela细胞,于 37℃、5%CO,环境下连续培养14 d。通过甲醇固定 30 min后,经过0.5%结晶紫染色30 min后,于光镜下对含 50个细胞以上的克隆数计数。
1.8 Transwell侵袭实验 分组及加药情况同1.2,培养48 h后收集三组Hela细胞,按50 μg/孔Matrigel 包被Transwell小室底部膜,下室内为含10% FBS的RPMI 1640培养基500 μL/孔,向上室加入每组200 μL(3×105/mL)细胞悬液,于孵育箱内常规培养24 h后取出,拭去上室未穿膜细胞,4%多聚甲醛固定后,结晶紫染色5 min,于显微镜下计数穿膜细胞。

2 结果
2.1 Caspase-3、Bax mRNA表达水平 RT-PCR检测显示Caspase-3、Bax mRNA表达空白对照组、低浓度DHA组分别为1.01±0.09、0.98±0.06,1.46±0.12、1.42±0.14,高浓度DHA 组为2.45±0.27、2.37±0.21,明显高于上述两组(P<0.05),因此,DHA干预调高Caspase-3、Bax mRNA在宫颈癌Hela细胞的表达,可能通过该途径调控诱导细胞凋亡,见图1。
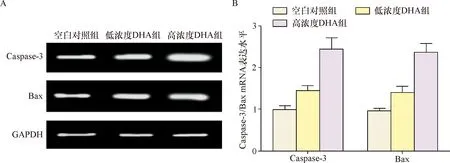
注:A:Caspase-3、Bax mRNA电泳图;B:Caspase-3、Bax mRNA的表达
2.2 Caspase-3、Bax蛋白表达水平 Western blot检测显示Caspase-3、Bax蛋白表达空白对照组、低浓度DHA组分别为0.48±0.07、0.51±0.08,0.95±0.11、1.03±0.12,高浓度DHA 组为2.28±0.21、2.35±0.23,明显高于上述两组(P<0.05),因此,DHA干预调高Caspase-3、Bax蛋白在宫颈癌Hela细胞的表达,可能通过该途径调控诱导细胞凋亡,见图2。
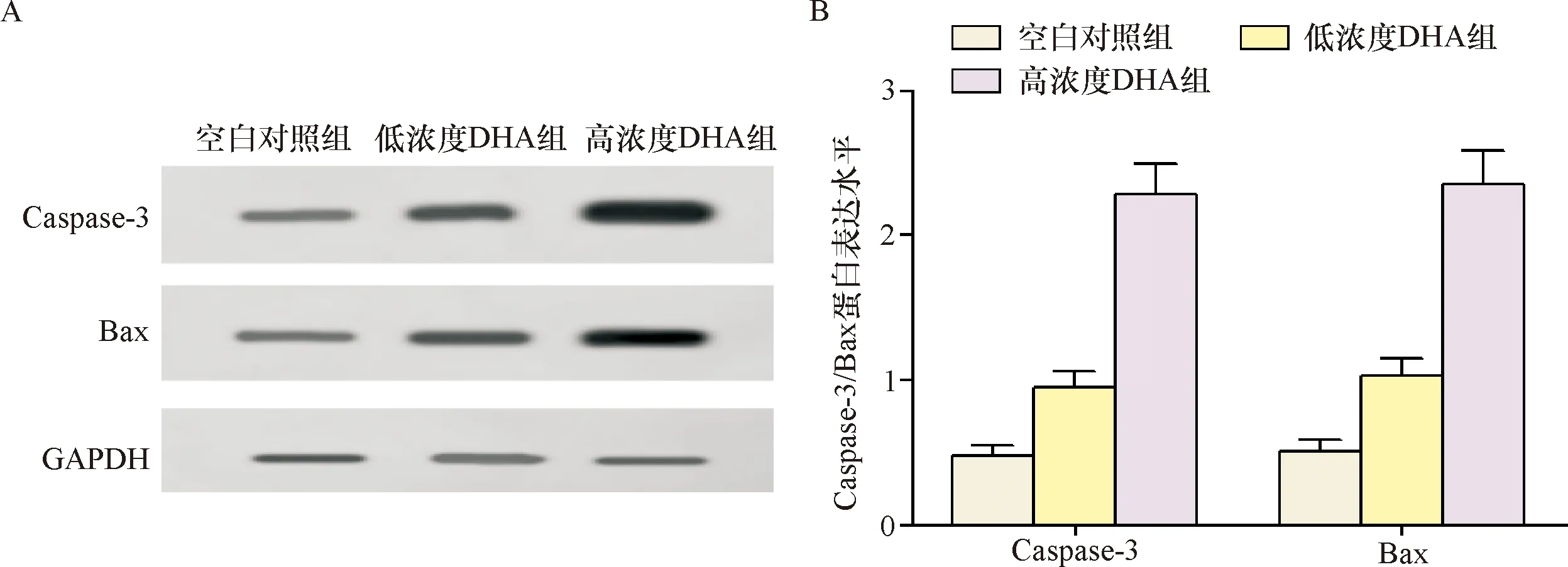
注:A:Caspase-3、Bax蛋白的电泳图;B:Caspase-3、Bax 蛋白的表达
2.3 细胞周期及凋亡率变化 细胞周期结果显示G0/G1期与S 期细胞空白对照组、低浓度DHA组分别为53.52±3.98、32.84±3.18,61.64±3.92、22.47±2.31,高浓度DHA 组为76.48±5.26、13.25±1.22,细胞周期阻止于G0/G1期(P<0.05),S期细胞数减少(P<0.05);凋亡率结果显示高浓度DHA 组凋亡率为(20.41±0.92)%,明显高于空白对照组和低浓度DHA组[(2.84± 0.61)%、(10.36±0.63)%](P<0.05)。可见DHA干预后Hela细胞凋亡率升高,高浓度DHA较低浓度效果明显(P<0.05),见表1,图3。

注:A:空白对照组;B:低浓度DHA组;C:高浓度DHA 组
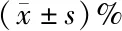
表1 各组细胞周期的分布以及凋亡率
2.4 Hela细胞增殖活力 平板克隆实验显示高浓度DHA 组细胞克隆形成数为(62.54±12.46)个,明显低于空白对照组和低浓度DHA组[(188.56±14.84)个,(119.56±14.62)个](P<0.05),可见DHA干预后Hela细胞增殖活力降低,高浓度DHA较低浓度效果明显(P<0.05),见图4,表2。

注:A:空白对照组;B:低浓度DHA组;C:高浓度DHA 组
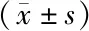
表2 每组细胞克隆形成数和穿膜细胞数 个
2.5 Transwell侵袭实验 Transwell侵袭实验穿膜细胞数高浓度DHA 组为(92.75±13.54)个,明显低于空白对照组和低浓度DHA组[(232.58±26.42)个,(128.75±22.16)个](P<0.05)。可见DHA干预抑制了 PATU8988细胞的侵袭能力,高浓度DHA较低浓度效果明显(P<0.05),见图5,表2。
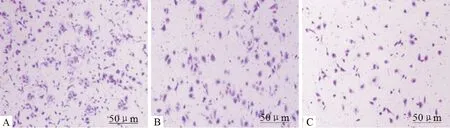
注:A:空白对照组;B:低浓度DHA组;C:高浓度DHA 组
3 讨论
DHA被世界卫生组织推荐为有效的抗疟复方中药。最近研究表明DHA可对多种癌细胞的生存能力发挥作用[6]。DHA是青蒿素的活性代谢物,据报道它抑制了一些癌细胞体外培养及抑制病毒诱导的小鼠肿瘤的形成[7]。最近的一些研究表明DHA在不同细胞系中的作用与DHA介导的细胞凋亡和细胞周期调控有关。有研究表明DHA对癌细胞的抑制作用是基于p53和线粒体的激活细胞凋亡途径,DHA可通过p53非依赖性Caspase-9介导的途径诱导细胞凋亡,抑制宫颈癌细胞的增殖[8]。DHA在多种癌细胞中的细胞毒性已经被许多研究证明[9-11]。然而,DHA的抗癌作用机制还没有充分的证据。
据报道,DHA可通过诱导对癌细胞产生抑制、凋亡作用。DHA治疗已被证明会导致线粒体膜去极化、细胞色素c释放和Caspase活化[12]。已有研究表明,DHA可诱导Bax蛋白表达上调,促进癌细胞凋亡[13]。而且,一些临床试验报告显示,DHA可通过抑制细胞增殖,促进细胞凋亡改善临床晚期子宫颈癌患者的症状[14-16]。在本研究中我们使用不同浓度的DHA处理后,Caspase-3、Bax蛋白得到不同程度的上调。我们发现,高浓度DHA 组中Caspase-3、Bax蛋白表达明显高于空白对照组及低浓度DHA组,表明DHA通过上调Caspase-3和Bax抑制宫颈癌的生长。
此外,通过流式细胞仪检测细胞周期及凋亡变化我们发现,DNA损伤诱导G1期阻滞,DNA损伤诱导最显著的阻滞发生在G0/G1期,S期延迟进展。因此我们推测DHA能抑制Hela细胞的细胞周期、促进细胞凋亡。我们采用平板克隆实验检测DHA作用 Hela细胞后的细胞增殖情况,结果表明DHA对Hela细胞有增殖抑制作用。实验结果显示高浓度DHA 组细胞克隆形成数明显低于空白对照组和低浓度DHA组,表明其伴有剂量依赖性。我们采用Transwell侵袭实验检测细胞侵袭力,结果显示高浓度DHA 组的穿膜细胞数明显低于空白对照组和低浓度DHA组,说明DHA对Hela细胞有降低细胞侵袭能力的作用,且其伴有剂量依赖性。
总而言之,我们对人宫颈癌Hela细胞进行DHA处理后,Caspase-3、Bax蛋白表达迅速上调。此外,我们证明了高浓度DHA 可以加速人宫颈癌Hela细胞的凋亡,降低细胞侵袭能力,DHA在Hela细胞中可表现出良好的抗肿瘤作用。上述这些结果将为DHA用作癌症治疗目标提供了证据支持,这种抗疟药的临床应用可以扩展以补充癌症治疗药物战略。尽管取得了这些进展,但是治疗后细胞死亡途径上游信号传导与下游激活的确切联系与DHA的关系尚不清楚,有待进一步研究阐明。
猜你喜欢
资源节约与环保(2022年8期)2022-09-20
有色金属(矿山部分)(2021年4期)2021-08-30
基层中医药(2020年8期)2020-11-16
农药科学与管理(2019年10期)2019-04-20
国际呼吸杂志(2019年4期)2019-03-12
浙江农业科学(2016年11期)2016-05-04
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
现代冶金(2016年6期)2016-02-28
医学研究杂志(2015年5期)2015-06-10
