
中国大菱鲆引进群体三代选育之后的收获体重遗传进展评估
2021-02-15 09:26:20胡玉龙王伟继
中国农学通报 2021年35期
孙 松,胡玉龙,吕 丁,王伟继
(1中国水产科学研究院黄海水产研究所,农业农村部海洋渔业可持续发展重点实验室,山东青岛 266071;2青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室,山东青岛 266071)
0 引言
大菱鲆(Scophthalmus maximus,Linnaeus,1758),广泛分布于欧洲大西洋沿岸,具有生长快、耐低温能力强、风味鲜美等优点,是世界上养殖最广泛的鲆鲽鱼类[1-2]。自1992年将大菱鲆引进到中国以来,大菱鲆产业在国内迅速发展。2010年以来,中国大菱鲆养殖产量一直维持在6万t左右,是世界上大菱鲆产量最高的国家。由于大菱鲆产业的巨大价值,开展遗传改良工作显得尤为重要,针对一系列重要经济性状的遗传改良是进一步促进大菱鲆产业发展的关键途径。在水产动物养殖生产活动中,生长性状是最具有商业价值的重要性状,生长速度越快,越能缩短饲养周期,从而降低成本。
选择育种是对物种进行遗传改良的基础方法,它可以随着核心育种群的世代的延续而持续获得遗传进展,而对核心选育群体的扩繁可以将使获得遗传进展在数以万计的后代得到表达[3-7]。选择育种已经应用到许多水产动物的遗传改良中,其作用得到广泛证实[8-9],例 如 罗非 鱼(Oreochromis niloticus)[10],凡纳 滨对 虾(Litopenaeus vannamei)[11-12],罗氏沼虾(Macrobrachium rosenbergii)[13-15]以 及 中 国 对 虾 (Fenneropenaeus chinensis)[16]等,涉及到的目标性状包括生长、出肉率、存活率和抗病力等。而在大菱鲆研究领域,关于选择育种效果评估的研究还鲜有报道。
2006年开始,黄海水产研究所启动了一项基于最佳线性无偏估计(Best linear unbiased prediction,BLUP)的大菱鲆选育项目[17]。截至2019年,已经完成590个家系的构建,并记录了3个选育世代(G1,G2和G3)的收获体重表型数据。本研究评估了不同选育世代中的收获体重遗传力,并以选择差异(Selection differential)和遗传进展(Genentic gain)为指标评估了多个世代选育之后得到的选育进展情况。本研究的结果将为评估现有大菱鲆遗传选育策略及下一步的大菱鲆选育工作提供参考。
1 材料与方法
1.1 实验材料
育种项目是在山东省海阳市黄海水产研究所鲆鲽鱼类遗传育种中心开展。大菱鲆奠基群体是在2003年和2004年引自5个国家(丹麦,法国,西班牙,英国和智利)的幼鱼。选择形态表现正常的个体作为候选亲本,注射无线射频标记(Radio Frequency Identification,RFID)标记后将其放养在5 m×5 m×0.6 m(长×宽×高)的养殖池中,密度为2~3 kg/m2。繁殖前3个月,通过控制养殖池的光照,水温和营养强化以刺激性腺发育。光照为200~600 lx,照射时间从8 h/天逐渐增加到10 h/天,水温从8℃升高到14℃,以鲜杂鱼代替商品料进行营养强化。基础群体G0于2006年和2007年通过不同来源群体间的双列杂交和人工授精建立。随机交配不同来源的雄性和雌性,通过不平衡的巢式交配设计构建全同胞家系和半同胞家系[6]。
受精卵在孵化72~120 h后,将所有家系上浮卵移入0.5 m3玻璃钢桶内,每个家系桶尽量保证培养过程一致。分别在孵化后第35天和第70天进行两次家系苗种数量标准化,即每个家系分别随机选取1000尾和400尾个体继续培养。孵化后100天,每个家系随机选取80尾大于平均体重的个体,称取平均体重,注射可视荧光标记(Visible Implant Fluorescent Elastomer,VIE)记录家系信息,建立系谱。荧光标记后将其随机分配到多个5 m×5 m×0.6 m性状测试池中。孵化后15个月,测量所有存活个体收获体重,进行BLUP遗传评估,综合家系和个体的育种值大小,选取育种值较大的60%的家系和个体作为亲鱼备用,注射RFID标记(子弹)以区分个体。G1,G2,G3的培育过程与G0一样,并根据系谱信息将子代近交系数控制在0.1以下[3,18]。
1.2 数据分析
本研究梳理了2006—2019年大菱鲆构建家系的系谱信息,利用整理后的系谱计算个体间的亲缘关系,构建分子亲缘关系矩阵(Numerator relationship matrix),单性状动物模型和限制性极大似然法(REML)方法(ASReml软件)[19]用于估计各方差组分和计算遗传参数,混合效应模型是经过条件AIC指数比较得到的最优模型[20],计算见公式(1)。
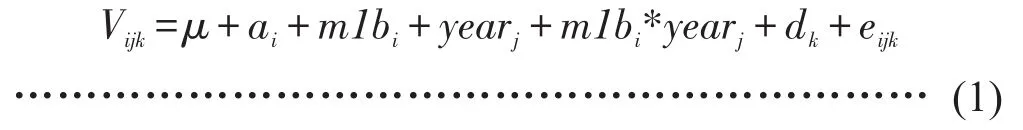
式中,Vijk为体重表型观察值;μ是总体均值;ai为第i个子代的随机加性遗传效应;m1bi是第i尾个体的初始体重;yearj是年份j的固定效应;m1bi*yearj为年份与初始体重的互作效应;dk是随机母本和共同环境效应;eijk是随机残差。
假设随机效应ai,dk和eijk的分布是正态的,其均值为0。方差-协方差矩阵表示见公式(2)。
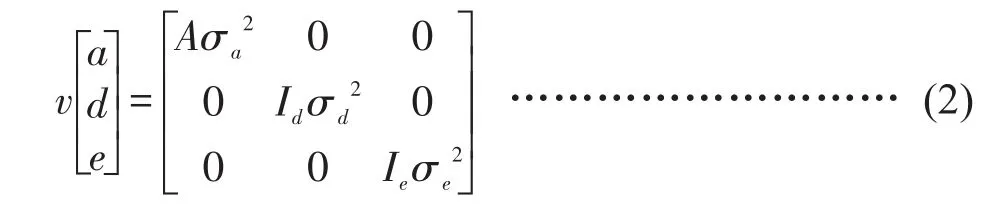
式中,ai,dk和eijk为随机效应a、d、e的方差;A是分子亲缘关系矩阵,Id、Id是单位矩阵。
选择差(Selection differential)为下一个世代个体的亲本的平均估计育种值与亲本所属世代平均估计育种值的差值。通过比较当前和上一代的平均估计育种值,还可以计算出每一代的遗传进展。累积遗传进展表示为百分比,见公式(3)。

式中,pc为累计遗传进展,pi为第i个世代的遗传进展,%,i的取值范围为1~3。
2 结果
2.1 描述性统计
主要统计性结果如表1所示。2006—2019年共建立了590个全同胞家系,收集了12980尾个体的收获体重。2008、2011和2018年由于亲鱼性腺发育的数量较少,所以这3年没有构建家系,而2010年的数据未记录谱系信息因而没有纳入数据统计中。将整理后的系谱作树状图(visPedigree package),4个世代包括引进群体如图1所示,由于亲本个体发育的速度并不一致,因此1个世代包含2~3年构建的家系。
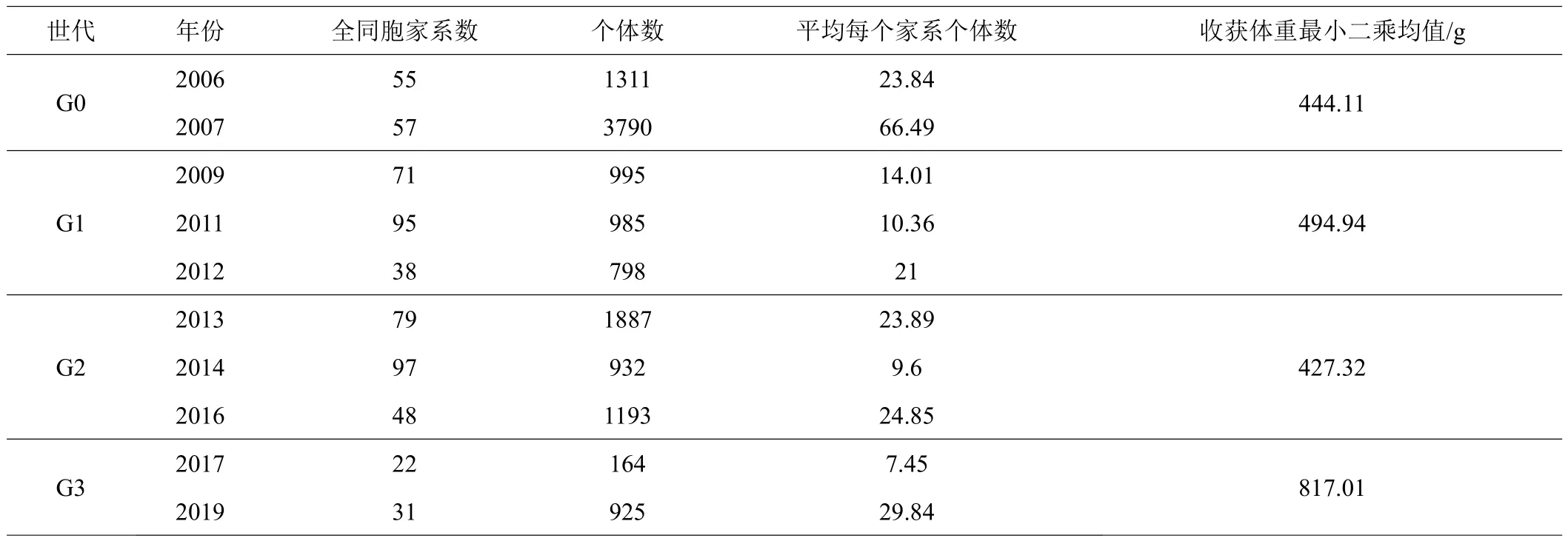
表1 每个世代的培育年份、家系数量、个体数量、收获体重最小二乘均值
2.2 遗传分析
不同世代方差组分,遗传力和共同环境效应等如表2所示。GO、G1的体重遗传力均低于G2和G3,其中G2世代遗传力最大为0.51±0.09,跨世代的遗传力最低,仅有0.16±0.03。共同环境效应G3世代最小为0.01±0.05,跨世代的共同环境效应最大,为0.28±0.02,且跨世代遗传力和共同环境效应的标准误均小于单一世代。
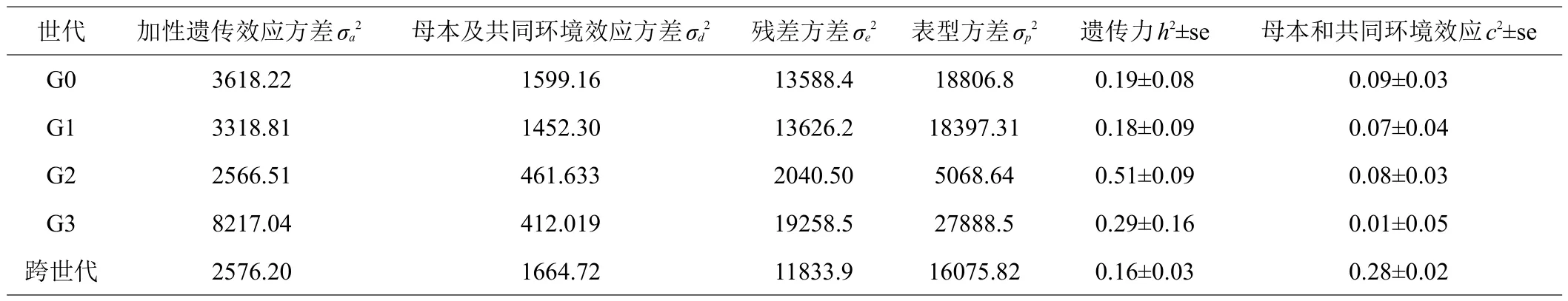
表2 各个世代及跨世代群体体重性状方差组分,遗传力,母本和共同环境效应
2.3 遗传进展
表3列出了每一代所有个体和亲鱼的平均估计育种值。生长选择差在G0中为45.6 g,在G1中为13.1 g,而在G2代是-27.9,相当于每代平均10.3 g。遗传进展在G1中为10.2 g,在G2中为37.4 g,G3为0.3 g,分别相当于2.06%、8.76%和0.04%。三代后的累积遗传进展为体重的10.86%,平均每代3.56%。

表3 每一代平均估计育种值,繁殖个体平均育种值,选择差异及遗传进展
3 讨论
3.1 遗传力
本研究中,除G2世代为高遗传力,其余估计到的体重遗传力均为中低等遗传力(基于以下的标准:低水平遗传力(0.05~0.15),中水平遗传力(0.20~0.40),高水平遗传力(0.45~0.60),及非常高(>0.65)[21]。遗传力估计值在不同世代之间变化很大(0.16~0.51),较高的遗传力与较低的共同环境效应有关,而较低的遗传力一般也是由较高的共同环境效应造成,这表明当前数据中加性遗传效应和共同环境效应之间存在一定程度的混淆[22-23]。另外也有可能是由于不同时代的家系数目不同,个体数量不同或者家系构建周期较长造成。在以前的几项研究中,得到的大菱鲆生长性状遗传力均不同程度大于本研究的结果[24-26]。虽然不同的群体,不同的环境或者统计模型对于遗传力的大小有影响,但是笔者还是推断本研究中大菱鲆遗传力被低估了[27]。在选育计划中,基础种群(G0)是通过杂交不同来源的个体而建立的。在随后的世代中,为了获得可接受的近交系数并保持较高的有效种群规模,笔者在遗传分化个体之间进行了交配。交配实践可能会导致每代杂合度过高,在表型上可以表示为杂种优势。这可以解释缩小的遗传力估计值。本研究得到的收获体重遗传力表明该大菱鲆群体仍然有较大的遗传改良潜力。在目前的选育项目中,大菱鲆的繁殖种群具有可观的加性遗传效应变异,可用于进一步提高其生长性能。
3.2 共同环境效应
本研究就发现跨世代的共同环境效应较大(0.28±0.02),这主要是因为在大菱鲆家系培育中,每个家系单独养殖在玻璃缸桶中,直至达到100日龄时,个体规格可以满足VIE标记注射后,不同家系的个体通过VIE标记后才混养在一起进行性状测评,虽然在此过程中尽量保证不同家系养殖、管理的一致性,但仍旧不可避免地在不同家系间产生了共同环境效应。统计结果发现跨世代的标准误(遗传力和共同环境效应)均小于单个世代,说明大数据组对于遗传评估精确性影响较大。另外,对于单一世代,亲本和后代的遗传联系可能不足以准确区分遗传和环境效应,因此通过世代相传的父母之间的牢固遗传联系将提高遗传力估计的准确性。
3.3 遗传进展
研究结果中,平均估计育种值大小总体呈上升趋势,这表明针对大菱鲆体重性状开展的BLUP选育总体上是成功的。G0,G1世代选择差为正值,而G2世代的选择差为负值,这可能是由于G2世代由于环境因素,体重较高的个体死亡率较高(图1所示),导致选择差为负值。遗传选育提高了大菱鲆的生长性能,经过3代的选育累计实现的遗传进展为10.86%,平均每代遗传进展为3.56%,与早期的水产动物研究相比是一个相对较低的水平[28-29]。这种较低遗传进展结果可能是由于大菱鲆性腺发育率较低,这造成每一世代选育群体规模较小,只能进行较低强度的选择,特别是G3代遗传进展0.04%。在基础群体已经构建且估计遗传力等参数固定的情况下,育种过程中增加家系数量、扩大选育群体规模和提高选择强度可有效增加遗传进展[30]。而在大菱鲆选育过程中,世代周期较长,需3~4年才能性成熟,且性腺发育率较低(特别是雌鱼),这导致每一代只能进行较低强度的选择。在本研究中,不同年份所建立的全同胞家系数量和最终收获体重的最小二乘均值差异都较大,这些结果表明养殖环境和管理尚未达到最理想的水平。在后续的选育计划中,提高候选亲本的性腺发育率是增加选择强度,维持遗传多样性,提高遗传进展的基础。

图1 谱系树状图
4 结论
大菱鲆群体经过三代选育,遗传进展分别为10.2、37.4、0.3 g,分别相当于2.06%、8.76%、0.04%,累积遗传进展为10.86%,以上说明针对大菱鲆体重性状的BLUP选育是成功的。相比于基础群体,3个选育世代的体重性状遗传力并未出现下降,说明该选育群体仍具有选育潜力。
猜你喜欢
国外畜牧学·猪与禽(2024年4期)2024-01-01 00:00:00
养猪(2022年4期)2022-08-17 07:07:02
食品工业(2021年9期)2021-04-16 09:49:47
中国水产(2019年3期)2019-03-25 09:27:20
浙江农业学报(2016年7期)2016-06-15 20:30:11
湖北畜牧兽医(2015年11期)2016-01-11 10:08:24
地理与地理信息科学(2015年4期)2015-10-13 08:29:22
水利水电科技进展(2014年1期)2014-10-17 02:29:06
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28 12:21:37
河南科技(2014年8期)2014-02-27 14:08:06
