
瘤黑粉菌侵染对抗感玉米自交系苗期叶片的影响
2021-02-13 07:18张文洁李昕洋马育庭吕香玲杜万里
沈阳农业大学学报 2021年6期
张文洁,王 洋,李 阳,杨 悦,李昕洋,马育庭,吕香玲,杜万里
(1.黑龙江八一农垦大学农学院,黑龙江大庆163000;2.沈阳农业大学农学院,沈阳110161)
玉米瘤黑粉病是由玉蜀黍黑粉菌[Ustilagomaydis(DC.)Corda]侵染所致。玉蜀黍黑粉菌属于担子菌亚门,黑粉菌目,黑粉菌科,黑粉菌属,是一类依赖有性生殖而致病的营养型寄生性真菌[1]。我国玉米瘤黑粉病普遍发生在东北、华北、西北春玉米区及黄淮海夏玉米区,发病率一般为5%~10%[2]。大量研究表明,培育抗病品种提高遗传抗性是控制瘤黑粉病最经济有效的措施。
WGA-AF-488染色是检测瘤黑粉菌侵染程度的重要方法,可为发现病菌入侵寄主提供直接证据[3-5],超微结构观察可在亚细胞水平揭示寄主对病原菌侵入的反应[6-7]。当病原菌侵染时,不仅植物体组织细胞发生变化,可溶性糖含量也发生变化[8]。HORST等[9]对黑粉菌侵染1,2,4,8d时叶片代谢物含量进行分析,结果表明可溶性糖(葡萄糖、果糖、蔗糖)含量在不同侵染时期均高于对照,蔗糖含量呈持续增长趋势,果糖和葡萄糖含量在侵染8d时极显著高于侵染4d。Str1是一个对蔗糖具有高亲和力特异性的转运蛋白,仅在感染时表达,其缺失可显著降低真菌的毒力[10];LANVER等[11]对瘤黑粉菌进行转录组分析,发现瘤黑粉菌编码至少19个糖转运蛋白,且在瘤黑粉菌侵染期间,糖转运蛋白均表现为上调表达。以上研究表明,瘤黑粉菌侵染会导致可溶性糖大量积累,对真菌生长和肿瘤形成具有至关重要的作用。病原菌与寄主互作过程中,寄主将启动一系列防御反应,抗氧化酶通过清除过量活性氧维持细胞稳态就是重要的防御反应之一[12]。张骥如飞[13]研究发现糜子高抗黑穗病品种叶片中POD活性与对照相比大幅升高,感病品种与对照相比显著降低,SOD活性在叶片受侵染后最先响应,且高抗品种增幅大于高感品种。保护酶是植物抵抗病原菌侵染的重要标记物质,与抗病性呈正相关。
本研究以2个抗感瘤黑粉病玉米自交系为材料,通过激光共聚焦显微镜、体视显微镜、扫描电镜、透射电镜和生理物质检测比较抗、感自交系受瘤黑粉菌侵染后的组织、细胞和生理变化差异,研究结果可为解析玉米抗病防御途径和筛选抗病种质提供理论参考。
1 材料与方法
1.1 材料
供试瘤黑粉菌FB1×FB2由吉林农业科学研究院郑岩研究员提供。病原菌培养参照严理等[14]的方法。供试玉米(Zea mays L.)感病自交系为掖478;抗病自交系为齐319。
1.2 方法
待掖478和齐319播种后25d,利用10mL一次性注射器吸取OD600值为0.8~1.0的FB1×FB2菌液,采用人工注射法接种于玉米茎中上部,每株注射1mL直至菌液从心叶冒出即可。接种后0,2,4,8,12d取接种部位叶片,以接种蒸馏水为对照,接种FB1×FB2菌液为处理组,3次重复,用于组织、细胞和生理指标检测。
1.2.1 数据处理及抗病性评价 感病自交系掖478和抗病自交系齐319的病情指数及抗病性评价参照严理等[14]对玉米苗期瘤黑粉病的划分标准进行评价。
1.2.2 WGA-AF-488/PI染色观察 样品制备参照REDKAR等[15]的方法,处理后利用抗荧光淬灭封片剂封片,于激光共聚焦显微镜(ZEISS,LSM 780)下观察菌丝形态差异,WGA-AF-488激发波长488nm,发射光波长500~540nm;PI(碘化丙啶)的激发波长561nm,发射光波长580~630nm。
1.2.3 石蜡切片观察 石蜡切片方法参照王洋[16]的方法。利用体视显微镜(ZEISSAxio Zoom V16)观察抗、感自交系叶片组织细胞差异,利用ZEN2(blue edition)软件处理图像。
1.2.4 扫描电镜观察 取接菌部位2cm×2cm叶片,参照杨国一等[17]的方法并进行优化,经70%、80%、90%、95%、100%乙醇梯度脱水,无水乙醇∶叔丁醇=2∶1(V/V)、无水乙醇∶叔丁醇=1∶1(V/V)、无水乙醇∶叔丁醇=1∶2(V/V)、100%叔丁醇梯度置换,冷冻干燥仪(LyoQuest-85plus)冷冻干燥24h后置于真空镀膜机(Hitachi,MSP-2S)镀膜,扫描电镜(Hitachi,TM-3030)观察病菌侵染抗、感自交系叶片差异。
1.2.5 透射电镜观察 参照吕静波[18]的方法,经乙醇梯度脱水、无水丙酮置换,环氧树脂浸透、包埋、聚合后,使用莱卡超薄切片机(LEICA,EM UC7)切片,醋酸双氧铀和柠檬酸铅双重染色。透射电子显微镜(Hitachi,HT7700)观察抗、感自交系叶片超微结构差异。
1.2.6 可溶性糖含量与保护性酶活性测定 可溶性糖含量参照刘海英等[19]的方法进行测定。保护酶POD、SOD和CAT活性分别采用愈创木酚法[20]、氮蓝四唑还原法[21]和紫外吸收法[22]进行测定。
2 结果与分析
2.1 玉米自交系掖478和齐319的病情指数及抗病性评价
本试验连续3年对玉米自交系掖478和齐319进行人工接种鉴定,经田间表型统计分析得出,自交系掖478发病率为93.3%,病情指数84.2%,为高感自交系;自交系齐319发病率为1.3%,病情指数1.85%,为高抗自交系。
2.2 瘤黑粉菌侵染玉米苗期叶片的WGA-AF-488/PI染色观察
WGA-AF-488/PI染料可使瘤黑粉菌在激光共聚焦显微镜488nm波长下呈现绿色荧光(图1 A),PI可使叶片组织细胞在561nm波长下呈现红色(图1 B)。接种2d后,掖478表面菌株孢子萌发伸长形成侵染菌丝(图1 C);齐319叶片表面观察到绿色荧光信号,与对照菌液细胞比较明显伸长(图1 c)。接种4d后,掖478叶片菌丝大量增殖,菌丝出现不规则分支(图1 D);齐319叶片表面菌株孢子形态与接种2d后基本一致,未形成侵染菌丝(图1 d)。接种8d后,掖478叶片中菌丝大量增殖,分支原基与分支数量显著增加(图1 E);齐319叶片形成侵染菌丝(图1 e)。接种12d后,掖478叶片接种部位被菌丝覆盖(图1 F);齐319侵染菌丝分支数量增加,长度伸长(图1 f)。
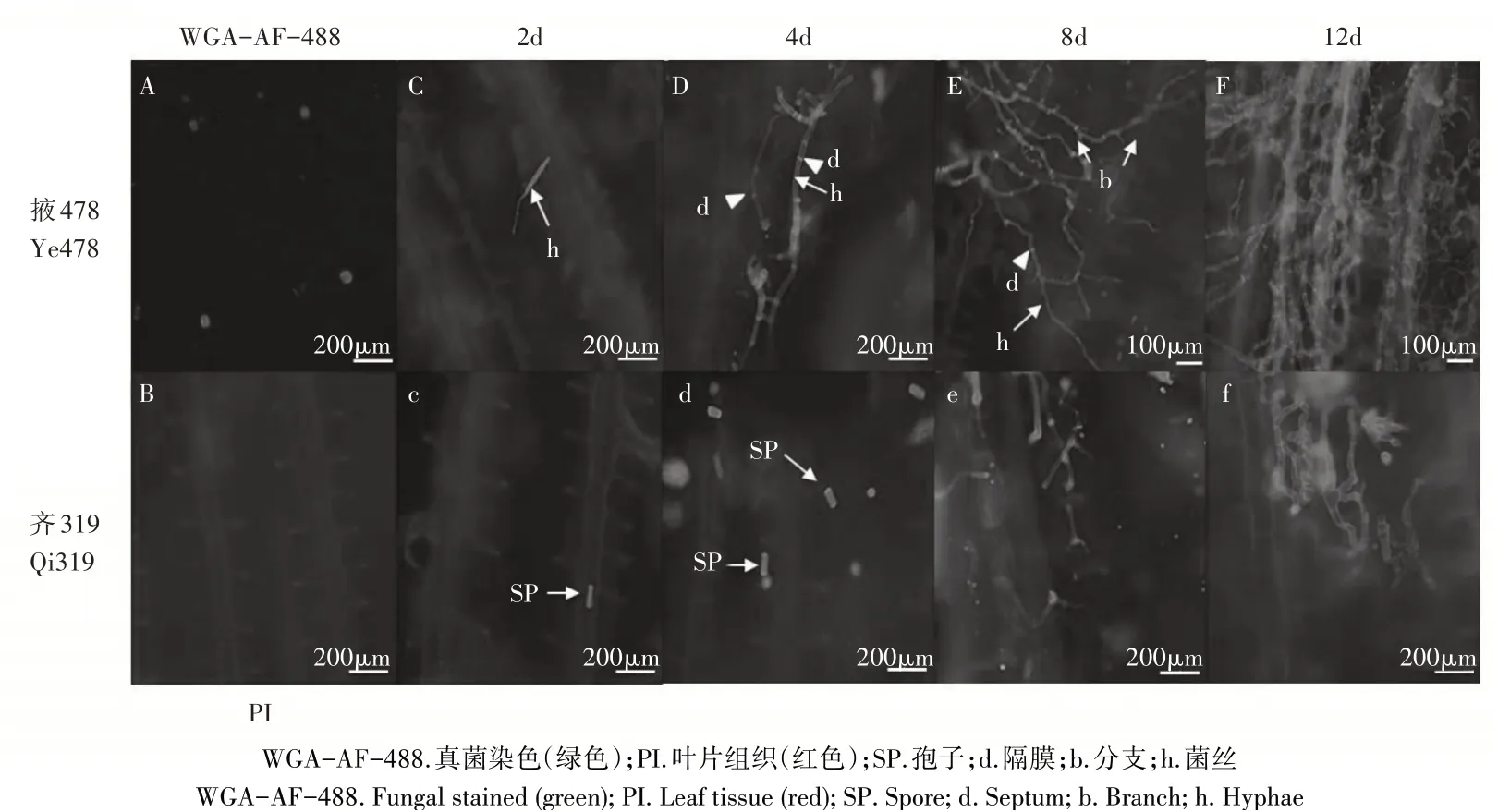
图1 瘤黑粉菌侵染感病玉米自交系掖478和抗病玉米自交系齐319叶片的WGA-AF-488/PI染色Figure 1 WGA-AF-488/propidium iodide co-stained susceptive and resistant maize inbred line Ye478 and Qi319 leaves infected with Ustilago maydis
2.3 瘤黑粉菌侵染玉米苗期叶片的石蜡切片观察
接种0~2d时,掖478和齐319叶片花环结构完整,维管束鞘细胞和叶肉细胞结构清晰(图2 A、图2 a、图2 B和图2 b)。接种4d,掖478维管束鞘细胞数量增多,叶肉细胞数量无明显变化,花环结构膨胀(图2 C);齐319维管束鞘细胞和叶肉细胞数量无显著变化(图2 c)。接种8d,掖478叶片维管束鞘细胞分裂加剧,叶肉细胞体积明显增大,花环结构模糊(图2 D);齐319叶肉细胞和维管束鞘细胞逐渐肿胀(图2 d)。接种12d,掖478叶片形成结节状凸起,组织结构完全被破坏,维管束鞘细胞形态消失(图2 E);齐319叶片叶肉细胞无明显变化,维管束鞘细胞继续分裂,花环结构保持完整(图2 e)。为明确接种12d掖478凸起囊状物的成分,通过淀粉-碘化钾染色发现,凸起囊状物积累了大量淀粉(图2 F)。
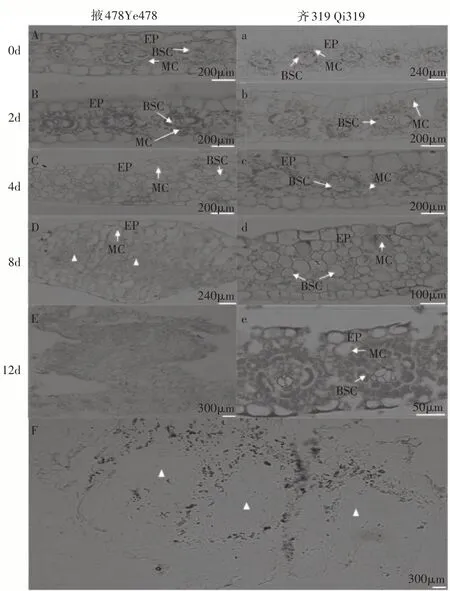
图2 瘤黑粉菌侵染感病玉米自交系掖478(A~F)和抗病玉米自交系齐319(a~e)叶片的石蜡切片观察Figure 2 The susceptible and resistant maize inbred line Ye478(A-F)and Qi319(a-e)leaves was investigated by paraffin section infected with Ustilago maydis
利用正置荧光显微镜观察叶片维管束鞘细胞结构(图3),接种4d,掖478维管束鞘细胞开始分裂,形成新的细胞,维管束鞘细胞数量增多;齐319维管束鞘细胞未见异常(图3 A和图3 a)。接种8d,掖478维管束鞘细胞进一步分裂,叶肉细胞出现明显增长;齐319维管束鞘细胞出现分裂现象(图3 B和图3 b)。接种12d时,掖478分生细胞持续分裂导致细胞数量显著增加,且叶肉细胞进一步膨胀;齐319维管束鞘细胞分裂迟缓(图3 C和图3 c)。
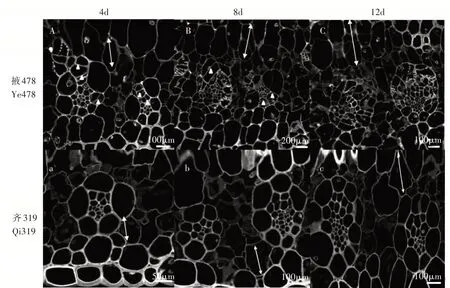
图3 瘤黑粉菌侵染感病玉米自交系掖478(A~C)和抗病玉米自交系齐319(a~c)叶片的维管束鞘分析Figure 3 Analysis of vascular bundle sheath on leaves of susceptible and resistant maize inbred line Ye478(A-C)and Qi319(a-c)after inoculation Ustilago maydis
2.4 瘤黑粉菌侵染玉米苗期叶片的扫描电镜观察
基于抗感自交系WGA-AF-488鉴定结果,本试验利用扫描电镜进一步观察瘤黑粉菌侵染抗、感自交系的动态过程。接种0d,掖478和齐319叶片表面、内部均无菌丝,气孔清晰可见(图4 A、图4 A-1、图4 a和图4 a-1)。接种2d,掖478叶片表面可观察到菌丝端部形成附着胞,穿透寄主细胞,叶肉组织可见少量菌丝(图4 B和B-1);齐319叶片表面仅观察到菌株孢子,且叶肉组织无菌丝出现(图4 b和图4 b-1)。接种4d,掖478叶片表面菌丝伸长,与寄主的共生关系建立(图4 C),叶片内部菌丝扩展伸长,数量增多(图4 C-1);齐319叶片表面菌株孢子形态未发生显著变化(图4 c),叶片内部未观察到菌丝(图4 c-1)。接种8d,掖478叶片表面菌丝长度持续伸长(图4 D),叶片内部可见大量分支菌丝生长(图4 D-1);齐319叶片表面观察到菌丝伸长(图4 d),叶片内部仍未观察到菌丝(图4 d-1)。接种12d,掖478叶片表皮附着大量菌丝,叶片内部菌丝大量增殖并产生次生分支,逐渐向周围细胞入侵(图4 E和图4 E-1);齐319叶片表面菌丝进一步伸长(图4 e),叶片内部可见菌丝跨细胞侵染(图4 e-1)。

图4 瘤黑粉菌侵染感病玉米自交系掖478和抗病玉米自交系齐319叶片的扫描电镜观察Figure 4 SEM observation on the leaves of susceptible and resistant maize inbred line Ye478 and Qi319 infected with Ustilago maydis
2.5 瘤黑粉菌侵染玉米苗期叶片的超微结构观察
利用超薄切片技术探究抗、感自交系苗期不同时间节点叶片维管束鞘细胞中各细胞器变化(图5)。接种0d和2d,掖478和齐319细胞结构完整,叶绿体均匀地分布在维管束鞘细胞中,叶绿体内椭圆形淀粉粒清晰可见,且叶绿体基质片层有序排列,线粒体被膜完整,内嵴结构清晰(图5 A1、图5 B1、图5 a1和图5 b1)。接种4d,掖478细胞中淀粉粒大量积累,椭圆形淀粉粒逐渐向圆形转变(图5 C1),导致叶绿体形态肿胀(图5 C),叶绿体内基质片层出现断裂(图5 C1-1),线粒体内嵴结构模糊(图5 C1-2);齐319细胞叶绿体中淀粉粒数目增多(图5 c1),叶绿体基质片层完整清晰、无断裂现象(图5 c1-1),线粒体结构无显著变化(图5c1-2)。接种8d,叶绿体逐渐降解,出现质壁分离(图5 D),叶绿体中淀粉粒数量减少并随之解离(图5 D1),叶绿体内基质片层结构模糊(图5 D1-1),线粒体形态改变(图5 D1-2);齐319细胞结构完整(图5 d),叶绿体中淀粉粒含量增加,基质片层结构完整(图5 d1和图5 d1-1),线粒体增多,内嵴清晰(图5 d1-2)。接种12d,掖478维管束鞘细胞中细胞器彻底溶解消失,细胞边界无法辨识(图5 E);齐319叶绿体变化明显(图5 e),叶绿体基质片层结构清晰完整,淀粉粒数量与接种8d时相比显著降低(图5 e1和图5 e1-1),线粒体数量明显增多,呈聚集性排列并伴有空泡化现象(图5 e1-2)。

图5 瘤黑粉菌侵染感病玉米自交系掖478(A~E)和抗病玉米自交系齐319(a~e)叶片的透射电镜观察Figure 5 TEM observation on the leaves of susceptible and resistant maize inbred lines Ye478(A-E)and Qi319(a-e)infected with Ustilago maydis
2.6 玉米抗感自交系叶片的可溶性糖含量和保护酶活性变化
玉米瘤黑粉菌侵染2,4,8,12d掖478可溶性糖含量逐渐增加,侵染12d含量最高。齐319可溶性糖含量在侵染2,4,8d时均呈上升趋势,侵染12d含量下降,且显著低于掖478(图6 A)。抗、感自交系的SOD活性均呈现先升高,侵染12d下降的趋势,且齐319的SOD活性始终高于掖478(图6 B)。随瘤黑粉菌侵染天数的增加,抗、感自交系的POD和CAT活性均呈现稳步上升的趋势,且齐319高于掖478,侵染4d齐319的POD活性显著激增,侵染12dPOD和CAT活性达到峰值(图6 C和图6D)。
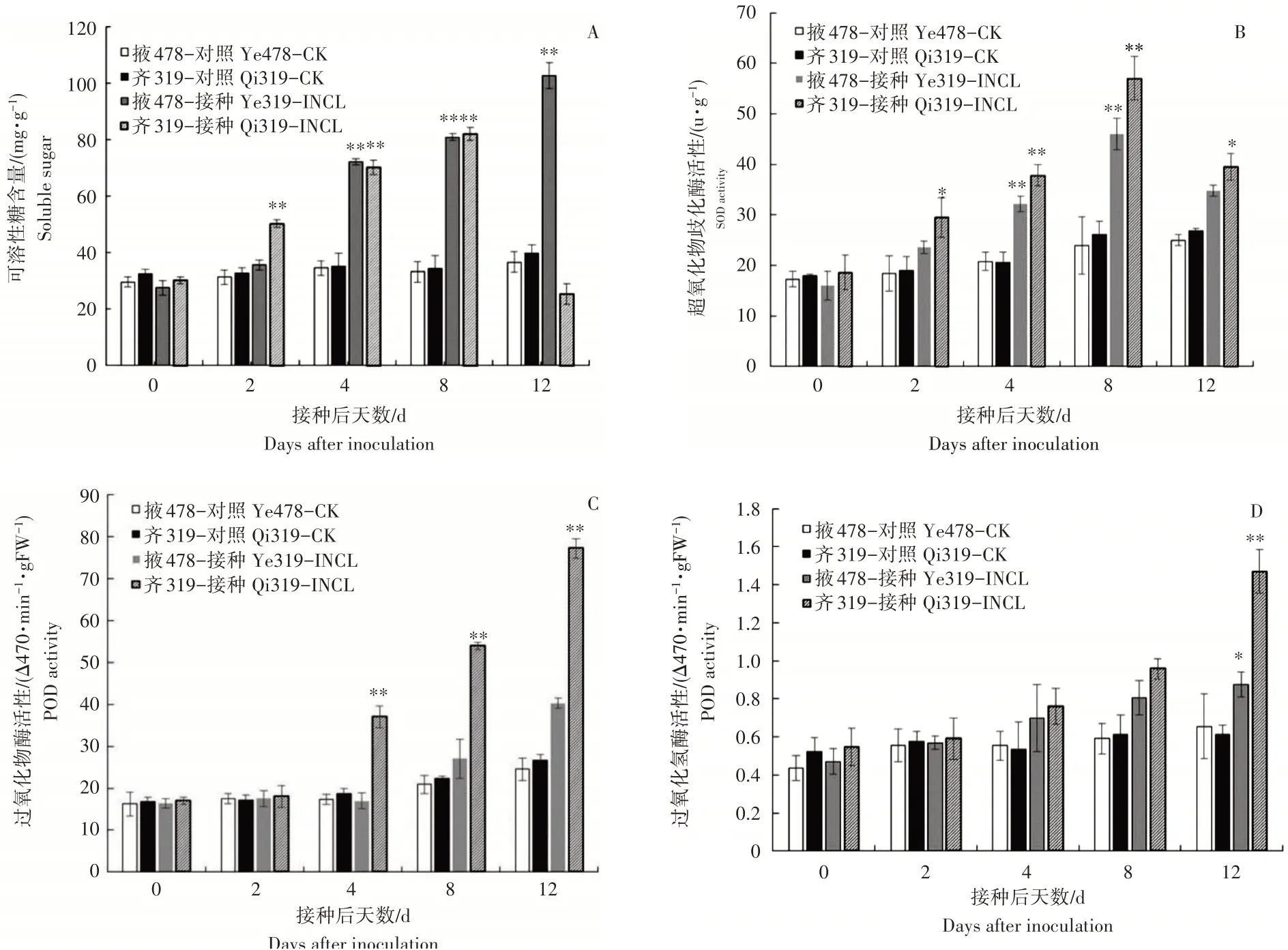
图6 瘤黑粉菌侵染抗感玉米叶片的可溶性糖含量和保护性酶活性变化Figure 6 Changes of soluble sugar content and defensive enzymes activity in susceptible and resistant maize leaves infected with Ustilago maydis
3 讨论与结论
Alexa Flour 488(AF 488)标记的麦胚凝集素(WGA)荧光染料可对黑粉菌进行特异性识别,与黑粉菌丝细胞质成分特异性结合,被广泛应用于黑粉菌侵染寄主的动态观察,如瘤黑粉菌侵染玉米花药[3]、小麦腥黑粉菌侵染小麦子房[4],本研究利用WGA-AF-488/PI对抗、感自交系掖478和齐319进行染色鉴定,清晰观察到抗、感自交系中菌丝生长发育差异。石蜡切片是研究病原菌侵染植物,引起组织结构变化的常用方法。MATEI等[6]对瘤黑粉菌侵染形成肿瘤的叶片进行淀粉-碘化钾染色,表明肿瘤部位聚集了大量淀粉。本研究利用石蜡切片方法解析抗、感自交系的组织细胞差异,结果表明掖478叶肉细胞逐渐膨大,维管束鞘细胞快速分裂,细胞数量急剧增多,最终形成囊状物,齐319维管束鞘细胞数量增长较慢,叶肉细胞逐渐肿胀,花环结构完整,进一步通过淀粉-碘化钾染色,发现侵染12d囊状物部位的淀粉含量明显高于正常叶片部位,研究结果与MATEI等(2018)一致。组织和细胞观察表明,抗、感自交系主要差异在维管束鞘细胞,推测齐319可能通过抑制菌丝向维管束鞘细胞侵染而形成抗病机制,掖478则无法有效抑制维管束鞘细胞的分裂。扫描电镜在探究病原菌入侵机制中发挥重要作用,如在水稻抗白叶枯病[23]和玉米瘤黑粉病[24]研究中被广泛应用。本研究利用扫描电镜观察,发现掖478和齐319叶片表面菌丝聚集,形成附着胞,穿透叶片表皮细胞壁进入细胞内的动态变化(图4)与上述WGA和石蜡切片研究结果一致。玉米瘤黑粉菌穿透玉米叶片组织过程中,菌丝最可能通过细胞壁降解酶的分泌逐渐穿透表皮细胞进入叶片组织[25],由此可见瘤黑粉菌对抗感玉米自交系的入侵方式相同,但病菌在叶片上表皮细胞壁的识别过程中机制可能不同,因而存在抗性差异,本结果明确了病原菌对不同抗性材料侵染动态的差异,可为瘤黑粉菌侵染玉米不同抗性材料研究提供参考。透射电镜在研究病原菌与寄主互作、寄主超微结构变化中发挥着重要作用[26]。MATEI等[6]利用透射电镜观察感病材料叶绿体的超微结构,发现叶绿体在类囊体膜之间大量积累淀粉粒,导致细胞器严重变形。本研究利用透射电镜技术,观察到瘤黑粉菌侵染掖478后,维管束鞘细胞中叶绿体由椭圆形膨胀为圆形,淀粉粒大量积累,最终各细胞器溶解消失,该结果与石蜡切片观察维管束鞘细胞分裂扩张导致花环结构消失一致(图3,图5D)。齐319细胞始终保持完整,但侵染后期淀粉粒数量降低,并出现大量线粒体(图5e)。因线粒体是能量代谢的主要场所,糖类是植物生长发育主要能量供应者,推测齐319在受到瘤黑粉菌侵染时可能通过大量的能量代谢抵抗病原菌侵染,淀粉粒的缺失、转化可能为线粒体维持细胞代谢提供主要能量。
糖类是异养微生物生长、繁殖的物质基础之一,瘤黑粉菌冬孢子加入己糖和蔗糖后更易萌发[27]。研究表明瘤黑粉侵染的叶片己糖含量升高,在玉米幼苗侵染过程中,3种ZMSWEETs糖转运蛋白表达上调[25,28]。本研究显示玉米苗期叶片受到瘤黑粉菌侵染后,齐319和掖478的可溶性糖含量均呈上升趋势,侵染12d掖478可溶性糖含量升至最高值,齐319则明显下降。瘤黑粉菌侵染掖478 4d时糖含量升高与透射电镜淀粉粒数量增多结果一致(图5),侵染12d可溶性糖含量显著升高且达到最大值与石蜡切片碘化钾-淀粉染色结果一致(图2),表明掖478糖含量的升高为瘤黑粉菌侵染提供了物质保障,齐319在侵染12d可溶性糖含量骤然降低与透射电镜维管束鞘细胞淀粉粒数量骤然减少相一致,糖含量的减少可能是植物抵御病原菌侵染的重要表现。因此推测玉米瘤黑粉病抗性与糖类物质合成和代谢转运相关。病原菌与寄主互作时会通过寄主组织中活性氧物质的大量积累和植物抗性相关酶类迅速变化来提高植物抗性[29]。本研究中瘤黑粉菌侵染后,保护酶活性迅速增加,且齐319始终高于掖478,证明齐319在玉米瘤黑粉菌侵染后通过清除活性氧而大量积累保护酶,以抵抗病原菌侵染。
本研究结果明确了瘤黑粉菌入侵后,感病材料菌丝快速增殖,通过维管束鞘细胞分裂和叶肉细胞膨大形成肿瘤,可溶性糖含量逐渐升高。在抗病材料中菌丝生长受抑制,穿透细胞表皮侵染受阻,花环结构完整清晰,可溶性糖含量降低,保护酶活性逐渐升高且始终高于感病材料。研究结果明确了抗、感病玉米自交系苗期叶片响应瘤黑粉菌侵染的组织、细胞和保护酶活性差异,可为进一步揭示瘤黑粉病的侵染机制,筛选抗病种质提供理论参考。
猜你喜欢
当代水产(2022年1期)2022-04-26
河北农业大学学报(2021年5期)2021-11-10
新疆农业科学(2020年9期)2020-10-13
河南农业科学(2019年9期)2019-09-24
北京农学院学报(2019年1期)2019-02-22
河南农业科学(2017年4期)2017-04-12
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
西南农业学报(2016年6期)2016-04-16
