
极北海带幼苗对氮、磷和铁营养元素需求的研究
2021-01-28 10:53杜欣欣刘福利袁艳敏梁洲瑞张朋艳刘升平
水产科学 2021年1期
杜欣欣,刘福利,袁艳敏,梁洲瑞,张朋艳,刘升平
(1.青岛农业大学 海洋科学与工程学院,山东 青岛 266109; 2.农业农村部海洋渔业 资源可持续利用重点开放实验室,中国水产科学研究院 黄海水产研究所,山东 青岛 266071; 3.青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛 266000 )
极北海带(Laminariahyperborea),属褐藻门、褐藻纲、海带目、海带科、海带属,是一种冷水性海藻,分布于北大西洋东北海区,北起斯堪的纳维亚半岛,南至加那利群岛,包括波罗的海、北海沿岸[1-3]。极北海带个体较大,长度一般为2~4 m,是多年生的海藻,寿命一般为10~20 a[4-6]。极北海带通常生长于潮下带的硬质基质上,根据海水透明度不同,其垂直分布的下限为海平面下10~35 m。在北大西洋东北海区临近大潮低潮线下底质为基岩的区域,极北海带是优势种,常与其他大型褐藻[糖海带(L.saccharina)等]形成“海藻森林”[7-8],可成为海洋动物的索饵场、产卵场和栖息地,其也对浅海生态环境的改善、海洋生物多样性的增加具有积极作用[8-9]。
海藻苗种培育是养殖业发展的前提,通过建立极北海带配子体繁育技术体系,可以实现人工增养殖的需要,但对其幼苗培养过程中营养盐的需求尚不清楚。国内外对海藻吸收氮、磷营养盐方面已有较多研究。王翔宇等[10]对6种大型海藻吸收营养盐的能力进行了研究;李文慧[11]分析了氮、磷营养盐因子对缘管浒苔(Enteromorphalinza)生长、叶绿素荧光特性和氮磷富集的影响;许忠能等[12]研究了环境因子对细基江蓠繁枝变种(Gracilariatenuistipitatavar.liui)氮、磷吸收速率的影响。氮、磷和铁是限制海藻生长的主要元素,这些元素在海水中的含量变化较大,其在海藻内的含量远高于海水中的含量,但对大型海藻中氮、磷、铁间的相互影响作用尚不清楚[13]。目前国内外对氮、磷营养元素的研究较多[14-16]。海藻对不同氮营养盐的利用效率不同,海藻对NH4+-N的吸收率大于对NO3--N的吸收率。田千桃等[17]发现,2种氮源同时存在时,浒苔(E.prolifera)对NH4+-N的吸收速率高于对NO3--N的吸收速率。吕冬伟等[18]以孔石莼(Ulvapertusa)为材料,研究其对水体中氮磷营养盐吸收情况时发现,孔石莼对NH4+-N和PO43--P的吸收效果明显高于NO3--N。铁也是海藻生长的主要限制性营养元素之一[19-23],其既能参与海藻光合及呼吸过程,又能参与光合和呼吸过程中的光合磷酸化和氧化磷酸化[21,24-25]。朱明远等[26]探讨了铁(EDTA-Fe)对三角褐指藻(Phaeodactylumtricornulum)生长、光合作用及生化组成的影响;陈静峰等[27]研究了磷、铁盐的协同作用对赤潮隐藻(Crytomonassp.)生长的影响;姚波等[28]综述了铁限制对浮游植物生长和群落组成的影响,认为铁是浮游植物生长的限制性因素,铁含量的改变会引起群落结构的变化。但关于铁离子对大型海藻尤其是海带方面的研究较少。
笔者通过表征氮、磷、铁在不同质量浓度条件下的生长速度和光合生理参数,探明极北海带幼苗生长所需要的适宜氮、磷、铁用量,为极北海带幼苗的人工培育提供依据。
1 材料与方法
1.1 试验材料
所用极北海带幼苗是通过配子体繁育技术所得,选取3~5 cm、无褶皱损伤、无病害的极北海带幼苗作为材料。幼苗培育条件为:充气培养,温度11~13 ℃,光合有效照射度(PAR)为40 μmol/(m2·s),光周期为12L∶12D,使用天然过滤灭菌海水(氮0.1 mg/L,磷0.01 mg/L,铁0.002 mg/L)作为培养液,添加营养盐质量浓度为NO3--N:4 mg/L,PO43--P:0.4 mg/L。培养液更换1次/3 d。试验开始前,从挑选出的极北海带幼苗中再次选择,使得试验各处理组平均质量为(1±0.05) g,质量差异性不显著(P>0.05)。
1.2 试验方法
1.2.1 不同NO3--N、PO43--P质量浓度对极北海带幼苗生长和光合生理的影响
以过滤、灭菌的天然海水作为培养液,以NaNO3和KH2PO4为氮源和磷源,设置7个不同氮、磷质量浓度[ρ(N)∶ρ(P)=10∶1]:NO3--N为0、0.5、2、4、6、8、10 mg/L。其他培养条件:光合有效照射度为40 μmol/(m2·s),光周期为12L∶12D,温度9 ℃,每隔2 d更换一次培养液。每个处理组设置3个对照。
称量每个处理组初始幼苗的质量,7 d后再次称量质量,计算相对生长速率(vRGR),计算公式为:
式中m0为初始藻的质量(g),mt为试验结束时藻的质量(g),t为试验持续的时间(d)。
在上述各质量浓度条件下培养幼苗12、24、36、48、60 h,然后测定其叶绿素荧光参数(光系统Ⅱ最大荧光产量,Fv/Fm)及培养12、24、48 h后幼苗的表观光合速率。使用水下饱和脉冲叶绿素荧光仪(DIVING-PAM,WALZ,Germany) 测定Fv/Fm值,使用液相氧电极(Hansatech Oxygraph,英国)测定表观光合速率。
1.2.2不同Fe3+质量浓度对极北海带幼苗生长及氮磷需求的影响
以NaNO3和NH4Cl为氮源,KH2PO4为磷源,FeCl3为铁源。设置不同氮、磷、铁的组合[ρ(N)∶ρ(P)=10∶1]。NH4+-N组设置2个Fe3+质量浓度(NH4+-N为3 mg/L):0、1 mg/L。NO3--N组设置4个Fe3+质量浓度(NO3--N为3 mg/L):0、0.2、1.0、5.0 mg/L。分别测定在上述各质量浓度条件下培养0、4、10、24、48 h后幼苗培养液中的PO43--P、NO3--N、NH4+-N和Fe3+质量浓度以及处理3 d后的极北海带幼苗的相对生长速率。每次试验设置3个平行组。
1.3 数据处理
采用SPSS 18.0数据统计软件进行单因素方差分析、Duncan多重比较,以P<0.05作为显著性差异,使用Prism软件绘制图形。
2 结 果
2.1 不同NO3--N、PO43--P质量浓度对极北海带幼苗生长和光合生理的影响
2.1.1 不同NO3--N、PO43--P质量浓度下极北海带幼苗的相对生长速率
方差分析结果显示,不同NO3--N、PO43--P质量浓度对极北海带幼苗的相对生长速率有显著性影响。在一定质量浓度范围内,极北海带幼苗的相对生长速率随着NO3--N、PO43--P质量浓度的升高而升高(图1),NO3--N:2、4、6、8 mg/L组的相对生长速率较高,且相互之间差异不显著(P>0.05)。NO3--N:0、0.5、10 mg/L的相对生长速率较低,且显著低于NO3--N:2、4、6、8 mg/L组(P<0.05),说明NO3--N质量浓度2~8 mg/L是极北海带生长的适宜质量浓度范围,氮、磷质量浓度过高和过低均不利于极北海带生长。
2.1.2 不同NO3--N、PO43--P质量浓度下极北海带幼苗的叶绿素荧光参数
极北海带的光合效率(Fv/Fm值)随着NO3--N、PO43--P质量浓度的升高呈现先升后降的趋势(图2)。由NO3--N:2 mg/L组到NO3--N:4 mg/L组,极北海带的Fv/Fm值逐渐上升,当达到NO3--N:4 mg/L时,Fv/Fm值达到最高值,此后随着质量浓度增加,Fv/Fm值呈现下降趋势。NO3--N:2、4 mg/L时的Fv/Fm值显著高于其他NO3--N、PO43--P质量浓度时的Fv/Fm值,说明NO3--N质量浓度在2~4 mg/L时,极北海带具有高水平的光合效率。另外,对于添加适量NO3--N、PO43--P营养盐的处理组,不同培养时间的Fv/Fm值差异不显著,而对于NO3--N:10 mg/L组,随着培养时间延长,Fv/Fm值显著升高。

图2 不同NO3--N、PO43--P质量浓度下处理12、24、36、48、60 h后极北海带幼苗的Fv/FmFig.2 Fv/Fm values of juvenile kelp L. hyperborea exposed to different NO3--N and PO43--P concentrations for 12 h, 24 h, 36 h, 48 h and 60 h 字母下标为相同数字的作为同一组,进行多重比较,同一组的不同字母表示在P<0.05水平具有显著性差异,下同. Fv/Fm values with the same letters in the histogram are within the same group, and the means with different letters in the same group are significant difference at the level of P<0.05, et sequentia.
2.1.3 不同NO3--N、PO43--P质量浓度下极北海带幼苗的表观光合速率
在不同的NO3--N、PO43--P质量浓度下(除NO3--N:0 mg/L处理12 h外),极北海带的表观光合速率具有先升后降的趋势,在NO3--N:2、4 mg/L组时数值较高(图3)。通过相同时间不同质量浓度之间比较发现,NO3--N:2、4 mg/L组在24、48 h的表观光合速率较大,与NO3--N:8、10 mg/L组具有显著性差异(P<0.05),12 h之内各组与NO3--N:0 mg/L组之间均差异不显著(P>0.05);通过相同质量浓度不同时间之间比较发现,NO3--N:0 mg/L组处理12 h显著大于处理48 h(P<0.05),NO3--N:10 mg/L组在12 h的表观光合速率较大,并且与24、48 h之间具有显著性差异(P<0.05),其余各组在48 h之内均无显著性差异(P>0.05)。
2.2 不同Fe3+质量浓度对极北海带幼苗生长及氮磷需求的影响
2.2.1 不同Fe3+质量浓度对极北海带幼苗相对生长速率的影响
NO3--N-Fe3+:1 mg/L组幼苗的相对生长速率最大(图4),显著大于NH4+-N-Fe3+:0 mg/L和NH4+-N-Fe3+:1 mg/L组(P<0.05),且显著大于NO3--N-Fe3+:0 mg/L和NH4+-N-Fe3+:5 mg/L组(P<0.05)。NO3--N-Fe3+:1 mg/L和NO3--N-Fe3+:0.2 mg/L组间无显著性差异(P>0.05)。NO3--N-Fe3+:5 mg/L组相对生长速率最小,显著小于其他各组(P<0.05),说明高质量浓度Fe3+会抑制极北海带生长。
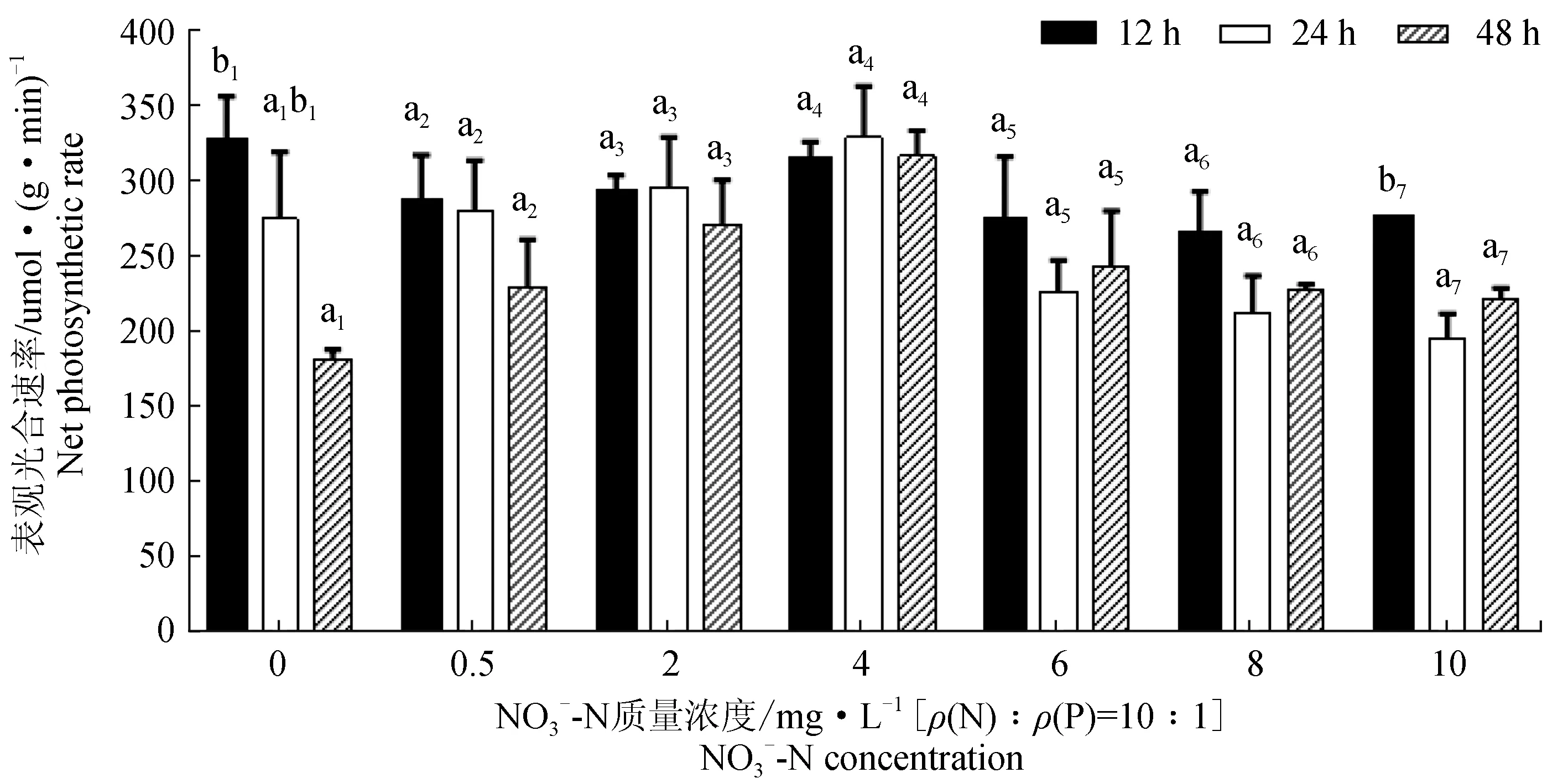
图3 不同NO3--N、PO43--P质量浓度处理12、24 h及48 h后极北海带幼苗的表观光合速率 Fig.3 The net photosynthetic rates of juvenile kelp L. hyperborea xposed to different NO3--N and PO43--P concentrations for 12 h, 24 h and 48 h

图4 不同Fe3+质量浓度处理3 d后极北海带幼苗的相对生长速率 (PO43--P均为0.3 mg/L)Fig.4 Relative growth rate (RGR) of juvenile kelp L. hyperborea cultivated under different concentrations of Fe3+ (PO43--P concentration of 0.3 mg/L) for 3 days
2.2.2 极北海带幼苗对不同Fe3+质量浓度的需求
极北海带对Fe3+的吸收主要在0~10 h之间(图5),单因素方差分析结果表明,Fe3+:0.2、1.0、5.0 mg/L组中Fe3+质量浓度在0 h与4 h之间差异显著(P<0.05),且均显著高于10、24、48 h(P<0.05);Fe3+:0.2 mg/L、1.0 mg/L组Fe3+质量浓度在10、24、48 h间均无显著性差异(P>0.05),Fe3+:5 mg/L组的Fe3+质量浓度在48 h时显著低于10 h和24 h(P<0.05)。因此,在0~5 mg/L范围内,Fe3+质量浓度越高,极北海带幼苗对Fe3+的吸收越多,且吸收作用主要在10 h内完成,随后吸收呈缓慢和稳定趋势。
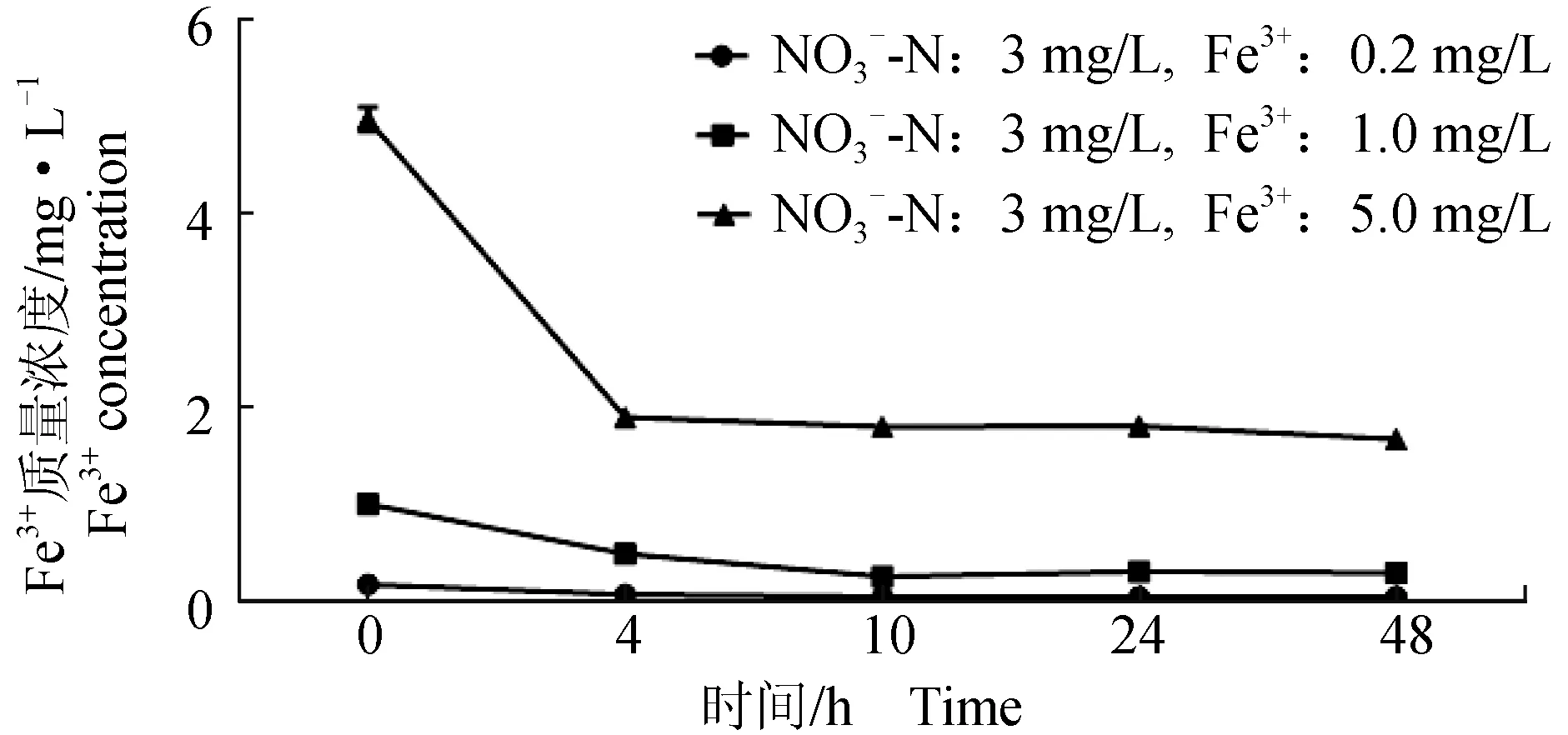
图5 极北海带幼苗培养液中的Fe3+质量浓度Fig.5 Fe3+ concentration in seedling culture solution of juvenile kelp L. hyperborea PO43--P均为0.3 mg/L,下同. PO43--P concentration of 0.3 mg/L, et sequentia
2.2.3 不同质量浓度Fe3+对极北海带幼苗NO3--N需求的影响
比较不同质量浓度间的处理,4 h时Fe3+:1 mg/L组NO3--N含量显著低于其他3组(P<0.05)(图6)。10 h时Fe3+:0 mg/L组NO3--N含量显著高于其他3组(P<0.05),Fe3+:0.2 mg/L组显著高于Fe3+:1 mg/L组(P<0.05),其余各组之间均无显著性差异(P>0.05)。24 h时各组间无显著性差异(P>0.05)。48 h时Fe3+:5 mg/L组NO3--N含量显著高于Fe3+:0.2 mg/L(P<0.05),其余各组间无显著性差异(P>0.05)。通过相同质量浓度不同时间之间的比较发现,Fe3+:0 mg/L组和Fe3+:0.2 mg/L组的4、10 h与24 h、48 h间NO3--N含量有显著性差异(P<0.05),且4 h与10 h含量显著高于24 h与48 h(P<0.05)。Fe3+:1 mg/L和Fe3+:5 mg/L组24 h与48 h间NO3--N含量无显著性差异(P>0.05),且含量显著低于4、10 h(P<0.05)。由此可见,Fe3+对极北海带吸收NO3--N具有促进作用,在0~10 h,且Fe3+质量浓度为0.2~1.0 mg/L时较为合适。

图6 不同Fe3+质量浓度条件下极北海带幼苗培养液中的 NO3--N含量Fig.6 The NO3--N concentration in culture medium of juvenile kelp L. hyperborea exposed to different concentrations of Fe3+
2.2.4 不同质量浓度Fe3+对极北海带幼苗NH4+-N需求的影响
与NO3--N相比,NH4+-N更易被吸收(图7),在4~48 h内培养液中NH4+-N的含量均显著低于同时期培养液中NO3--N的含量(P<0.05),NH4+-N-Fe3+:0 mg/L组在48 h时含量显著低于NH4+-N-Fe3+:1 mg/L组(P<0.05),0~24 h时,差异不显著(P>0.05)。
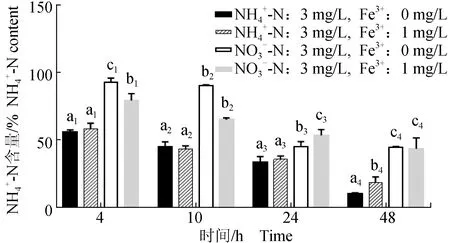
图7 不同Fe3+质量浓度条件下极北海带幼苗培养液中 NH4+-N的含量Fig.7 The NH4+-N concentration in culture medium of juvenile kelp L. hyperborea exposed to different concentrations of Fe3+
2.2.5 不同质量浓度Fe3+对极北海带幼苗PO43--P需求的影响
在相同温度、NO3--N、PO43--P条件下,加入不同质量浓度Fe3+后,培养液中PO43--P含量变化(图8)。通过相同时间不同质量浓度之间的比较发现,4 h时Fe3+1 mg/L组PO43--P含量显著低于其他3组(P<0.05);10 h时Fe3+1 mg/L组PO43--P含量显著低于Fe3+0、5 mg/L组(P<0.05),Fe3+0.2 mg/L组显著低于Fe3+5 mg/L组(P<0.05);24 h时各组的PO43--P含量均无显著性差异(P>0.05);48 h时Fe3+5 mg/L组PO43--P含量与其余各组具有显著性差异(P<0.05),其余各组间无显著性差异(P>0.05)。通过相同质量浓度不同时间之间的比较发现:Fe3+0 mg/L、0.2 mg/L组在48 h之内的PO43--P含量均有显著性差异(P<0.05);Fe3+:1 mg/L组10、24 h间的PO43--P含量无显著性差异(P>0.05),其他组之间均有显著性差异(P<0.05);Fe3+5 mg/L组24、48 h间的PO43--P含量无显著性差异(P>0.05),但均显著低于4、10 h时的PO43--P含量(P<0.05)。1 mg/L左右的Fe3+在24 h之内能促进极北海带对PO43--P的吸收,24 h后促进效果不明显。高质量浓度的Fe3+在10 h后对极北海带吸收PO43--P可能有抑制作用。

图8 不同Fe3+质量浓度条件下极北海带幼苗培养液中 PO43--P的含量Fig.8 The PO43--P concentration in culture medium of juvenile kelp L. hyperborea exposed to different concentrations of Fe3+
3 讨 论
3.1 不同NO3--N、PO43--P质量浓度对极北海带幼苗生长和光合生理的影响
随着NO3--N、PO43--P质量浓度的升高,极北海带的相对生长速率逐渐升高,并在NO3--N质量浓度为4 mg/L、PO43--P质量浓度为0.4 mg/L时达到峰值。这是因为氮是海藻蛋白质的重要组成部分[29-30],磷能促进海藻生殖细胞的形成和酶的活动与转化[31-32],二者是海藻生长的重要影响因子[33-36]。随着NO3--N、PO43--P质量浓度的升高,其相对生长速率逐渐下降。王萍等[37]探究了营养盐含量对繁枝蜈蚣藻(Grateloupiaramosissima)和孔石莼吸收氮、磷的影响, 结果显示,其对氮、磷的吸收随营养盐含量升高而增加。当氮、磷质量浓度过高时,则会产生O22-、OH-、H2O2等自由基,这些自由基则会引发膜脂过氧化,破坏细胞膜[38],从而影响海藻的生长。因此在高NO3--N、PO43--P质量浓度时,极北海带的相对生长速率下降。
Fv/Fm是衡量植物光合作用效率的重要指标[39-41]。当NO3--N质量浓度在2~4 mg/L时,极北海带Fv/Fm的值显著高于其他NO3--N、PO43--P营养盐组,这说明其在NO3--N、PO43--P营养盐质量浓度范围内具有较高的光合作用效率。当NO3--N质量浓度由0.5 mg/L升至4 mg/L[ρ(N)∶ρ(P)=10∶1],其表观光合速率随之上升,在该营养盐质量浓度下极北海带的表观光合速率最大。NO3--N:0.5 mg/L、PO43--P:0.05 mg/L时,表观光合速率的数值较低,是因为外源营养盐供应不足,降低了极北海带对营养盐的吸收,进而影响光合作用的酶和色素的合成[42]。综上所述,NO3--N在2~4 mg/L时极北海带具有相对较高的光合效率。
3.2 不同Fe3+质量浓度对极北海带幼苗生长及氮磷需求的影响
Fe3+对极北海带幼苗的生长有影响,主要是因为其在细胞生长、对NO3--N、PO43--P的吸收利用以及参与光合呼吸过程等方面发挥重要作用[43-44]。适宜质量浓度的Fe3+能够促进极北海带的生长,缺少Fe3+则会影响营养盐的吸收以致抑制细胞生长[45]。研究发现,当Fe3+质量浓度为0.2~1 mg/L、NO3--N质量浓度为3 mg/L[ρ(N)∶ρ(P)=10∶1]时极北海带的相对生长速率较高。在相同Fe3+质量浓度下,培养3 d后NO3--N组的相对生长速率高于NH4+-N组。高质量浓度的Fe3+对极北海带的生长具有一定抑制作用。
Fe3+对极北海带吸收氮磷营养盐也有影响。当2种氮源同时存在时,大型海藻可能更倾向于对NH4+-N的吸收利用[46-47]。刘静雯等[13]认为,海藻对NH4+-N的吸收率远大于对 NO3--N的吸收率。海藻对 NO3--N的吸收一般是主动运输,对NH4+-N的吸收则以被动扩散为主。大型海藻在NO3--N下比在NH4+-N下生长需要更多的铁,这可能与大型海藻对NH4+-N和NO3--N吸收的运输方式有关[48]。本试验表明,适宜质量浓度的Fe3+能够显著促进极北海带对NO3--N、PO43--P的吸收,其作用时间主要是10 h之内,对不同质量浓度Fe3+的吸收也主要集中在10 h之内,10 h之后培养液中的Fe3+含量趋于平衡,并且和对照组无显著差异。
4 结 论
无外源铁离子时,极北海带幼苗在NO3--N质量浓度2~8 mg/L[ρ(N)∶ρ(P)=10∶1]时的相对生长速率较大;在氮、磷、铁营养盐试验中,添加适宜质量浓度的Fe3+,在10 h内对极北海带吸收NO3--N、PO43--P有显著的促进作用,添加1 mg/L的Fe3+对极北海带吸收NH4+-N无显著影响,NO3--N为3 mg/L、Fe3+为0.2~1 mg/L时较有利于极北海带幼苗的生长。
猜你喜欢
作文周刊·小学四年级版(2022年8期)2022-03-11
第一财经(2020年10期)2020-10-15
第一财经(2020年3期)2020-02-29
第一财经(2019年11期)2019-12-02
第一财经(2019年7期)2019-07-25
食品与生活(2017年12期)2018-01-09
河北渔业(2017年4期)2017-05-26
绿色科技(2016年24期)2017-03-30
河北渔业(2017年1期)2017-02-21
世界热带农业信息(2014年6期)2014-09-12
