
氯虫苯甲酰胺对亚洲玉米螟解毒酶活性及生长发育的影响
2021-01-26 03:15支昊宇丁新华陈萍叶茂林闫阳阳姜卫华
南京农业大学学报 2021年1期
支昊宇,丁新华,陈萍,叶茂林,闫阳阳,姜卫华*
(1.南京农业大学植物保护学院,江苏 南京 210095;2.新疆农业科学院植物保护研究所/农业农村部西北荒漠绿洲作物有害生物综合治理重点实验室,新疆 乌鲁木齐830091)
亚洲玉米螟(OstriniafurnacalisGuenée)是我国玉米生产的主要害虫,可造成玉米产量损失10%~30%,大发生时可达 50%以上,严重威胁玉米的品质及产量。2003年以来,亚洲玉米螟(以下简称玉米螟)在全国的发生呈持续上升趋势[1]。化学药剂的使用是防治该虫有效的手段之一,但杀虫剂的大量频繁施用,玉米螟的抗药性也会成为不可避免的问题[2-3]。
氯虫苯甲酰胺(chlorantraniliprole)是新型邻氨基苯甲酰胺类杀虫剂,它作用于鱼尼丁受体(Ryanodine receptor,RyR),具有高效、低毒、对其他类型杀虫剂无交互抗性等特点,引起市场广泛关注[4]。随着氯虫苯甲酰胺的推广使用,一些重要农业害虫,如甜菜夜蛾(Spodopteraexigua)、小菜蛾(Plutellaxylostella)、二化螟(Chilosuppressalis)对氯虫苯甲酰胺都已产生中等至高水平抗性[5-7]。调查发现该药剂对玉米螟也具有良好的室内活性及田间控制效果[8-9],但伴随着使用频率的增加,玉米螟对氯虫苯甲酰胺的敏感性势必会逐渐降低,目前尚无玉米螟对氯虫苯甲酰胺产生抗性的相关报道。
害虫对双酰胺类杀虫药剂的抗性机制,主要涉及细胞色素P450多功能氧化酶系(cytochrome P450 enzymes,P450)、谷胱甘肽S-转移酶(GlutathioneS-transferase,GST)及酯酶(Esterase,EST)3种解毒酶活性增强导致的代谢抗性[10],以及鱼尼丁受体结构突变和表达量的改变所引起的靶标抗性[11-12]。抗药性的产生一方面由于杀虫剂过量、不合理的使用,另一方面也和农药产生的亚致死作用有关。由于生物和非生物因素的影响,导致田间用药时部分害虫没有接触到足以致死的农药量,这种亚致死剂量不仅影响害虫的行为、生长发育和繁殖,还能对解毒酶活性及靶标受体表达量有诱导作用,从而对害虫抗药性的发展产生影响。有关氯虫苯甲酰胺对害虫的亚致死效应有不少研究报道。孙丽娜等[13]发现,低剂量的氯虫苯甲酰胺连续和间断处理均可使小菜蛾RyR基因表达量增加。氯虫苯甲酰胺亚致死剂量处理可引起二化螟雌成虫体内卵黄蛋白CsVg表达量下降[14]。亚致死剂量氯虫苯甲酰胺连续筛选小菜蛾5 代,其体内羧酸酯酶(Carboxylesterase,CarE)和P450比活力显著增强[15]。Plata-Rueda等[16]发现,氯虫苯甲酰胺LD50处理不仅降低咖啡果小蠹的存活率,还可降低其呼吸率,引起生物适合度成本的提高,同时也削弱肌肉活动导致的瘫痪。对天敌的处理发现,氯虫苯甲酰胺对异色瓢虫、七星瓢虫的生长发育有明显抑制效应[17-18]。由此可见,亚致死效应研究也是综合、全面评价杀虫剂防治效果中重要的一面。最近Song等[19]研究发现,氯虫苯甲酰胺对玉米螟的生长发育有抑制作用,但目前尚未有关于氯虫苯甲酰胺胁迫对玉米螟解毒酶活性影响的报道。
新疆是全国玉米主产区之一,种植总面积位于新疆粮食作物第2位[9]。玉米螟近年来在新疆主产区发生不断加剧[1,9],氯虫苯甲酰胺作为防治的主要化学药剂,当地玉米螟对其敏感性水平变化尚不明确。本研究对新疆不同地区玉米螟进行抗性水平及3种主要代谢酶活性的测定,发现新疆玉米螟种群对氯虫苯甲酰胺出现敏感性降低的现象;进一步采用低剂量氯虫苯甲酰胺处理室内玉米螟3龄幼虫,分析其生长发育以及体内解毒酶活性的变化,研究结果将为预防玉米螟对氯虫苯甲酰胺的抗性发展及该虫的有效防治提供理论依据。
1 材料与方法
1.1 供试虫源
2018年3—4月于新疆乌鲁木齐、伊宁及喀什地区疏勒和泽普采集玉米螟越冬老熟幼虫,转至室内利用人工饲料饲养,3龄幼虫作为抗性测定试虫。室内品系(S):疏勒种群不接触药剂人工饲料饲养7代。筛选品系(CAP-Sel):用氯虫苯甲酰胺LD50以点滴法处理室内品系3龄幼虫,连续筛选9代。养虫室温度(26±1)℃、相对湿度60%、光/暗时间为16 h/8 h。
1.2 供试药剂
96%氯虫苯甲酰胺原药(chlorantraniliprole)为南京红太阳股份有限公司产品。
1.3 毒力测定方法
利用点滴法测定玉米螟对药剂的敏感性。选取生长一致的玉米螟 3 龄幼虫,放入培养皿(直径9 cm)中。用丙酮将各原药按不同浓度梯度稀释,用Hamillton点滴器吸取1.1 μL药液,滴于玉米螟3龄幼虫前胸背板处,点滴丙酮作为对照。处理后以人工饲料饲喂试虫,并置于(26±1)℃、相对湿度 75%、光/暗时间为16 h/8 h的恒温培养箱中饲养。每处理重复3次,每重复10头幼虫。氯虫苯甲酰胺处理72 h后检查玉米螟的死亡情况。
1.4 酶活性测定
1.4.1 酶液制备氯虫苯甲酰胺LD20和LD50分别处理室内品系3龄幼虫72 h的存活试虫、CAP-Sel品系每世代及不同田间种群的3龄幼虫,用液氮速冻后放入-80 ℃保存备用。对照为丙酮处理或室内敏感品系,每处理10个重复。取试虫单头于匀浆器内,分别加入 1.2 mL 0.1 mol·L-1磷酸缓冲液(pH7.6,含 1 mmol·L-1EDTA,1 mmol·L-1DTT,1 mmol·L-1PTU,1 mmol·L-1PMSF)匀浆,用于细胞色素P450多功能氧化酶活性测定;加入 1.2 mL 0.1 mol·L-1磷酸缓冲液(pH7.6)匀浆用于谷胱甘肽S-转移酶活性测定;加入 1.2 mL 0.02 mol·L-1磷酸缓冲液(pH7.0)匀浆用于酯酶活性测定。匀浆液于 4 ℃、 13 000g离心 15 min 后,将上清液转移到新的 1.5 mL 离心管,重复离心1次后,取上清液作为酶液。
1.4.2 P450活性测定根据Rose等[20]的方法测定。在96孔酶标板中依次加入100 μL 2 mmol·L-1对硝基苯甲醚(P-NA)和90 μL酶液,在30 ℃温育3 min后,加入10 μL 9.6 mmol·L-1NADPH 开始反应。在SPECTRA max®340-PC型酶标仪(美国Molecular Devices公司生产)405 nm处每隔20 s记录15 min内的吸光值变化,酶促反应温度为30 ℃。P450比活力用ΔmD·min-1·mg-1表示。
1.4.3 EST活性测定根据Yang等[21]的方法测定。在96孔板中依次加入10 μL酶液、205 μL固蓝RR盐和α-NA混合液。用酶标仪在450 nm处每隔30 s记录10 min内的吸光值变化,酶促反应温度为27 ℃。EST比活力以ΔmD·min-1·mg-1表示。
1.4.4 GST活性测定根据Yang等[21]的方法测定GST活性。当底物为CDNB或DCNB时,分别在96孔酶标板中加入10 μL或25 μL酶液、1.2 mmol·L-1CDNB或DCNB 100 μL、6 mmol·L-1还原型谷胱甘肽100 μL。用酶标仪在波长340 nm处每隔10 s记录20 min内的吸光值变化,酶促反应温度为30 ℃。GST比活力以 ΔmD·min-1·mg-1表示。
1.4.5 总蛋白测定参照Bradford[22]考马斯亮蓝G-250法测定蛋白含量。
1.5 氯虫苯甲酰胺短期及长期处理对玉米螟生长发育的影响
利用氯虫苯甲酰胺亚致死剂量LD20点滴法处理室内品系3龄幼虫,72 h后将存活幼虫100头转入无毒人工饲料上继续饲养,每处理共设4次重复,以丙酮为对照组。另外,用氯虫苯甲酰胺LD50连续处理室内品系3龄幼虫9代,F10代建立生命表,每处理100头初孵幼虫,4个重复。每间隔24 h观察记载不同处理组幼虫的龄期,化蛹后分别记录化蛹数、蛹重、雌雄比及成虫羽化数。在各处理组中随机选取同日羽化的雌、雄玉米螟成虫各1头配对,每对作为1次重复,每处理至少60个重复。将配对成虫用玻璃试管移入塑料框(38 cm×26 cm×21.5 cm)内,顶部用黑色棉布封口,框底放置含10%蜂蜜水溶液的脱脂棉,以此作为成虫的补充营养。框内壁附着打印纸,供成虫产卵,每隔3 d更换新的产卵用纸,直至雌蛾死亡,收集并记录产于打印纸上的虫卵量,计算单雌产卵量。将卵块放入幼虫饲养盒中,放入人工饲料,待幼虫孵化记录幼虫孵化数,计算卵孵化率(幼虫孵化数/总卵量)。
1.6 数据分析
根据机率值分析法用Excel计算LD50值及95%置信限。抗性倍数(RR)=田间种群LD50/参照种群LD50,抗性级别划分标准:RR低于3倍为敏感;3.1~5.0倍为敏感性下降;5.1~10.0倍为低水平抗性;10.1~40.0倍为中等水平抗性;40.1~160.0倍为高水平抗性;高于160.0倍为极高水平抗性[23]。利用SPSS 18.0软件(Turkey’s检验)分析酶活性及生长发育数据的差异显著性(P<0.05)。
净增殖率(R0):指个体的总后代数,R0=∑lxmx。式中:x为特定年龄;lx为特定年龄x的存活率;mx为x时间的雌虫平均产卵数量。T为世代平均历期,T=∑lxmxx/R0。内禀增长率(r):指种群在理想状态下的最大种群增长率,r=lnR0/T。
2 结果与分析
2.1 新疆玉米螟种群对氯虫苯甲酰胺的敏感性
由表1可以看出:乌鲁木齐种群的敏感性最高,LD50为0.002 5 μg·头-1,以该种群作为相对敏感种群,伊宁种群也表现敏感(RR为1.32倍),而疏勒和泽普种群则敏感性降低,RR分别为4.00和4.20倍。

表1 新疆不同地区玉米螟对氯虫苯甲酰胺的敏感性Table 1 Sensitivity to chlorantraniliprole of different populations of Ostrinia furnacalis in Xinjiang using topical application(2018)
2.2 氯虫苯甲酰胺对玉米螟解毒酶活性的影响
2.2.1 新疆不同玉米螟种群解毒酶活性的差异从表2可以看出:对氯虫苯甲酰胺敏感性不同的玉米螟种群之间P450和GST(DCNB为底物)活性差异显著。其中,伊宁种群的P450酶活性最高,乌鲁木齐种群次之,泽普和疏勒种群的酶活性最低;对于GST(DCNB为底物),疏勒种群的酶活性最高,伊宁和泽普种群的酶活性最低。而GST(CDNB为底物)和酯酶的酶活性均为泽普最高,但与其他种群之间没有显著差异。
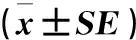
表2 新疆地区玉米螟种群解毒酶活性比较Table 2 Comparison of detoxification enzyme activities of O.furnacalis in Xinjiang
2.2.2 氯虫苯甲酰胺短期及长期处理对玉米螟解毒酶活性的影响测定玉米螟室内品系对氯虫苯甲酰胺的敏感性,得出LD20和LD50分别为0.30和0.71 ng·头-1。由表3可见:氯虫苯甲酰胺LD20和LD50处理玉米螟72 h,3种解毒酶活性随处理剂量的升高而增加。LD20和LD50处理组的GST(DCNB为底物)活性显著高于对照,分别增加46%和86%(活性比分别为1.46和1.86),以CDNB为底物的GST活性高于对照但差异不显著。LD50处理组的EST酶活性显著高于LD20,但与对照均无显著差异。
当使用LD50剂量连续筛选玉米螟时,发现随筛选代数的增加,3种解毒酶活性都呈增加的趋势(图1),但第2、5和8代(F2、F5、F8)的EST活性与敏感品系相比都无显著差异。P450酶在F5和F8代的活性是敏感品系的4.27和5.09倍(P<0.05);F5和F8代GST(底物CNDB)及F8代的GST(底物DCNB)活性分别是敏感品系的1.27、1.33和1.36倍(P<0.05)。可以看出:氯虫苯甲酰胺长期与短期处理对玉米螟解毒酶活性影响的趋势接近。
2.3 氯虫苯甲酰胺对玉米螟生长繁殖的影响
从表4可以看出:与对照相比,经氯虫苯甲酰胺亚致死剂量LD20处理72 h,玉米螟蛹重和单雌产卵量显著降低,蛹重降低9.2%,单雌产卵量减少约30%;此外,幼虫生长期和蛹期都延长,成虫羽化率、成虫雌雄比、卵孵化率降低,但与对照相比差异不显著。最终处理组的内禀增长率低于对照,但无明显差异,表明氯虫苯甲酰胺短期处理对玉米螟的生物适合度有一定影响但不显著。
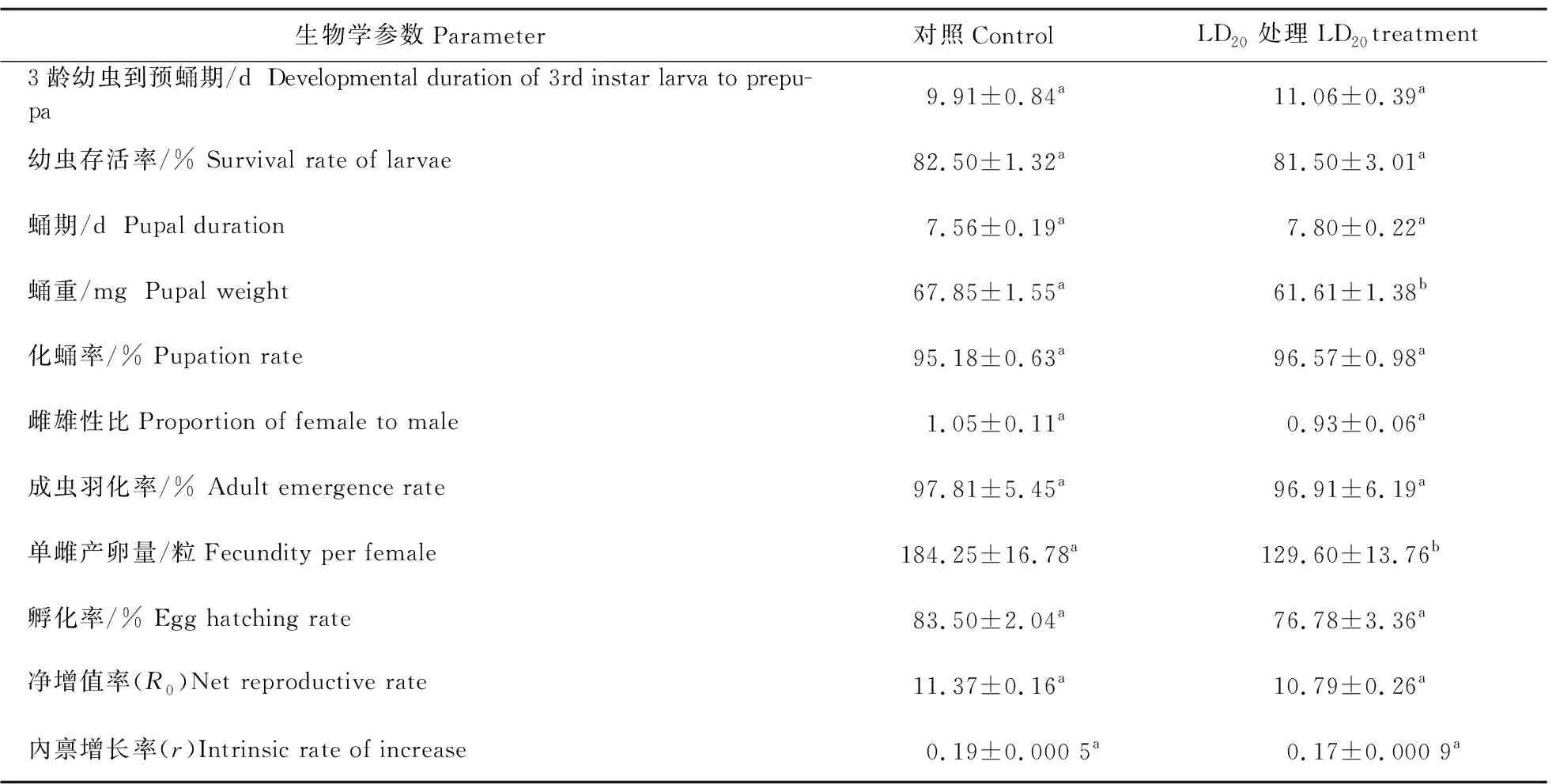
表4 氯虫苯甲酰胺亚致死剂量(LD20)处理72 h对玉米螟存活和生长发育的影响Table 4 Effects of chlorantraniliprole sublethal dose LD20 treated for 72 h on development of O.furnacalis
从表5可以看出:与敏感品系相比,玉米螟筛选品系F10代1龄和3龄幼虫期显著延长,蛹重、羽化率显著增加6.58%和4.27%;单雌产卵量及孵化率比敏感品系显著降低14.83%和5.84%。筛选品系的内禀增长率显著低于敏感品系,表明氯虫苯甲酰胺长期处理对玉米螟的生物适合度有显著影响。
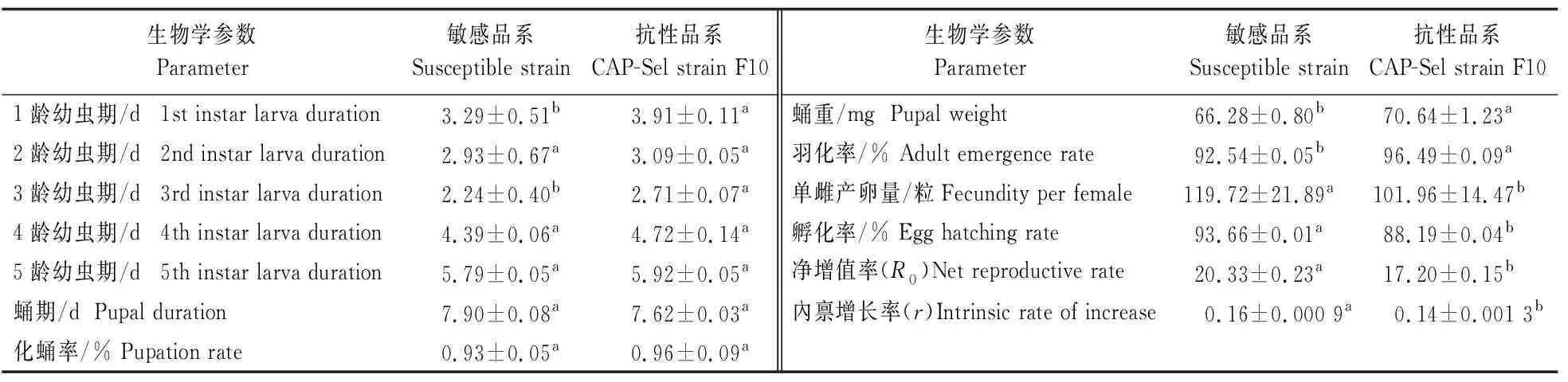
表5 氯虫苯甲酰胺(LD50)连续处理对玉米螟存活和生长发育的影响Table 5 Life tables of the susceptible strain and tenth generation of O.furnacalis selected continuously by LD50 dose of chlorantraniliprole
3 讨论
新疆地区近些年玉米螟的发生呈爆发态势,除了以赤眼蜂为主的生物防治以外,化学药剂的使用对于有效、快速地控制玉米螟种群数量尤为重要。氯虫苯甲酰胺是近几年来才开始用于玉米螟的防治,本文抗性监测结果发现相对于乌鲁木齐种群,疏勒和泽普种群对氯虫苯甲酰胺呈现敏感性降低,可见玉米螟对该药剂抗性发展的风险较高,有必要加强当地玉米螟的抗性监测,为合理用药和有效防治提供科学指导。
害虫抗药性机制的阐明是抗性治理的重要前提。其中解毒酶活性的增强是害虫抗药性产生的重要机制之一。亚致死效应涉及害虫的行为学、生物学、生理学及抗药性发展等很多方面,有关研究可为药剂的合理使用、避免或减少药剂的副作用提供一定的理论依据。本研究初步发现,疏勒和泽普种群对氯虫苯甲酰胺有发展抗性的潜在趋势,通过亚致死效应研究可初步明确玉米螟对氯虫苯甲酰胺抗性产生相关的解毒酶。
氯虫苯甲酰胺处理对不同害虫解毒酶的诱导作用不同。Mao等[24]发现,0.01 mg·L-1氯虫苯甲酰胺处理家蚕可使其P450和GST比活力显著升高。陈羿渠等[25]发现,甜菜夜蛾氯虫苯甲酰胺抗性品系的GST与P450活性显著高于敏感品系,用亚致死剂量氯虫苯甲酰胺处理抗性品系后其3 种解毒酶活性均有一定程度的上升,且P450 活性的增长倍数最高。与之结果相似的是,本研究发现玉米螟筛选品系F5和F8代的P450和GST活性都显著升高,其中 P450活性增长最为显著;另外氯虫苯甲酰胺LD20、LD50短期处理对玉米螟解毒酶活性都有诱导作用,但仅GST活性显著升高;酯酶在玉米螟对氯虫苯甲酰胺的代谢中不起主要作用。Li等[26]研究表明,不同褐飞虱田间种群的GST活性与其对氯虫苯甲酰胺的LD50值呈正相关。本研究也发现,对氯虫苯甲酰胺敏感性降低的疏勒和泽普玉米螟的GST活性也相对较高。综合以上结果作者认为,GST和P450可能与玉米螟对氯虫苯甲酰胺的敏感性下降有关。但也有与本研究结果不同的报道,如亚致死剂量氯虫苯甲酰胺对棉铃虫的GST活性有抑制作用[27];而东亚飞蝗经氯虫苯甲酰胺亚致死剂量处理7 d可导致其GST活性显著升高,但P450活性显著降低[28]。究其原因可能是由于受药昆虫的种类和生理状况、施用杀虫剂及其浓度、处理方法等差异所引起的。解毒酶活性增强是害虫产生抗性的主要机制之一。研究发现在杀虫剂胁迫下昆虫解毒代谢酶可通过基因表达量上调和突变导致解毒能力加强,从而使昆虫产生抗药性[10,29-30]。本研究中玉米螟经氯虫苯甲酰胺处理引起P450和GST活性增加的分子机制有待进一步研究探明。
杀虫剂亚致死剂量不仅对昆虫的解毒酶具有诱导或抑制作用,也可对害虫种群的生物学特性产生影响。欧善生等[27]利用氯虫苯甲酰胺亚致死剂量(LC10、LC25和LC50)处理可引起棉铃虫幼虫和蛹重显著降低,幼虫期显著延长,而成虫期显著缩短。Song等[19]用氯虫苯甲酰胺LC10(0.038 mg ·L-1)和LC40(0. 098 mg ·L-1)处理玉米螟3 龄幼虫120 h后,发现幼虫和蛹的发育历期显著延长,产卵量也显著降低。本研究发现,氯虫苯甲酰胺LD20短期处理可造成玉米螟的蛹重和产卵量显著降低,3龄幼虫至预蛹的生长期延长但与对照无显著差异,这与Song等[19]的结果不尽相同但具有相似的趋势。陈羿渠等[25]利用亚致死剂量氯虫苯甲酰胺连续筛选甜菜夜蛾6代后,发现抗性品系成虫的产卵前期和总产卵前期延长,而产卵量减少,其内禀增长率、周限增长率和净增殖率均显著小于敏感品系。与之类似的是,本研究中氯虫苯甲酰胺连续筛选玉米螟9代可导致F10代1龄和3龄幼虫期显著延长,蛹重、羽化率显著增加,但产卵量、孵化率显著降低,最终内禀增长率显著低于对照,可以看出本研究中无论是短期或长期处理的结果都表明氯虫苯甲酰胺对玉米螟的生长发育具有抑制作用。
本文对新疆玉米螟抗性监测结果显示:泽普和疏勒种群对氯虫苯甲酰胺敏感性降低,且分别具有最高的GST活性(底物分别为CDNB和 DCNB);氯虫苯甲酰胺短期和长期处理不仅抑制玉米螟的生长发育,也可诱导GST和P450活性增加,有利于玉米螟对氯虫苯甲酰胺的抗性发展。因此,新疆玉米螟具有对氯虫苯甲酰胺产生抗性的风险,其中GST和P450活性的增加可能是敏感性降低和抗性形成的机制之一,这有待于进一步的研究证实。
猜你喜欢
农产品加工(2021年21期)2021-12-31
农药科学与管理(2021年8期)2021-10-22
右江民族医学院学报(2021年4期)2021-09-18
湖北植保(2021年4期)2021-08-23
现代农药(2021年2期)2021-05-07
今日农业(2020年17期)2020-12-15
今日农业(2020年14期)2020-12-14
现代农村科技(2020年9期)2020-09-08
中国果菜(2020年1期)2020-02-23
云南农业科技(2019年2期)2019-04-04
