
“十三五”我国甘蓝遗传育种研究进展
2021-01-20 14:40
中国蔬菜 2021年1期
(中国农业科学院蔬菜花卉研究所,农业农村部园艺作物生物学与种质创制重点实验室,北京 100081)
结球甘蓝(Brassica oleraceaL.var.capitataL.)简称甘蓝,是一种重要的十字花科蔬菜作物。甘蓝具有较强的适应性和抗逆性,其叶球中含有丰富的营养物质,在世界各地普遍栽培。据统计,我国甘蓝的种植面积约90 万hm2(杨丽梅 等,2020),居世界第一位,在全国蔬菜周年供应及出口贸易中占有十分重要的地位。“十三五”以来,在国家重点研发计划、大宗蔬菜产业技术体系、国家自然科学基金及一些省部级科研项目的支持下,我国甘蓝遗传育种取得了重要进展:获得省部级奖励3 项,授权发明专利30 余项,发表文章90 余篇;通过远缘杂交、小孢子培养及基因编辑等技术创制出优良育种材料逾100 份,对重要农艺性状基因进行了定位或克隆,并开发了可用于辅助选择的分子标记,培育出一批优质、多抗、适应性强的甘蓝新品种,显著提升了我国甘蓝遗传育种的水平。
1 种质资源的搜集、鉴定与创新
1.1 种质资源的搜集与鉴定
优良种质资源是甘蓝新品种培育的基础。据中国农业科学院蔬菜花卉研究所、北京市农林科学院蔬菜研究中心、江苏省农业科学院蔬菜研究所等国内7 家科研单位统计,近5 年来国内主要甘蓝育种单位共搜集、引进国内外甘蓝种质资源逾500份,为优良新品种培育奠定了材料基础。王神云等(2016)对搜集的88 份甘蓝种质材料进行苗期和成株期根肿病抗性鉴定,获得抗ECD17/31/13 生理小种的材料5 份。马丹丹等(2016)通过人工注射法结合分子标记辅助选择对94 个甘蓝类蔬菜品种进行根肿病抗性鉴定,从中筛选出高抗4 号生理小种的甘蓝品种11 个。孙超等(2016)对22 份甘蓝材料的根肿病抗性进行鉴定,筛选出3 份对河南新野菌种免疫的材料和1 份对陕西太白菌种表现高抗的材料。Ning 等(2018)对102 份甘蓝种质资源进行根肿病抗性鉴定,筛选出高抗4 号生理小种的材料4 份。张小丽等(2019)对306 份甘蓝类蔬菜品种(系)进行根肿病抗性鉴定发现,甘蓝类蔬菜中抗根肿病资源严重匮乏,仅从青花菜中筛选出1份高抗材料。孔枞枞等(2018)通过人工接种鉴定从99 份国内外甘蓝种质资源中筛选获得54 份高抗枯萎病材料和26 份抗黑腐病材料,其中包括5 份对枯萎病和黑腐病表现双抗的材料。李岸(2019)对64 份甘蓝种质资源进行苗期枯萎病抗性鉴定,筛选出抗病材料41 份,其中高抗材料12 份。
对上述抗性材料的地理来源进行分析发现,日本、韩国等国家的抗病资源较我国丰富。广泛搜集和引进优良种质资源仍将是未来一段时间内我国甘蓝遗传育种尤其是抗病育种的重要任务。
1.2 优异种质创新
1.2.1 通过小孢子培养创制优良DH 系 甘蓝常规育种中,通常需要6~7 代的连续自交才能育成纯合稳定的亲本自交系,育种周期较长,而通过游离小孢子培养技术可在2 a 内获得纯合的双单倍体DH 系,大大缩短育种周期,提高育种效率。苏贺楠等(2018)通过对小孢子培养条件、培养基成分和培养方式进行改进和优化,显著提高了甘蓝小孢子成胚率,并成功诱导20 个基因型产生了胚状体,其中杂交种中甘628 和自交系01-88 出胚率最高,分别达到19.8、47.5 胚·蕾-1。通过田间表型鉴定,初步筛选出一批农艺性状优良的DH 系如2189 DH13、SH11DH5、SH10DH22、根372DH13、根373DH7、2006DH1 等,已开始用于试配杂交组合(苏贺楠,2020)。
1.2.2 通过远缘杂交创制优异育种材料 为实现甘蓝类蔬菜Ogura 胞质不育资源的利用,于海龙(2018)利用远缘杂交结合胚挽救技术,将甘蓝型油菜中的Ogura 胞质不育恢复基因Rfo成功导入芥蓝中,并通过多代回交结合标记筛选,成功获得育性恢复稳定、染色体数目正常(2n=18)的芥蓝Ogura CMS 恢复系。以该恢复系作为桥梁,对抗根肿病甘蓝Ogura 胞质不育材料进行了育性恢复,获得了含有根肿病抗性位点CRb的育种材料,为抗根肿病品种培育提供了一条新途径(Yu et al.,2016;Ren et al.,2020)。此外,宁宇(2019)利用抗根肿病大白菜与甘蓝进行远缘杂交和回交,获得4 株含有根肿病抗性位点CRa和CRb的BC1抗病材料。彭丽莎等(2016)利用抗根肿病大白菜品种CR-春美、CR-英雄与甘蓝进行远缘杂交,结合胚挽救技术获得对根肿病4 号生理小种和11 号生理小种表现抗病的种间新种质,丰富了甘蓝抗根肿病的育种材料。
1.2.3 通过基因编辑技术创制优良育种材料 以CRISPR/Cas9 为代表的基因编辑技术是植物基因功能研究及作物改良的有力工具,目前该技术已在多种作物精准育种方面发挥了重要作用。Ma 等(2019)在结球甘蓝中建立了高效的基因编辑技术体系,结合tRNA 多靶点表达载体,完成甘蓝八氢番茄红素脱氢酶基因BoPDS、雄性不育相关基因MS1、自交不亲和基因BoSRK的定点突变,不同基因突变效率在30%以上,自交结果显示,突变位点可以遗传到子一代。Cao 等(2021)利用CRISPR/Cas9 技术对甘蓝蜡质合成相关基因BoCER1进行敲除,成功获得蜡质缺失亮绿甘蓝材料。CRISPR/Cas9 基因编辑技术在甘蓝中的成功应用,为后续甘蓝多基因聚合育种奠定了坚实的技术基础。通过基因编辑获得的甘蓝自交亲和系、雄性不育系、蜡质缺失突变体等对后续甘蓝遗传育种研究具有重要的应用价值。
2 甘蓝重要基因/QTL 研究
“十三五”以来,为了提高育种效率,加快育种进程,各单位先后开展了与甘蓝抗病、抗逆、雄性不育及无蜡粉亮绿等性状相关的基因/QTL 的定位与克隆研究,并开发了紧密连锁的分子标记,为甘蓝分子聚合育种奠定了基础。
2.1 甘蓝抗病基因/QTL 研究
2.1.1 枯萎病抗性基因的克隆与功能分析 甘蓝枯萎病是一种由尖孢镰刀菌粘团专化型致病菌引起的土传病害,一旦发生将在土壤中存活10 a 以上且无法根除。Liu 等(2019)收集并鉴定了国内20 份枯萎病致病菌分离物,并与国际标准菌株进行比较,发现国内枯萎病致病菌几乎全部为1 号生理小种,且致病力均高于国际标准菌株。目前已知甘蓝对枯萎病1 号小种的抗性受显性单基因控制(刘星,2017;Liu et al.,2017a)。
在枯萎病抗性基因克隆方面,刘星(2017)构建了甘蓝抗枯萎病基因FOC1的过表达载体,并通过农杆菌介导的遗传转化将其导入感病甘蓝自交系材料XW 中,获得了FOC1基因高表达的阳性植株,抗性鉴定结果表明,转基因阳性植株的抗病能力显著提升,人工接种后未出现发病症状。此外,基于全基因组重测序数据,Liu 等(2017a)开发了与甘蓝枯萎病抗性紧密连锁的分子标记Frg13,物理距离为75 kb,利用标记辅助育种准确率达95%以上,提高了抗病材料的筛选效率;与此同时,以抗病DH 系材料D134 作为抗性供体亲本,结合标记筛选通过回交的方式将抗枯萎病基因成功转移到优质的骨干亲本01-20 中,为甘蓝抗枯萎病优质品种的培育奠定了基础。
2.1.2 黑腐病抗性QTL 定位 甘蓝黑腐病是由野油菜黄单胞菌野油菜致病变种侵染引起的细菌性病害,近年来,随着甘蓝栽培面积的增加,其危害程度也日益严重。目前多数研究表明,甘蓝对黑腐病的抗性受多个基因控制。孔枞枞(2019)通过遗传分析发现,甘蓝对黑腐病1 号生理小种的抗性由1对加性主基因+加性-显性多基因控制;利用抗感双亲构建的F2群体结合QTL-seq 技术,将抗1 号生理小种的QTL 定位在3 号染色体25.44~31.72 Mb 区间内,两侧标记分别为I254 和I317,该区域最高可解释38.4%的表型变异。此外,刘泽慈(2019)利用重组自交系与染色体片段替换系将甘蓝抗黑腐病3 号生理小种的主效QTL 位点qBR-7-3定位在7 号染色体2.91 Mb 的区间内,该区域可解释16.72%的表型变异,推断该区间内NBSLRR 类型基因Bo7g111290可能为抗病基因。上述开发的标记以及确定的抗性位点为分子标记辅助抗病育种及黑腐病抗性基因的克隆奠定了基础。
2.1.3 根肿病抗性QTL 定位 根肿病是一种由芸薹根肿菌侵染引起的危害极大的土传病害,俗称“根癌”,目前已成为全球十字花科蔬菜产区的主流病害之一。随着十字花科蔬菜栽培面积的扩大、商品种子和蔬菜产品的相互调运及长期连作,我国根肿病的发生面积也在逐年扩大。
研究表明,甘蓝对根肿病的抗性是由多基因控制的数量遗传性状。张小丽(2017)将野生甘蓝B2013 中抗根肿病4 号生理小种的主效位点定位在C09 染色体32.01~40.01 Mb 的区间内,并推测区间内的Bol044005为可能的抗性候选基因。Peng 等(2018)利用SNP 基因芯片在甘蓝抗病材料中检测到23 个与根肿病抗性连锁的QTL 位点,可解释的表型变异介于6.1%~17.8%之间。宁宇(2019)结合QTL-seq 技术,将甘蓝抗病材料2358 中抗4号生理小种的1 个QTL 位点qBoCR8.1定位在8 号染色体500 kb 的区间内,该区间可解释16.4%的表型变异。这些抗根肿病QTL 位点的确定为抗源材料的筛选和抗病基因克隆奠定了基础。
2.2 甘蓝抗逆相关QTL 研究
2.2.1 耐抽薹QTL 定位 甘蓝属于绿体春化植物,如果在结球前遇到长时间的低温和长日照条件满足其春化要求,就会发生先期抽薹,严重影响其产量和品质。研究表明,甘蓝未熟抽薹的难易程度是受多个基因控制的数量遗传性状,同时受环境因素的影响(王五宏等,2020)。陈登辉等(2018)利用耐抽薹和易抽薹材料构建的重组自交系群体对甘蓝耐抽薹QTL 进行定位,在8 号染色体上检测到1 个QTL 位点qbt-2-2,贡献率为9.1%,获得连锁分子标记CB10139。此外,王五宏等(2020)采用SLAF-BSA 方法在甘蓝2 号染色体2.31~3.09 Mb 和33.57~34.40 Mb 区间内定位到2 个耐抽薹QTL 位点。以上耐抽薹位点及其连锁标记的获得可用于耐抽薹材料的辅助选择,提高育种效率。
2.2.2 耐裂球QTL 定位 在甘蓝栽培过程中,时常会出现叶球开裂的现象,严重影响外观和品质。研究表明,甘蓝耐裂球性状的遗传符合G-0 模型,即该性状的遗传受3对加性-上位性主基因+加性-上位性多基因控制(苏彦宾等,2019)。朱晓炜等(2018)结合QTL-seq 技术将甘蓝耐裂球QTL 定位在3 号染色体上46~51 Mb 的区间内,并将与拟南芥细胞壁合成相关基因At4g38770和At4g19120的同源基因Bol016058和Bol029881确定为候选基因。该研究可为甘蓝耐裂球分子设计育种提供参考。
2.3 甘蓝蜡质缺失基因的定位与克隆
普通甘蓝植株表面覆盖有一层蜡质,球叶颜色表现为绿色、灰绿色或深绿色,而甘蓝蜡质缺失突变体的球叶则呈现为亮绿色,具有良好的商品性。Liu 等(2017b)以甘蓝隐性蜡质缺失突变体LD10 及其野生型为材料,通过图位克隆将与拟南芥CER4同源的基因Bol013612确定为候选基因,突变体中该基因发生单碱基突变导致其mRNA 转录剪切异常,通过转基因试验最终证明LD10 的蜡质缺失性状是由Bol013612突变造成。Ji 等(2018)以甘蓝隐性蜡质缺失突变体g21-3 及野生型为材料,通过图位克隆技术获得g21-3 蜡质缺失候选基因BoCER1,突变体中该基因第4 个内含子中插入了1 个252 bp 的片段,该片段的插入严重抑制了该基因的表达,根据插入位点设计的分子标记与蜡质缺失性状100%连锁。Dong 等(2019)结合BSA 法对甘蓝显性蜡质缺失基因BoGL-3进行了精细定位,将BoGL-3定位于8 号染色体末端33.5 kb的区间内,开发了1 个与显性蜡质缺失性状紧密连锁的SSR 标记,将该区间内与拟南芥CER1同源的基因Bol018504确定为候选基因,该基因在显性蜡质缺失突变体中的表达受到严重抑制。这些蜡质缺失基因的克隆及标记的开发为无蜡粉亮绿甘蓝材料的筛选和创制奠定了基础。
2.4 甘蓝雄性不育基因的克隆
雄性不育是指植物雄性生殖器官不能产生正常有活力花粉的现象,雄性不育突变体在甘蓝杂种优势利用中发挥着重要作用,同时也是研究植物生殖发育调控的重要材料。Ji 等(2017)通过比较基因组学和转录组学分析,将BoCYP704B1确定为甘蓝83121A 隐性雄性不育的候选基因,测序发现83121A 突变体BoCYP704B1基因中插入了1 个反转录转座子,抑制了该基因表达,同时也改变了其转录剪切位点,根据突变位点设计的引物与不育性状100%连锁,可用于不育材料的辅助选择。Han等(2018)通过图位克隆技术将甘蓝隐性核雄性不育基因ms3定位在C01 号染色体187.5 kb 的区间内,并将BoTPD1确定为ms3的候选基因。研究发现,雄性不育突变体的BoTPD1基因中有一段182 bp 片段的插入,根据该变异位点设计的分子标记与雄性不育性状100%连锁,可用于雄性不育材料的辅助选择。此外,Han 等(2019)在显性不育基因Ms-cd1定位的基础上,基于重测序数据开发了1 个与显性不育性状紧密连锁的KASP 分子标记,可用于显性不育基因位点的快速鉴定。以上雄性不育基因的克隆和标记的开发为优良雄性不育材料的鉴定和创制提供了依据。
2.5 甘蓝小孢子胚胎发生能力QTL 定位
目前关于甘蓝游离小孢子培养体系的研究已取得较大进展,但是与小孢子胚胎发生相关遗传机制的研究较少。苏贺楠(2020)通过对高出胚甘蓝材料01-88 和难出胚甘蓝材料02-12 的杂交一代进行游离小孢子培养构建了包含109 个DH 系的群体,对该群体进行出胚率、表型鉴定及遗传分析,发现甘蓝小孢子的胚胎发生能力是由多个基因控制的数量性状。通过QTL-seq 分析发现甘蓝3 号染色体57.4~58.4 Mb 的区间内存在控制甘蓝胚胎发生能力的QTL 位点,进一步构建遗传图谱进行遗传连锁分析,将控制甘蓝出胚性状的主效QTL 定位在3号染色体54.1~57.7 Mb 的区间内,该区间LOD 值为2.5~3.6,最高可解释14.6%的表型变异。该研究为甘蓝胚胎发生相关基因的鉴定和克隆提供了理论依据。
3 甘蓝新品种选育
“十三五”期间,为满足甘蓝生产和多样化消费的需求,培育出一批生产上急需的抗枯萎病/黑腐病、耐抽薹、耐裂球、耐热/耐寒、品质优良的甘蓝新品种,部分完成登记和获得植物新品种权的品种见表1。例如中甘628、中甘828、中甘56、中甘1305、京甘4 号、京甘5 号、西园秋丰、西园冬秀、春秋婷美、苏甘65 等(曾爱松 等,2017;王神云 等,2018;张扬勇 等,2018,2020;吕红豪 等,2019)。这些新品种在产量、品质、抗病、抗逆等方面较原有品种有了显著提升,具有良好的推广应用前景。其中,中甘628、中甘828、京甘4 号等优良抗枯萎病新品种已大面积应用,累计在枯萎病危害严重地区推广逾6.67 万hm2(100 万亩),减少了农药用量及栽培成本。这些品种的推广基本满足了我国甘蓝周年栽培、周年供应和产品类型多样化的需求。
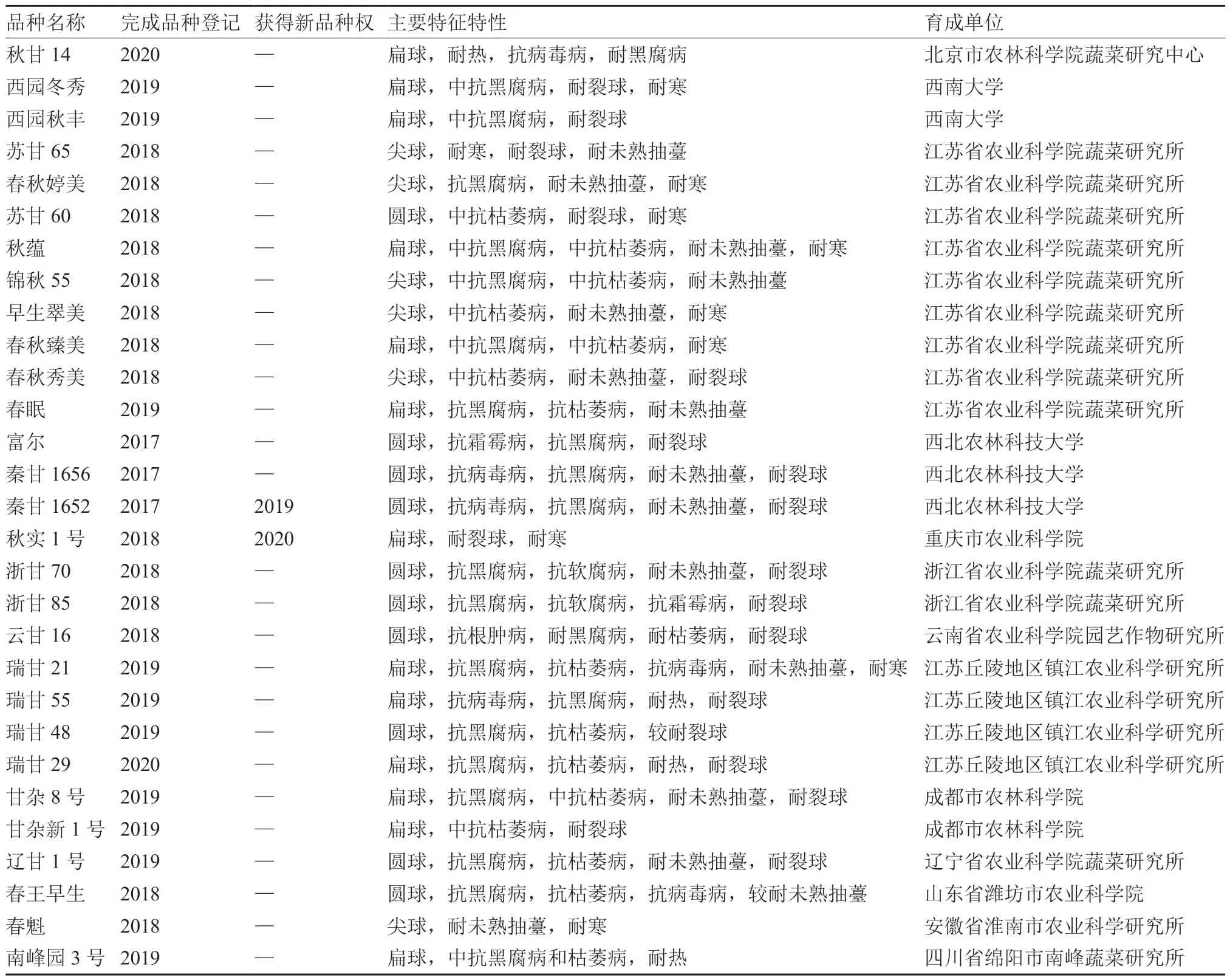
续表
4 问题与展望
4.1 进一步加强种质资源搜集工作和优异种质创新
种质资源是育种工作的基础,目前我国甘蓝种质资源还不够丰富,特别是抗黑腐病和根肿病等优异种质资源匮乏。今后应继续从国内外广泛搜集优良种质资源,并深入开展种质资源的鉴定和评价工作。此外,应用现代高新技术与常规育种技术相结合进行甘蓝种质创新,是加快新品种培育,提升甘蓝育种水平的基础性工作。
4.2 加强应用基础和育种技术研究,提升甘蓝育种水平
我国现阶段甘蓝育种中基础研究相对薄弱,特别是抗病育种基础研究有待进一步加强,例如根肿病、黑腐病等生理小种分化十分复杂,目前尚未创建精确的生理小种鉴别方法,且甘蓝根肿病、黑腐病抗性基因研究主要处于QTL 定位阶段。后续应进一步加大对基础研究的投入力度,在建立精确的根肿病、黑腐病生理小种鉴别体系的基础上,加快抗病基因的克隆。同时,要更加重视常规育种技术与生物技术的结合,进一步通过小孢子培养、远缘杂交和基因编辑等技术创制符合市场需求的优异育种材料,为今后高竞争力甘蓝新品种培育奠定坚实的材料基础。
4.3 根据生产和市场需求培育新品种
近年来,随着我国蔬菜产业的发展,甘蓝生产面积也迅速增加。随着市场需求、生产基地及茬口的变化,对甘蓝育种提出了更多新的需求。一方面,为实现甘蓝多个季节栽培和周年供应,要重视选育不同球形、不同熟性,适于春、夏、秋、冬多茬栽培的品种;另一方面,随着甘蓝规模化生产基地的建立,公司+农户的产销方式已经形成,即农户生产,公司收购,远距离运往市场销售,因此也要重视培育适于优势产区规模化生产基地的耐裂球、耐贮运的甘蓝品种。此外,还要有目的地了解国外市场需求,培育适应某些特定国家或地区需求的甘蓝品种,使我国培育的甘蓝品种逐渐打入国外市场。
猜你喜欢
热带农业科学(2022年4期)2022-11-23
现代园艺(2022年19期)2022-11-18
天津农业科学(2022年5期)2022-05-31
中国瓜菜(2022年3期)2022-05-05
作物学报(2021年2期)2021-12-24
农民文摘(2021年11期)2021-12-14
上海农业科技(2021年5期)2021-10-12
小天使·二年级语数英综合(2019年10期)2019-11-08
热带作物学报(2018年9期)2018-05-14
科学种养(2017年11期)2017-11-28
