
樟科楠属4种植物种实表型性状及其发芽特性研究
2021-01-16 13:07陆云峰徐沁怡李修鹏王建军
种子 2020年12期
陆云峰, 王 豪, 徐沁怡, 李修鹏, 王建军
(宁波市林特科技推广中心, 浙江 宁波 315012)
林木果实、种子作为生殖器官,相较于叶片等营养器官,其表型性状更具稳定性,是林木长期遗传和自然环境选择的综合体现,在遗传育种上具有重要意义[1-2]。林木遗传变异研究可通过表型测定、细胞学、生化和分子标记等方法,其中,表型测定具有简单、直观、易测等特点而被广泛应用[3-4]。种子发芽过程是实现种群更新和物种延续的重要阶段,是构建植物群落的关键环节[5]。同时,林木种实性状与发芽特性呈较高程度的相关性[6-7]。因此,开展种实表型性状及发芽特性的研究,是苗木培育和品种改良的关键。
楠属(Phoebe)是樟科(Lauraceae)中经济价值极高的一个属,全世界约有94种,我国有34种3变种,主要零散分布于长江流域及长江以南地区[8]。该属植物木材坚硬致密,纹理直,有光泽,耐腐且富楠木芳香,是家具、雕刻的高级用材;同时其树体高大通直,枝叶繁茂且常绿,也可作为优良园林绿化树种。其中,桢楠(Ph.zhennan)、闽楠(Ph.bournei)、浙江楠(Ph.chekiangensis)、紫楠(Ph.sheareri)等因其珍贵稀有而被称为“金丝楠木”。然而,由于气候环境变化,人类过度砍伐,自然更新能力弱等诸多原因,楠属植物天然资源越来越少,开展资源保护和良种选育工作迫在眉睫[9]。目前,针对楠属植物的研究主要集中在资源调查、遗传多样性分析、育苗技术、造林方法等方面[10-14],而对其果实、种子性状及其发芽特性的研究鲜见报道。鉴于此,本研究收集楠属4种植物(桢楠、闽楠、浙江楠、紫楠)的代表性种源的成熟果实16份,通过测定果实、种子的表型性状及播种试验,研究其不同种间、同一植物不同种源间果实、种子表型性状及其发芽特性,以了解其表型变异规律和实生繁育能力,为楠属植物的遗传改良和高效育种提供理论依据。
1 材料与方法
1.1 试验材料
在查阅文献记载和野外实地调查的基础上,于2018年11—12月在四川、重庆、湖北、湖南、江西、福建、浙江、江苏等省份开展楠属4种植物(桢楠、闽楠、浙江楠、紫楠)成熟果实的野外采集工作,共计16个采样点(表1)。采用全球卫星定位系统(Global Positioning System,GPS)记录各采样点的经纬度及海拔,并查阅获得其年均温、年降水量、年日照时长等气候因子。选择生长健壮、结实良好的单株作为采样母树,并尽量拉大两采样母树之间的距离,避免其亲缘关系。在各母树的树冠中上部东、南、西、北4个方向进行均匀采样,并将各采样点的样品混匀保湿保存,带回实验室进行测定。

表1 楠属4种植物16个群体的地理气候概况
1.2 试验方法
1.2.1果实、种子表型性状测定
每个参试种源随机选取300粒健康饱满果实,分为3个重复,每个重复100粒。用电子游标卡尺测量果实长与宽(测量精度为0.01 mm),计算果实长宽比(形态指标);用电子天平称100粒果实质量(称量精度为0.001 g),记为M果实;剥取果皮,使与种子分离,果皮放入40 ℃烘箱烘干至恒重后称重,记为M种皮。同理,测量种子长与宽,计算种子长宽比;称量去皮后的100粒种子质量,记为M种子。
种皮含水量(%)=[1-M种皮/(M果实-M种子)]×100%;
最后,先向100 mL量筒倒入适量自来水,记下读数1,再将测定完百粒重后的100粒种子加入量筒,记下读数2,则单粒种子的平均体积为:(读数2-读数1)/100。
1.2.2播种育苗
种子去皮后,采用混沙湿藏越冬催芽。播种育苗试验地位于浙江省宁波市林业局林特种苗繁育中心东吴苗圃(121°42′E,29°49′N),于2019年2月上旬开始播种,将泥炭与珍珠岩按1∶1的比例混合作为基质,每立方米基质中施入1.5~3.5 kg缓释肥。所施缓释肥为美国爱贝施(APEX)长效控释肥,全氮含量为180 g·kg-1,有效磷和有效钾含量分别为60 g·kg-1、120 g·kg-1[15]。各个种源随机选取300粒种子进行播种,分为3个重复,每个重复100粒。将上述基质装入无纺布容器袋,种子直接播于其中,育苗过程需要保持基质湿润。种子开始萌发后,持续观察其生长情况,每天定时记录当天萌发出土的种子数。当连续5 d无种子萌发出土,结束发芽试验[16-17]。
试验结束后分别计算种子的发芽率、发芽势和发芽指数,计算公式如下:
发芽率(%)=(正常发芽种子粒数/供试种子总粒数)×100%;
发芽势(%)=(达到高峰时正常发芽种子粒数/供试种子总粒数)×100%;
发芽指数=∑(Gt/Dt),式中,Gt为不同发芽时间(t)的发芽率,Dt为不同发芽试验时间(d)。
1.3 数据统计与分析
应用Microsoft Excel 2007与SPSS 22.0软件进行数据统计,计算楠属4种植物果实、种子表型性状及发芽特性的平均值、标准偏差、变异系数,并进行相关性分析、主成分分析及聚类分析等。
2 结果与分析
2.1 楠属4种植物果实、种子表型性状及发芽特性分析
由表2可知,桢楠的果实长、宽均值最大,分别为14.65 mm和7.82 mm,其中,果实最长的为PZ-YA种源(16.65 mm),果实最宽的为PZ-DJY种源(8.84 mm)。果实百粒重由大到小依次为桢楠(59.34 g)、闽楠(54.46 g)、浙江楠(49.99 g)、紫楠(40.69 g)。而果皮含水率由大到小依次为紫楠(70.60%)、浙江楠(69.47%)、闽楠(65.68%)、桢楠(54.34%)。楠属4种植物的种子长、宽与其果实长、宽表现一致,且种子百粒重与果实百粒重表现也一致,由大到小依次为桢楠(36.02 g)、闽楠(30.09 g)、浙江楠(29.15 g)、紫楠(27.27 g)。楠属4种植物发芽率由高到低依次为桢楠(88.48%)、闽楠(88.05%)、紫楠(85.76%)、浙江楠(84.48%),其中发芽率最高的是PZ-YA、PB-YX两种源,均达到91.79%。发芽势由高到低依次为桢楠(52.25%)、闽楠(48.94%)、紫楠(47.32%)、浙江楠(45.85%),而发芽指数由高到低则为桢楠(5.78)、紫楠(5.71)、闽楠(5.58)、浙江楠(5.10)。

表2 楠属4种植物果实、种子表型性状及发芽特性的变异情况(平均值±标准偏差)
2.2 楠属4种植物果实、种子表型性状及发芽特性的变异系数
变异系数(CV)能反映楠属4种植物不同种源各性状间的变异程度,变异系数越大,离散程度越大。由表3可见,楠属4种植物各性状平均变异系数由大到小依次为桢楠(11.85%)、闽楠(7.74%)、浙江楠(7.05%)、紫楠(6.64%)。桢楠各性状中变异系数最大的为种子长宽比(17.44%),最小的为发芽率(4.45%);闽楠最大的为种子长宽比(13.20%),最小的为果皮含水率(4.55%);浙江楠最大的为单粒种子体积(11.79%),最小的为果皮含水率(2.90%);且紫楠最大的为单粒种子体积(13.22 cm3),最小的为果皮含水率(2.82%)。同一树种4个不同种源的平均变异系数也存在明显差异,例如,闽楠最大的为PB-SC种源(6.46%),最小的为PB-TH种源(4.68%)。

表3 楠属4种植物16个群体的性状变异系数
2.3 楠属4种植物果实、种子表型性状及发芽特性的相关性分析
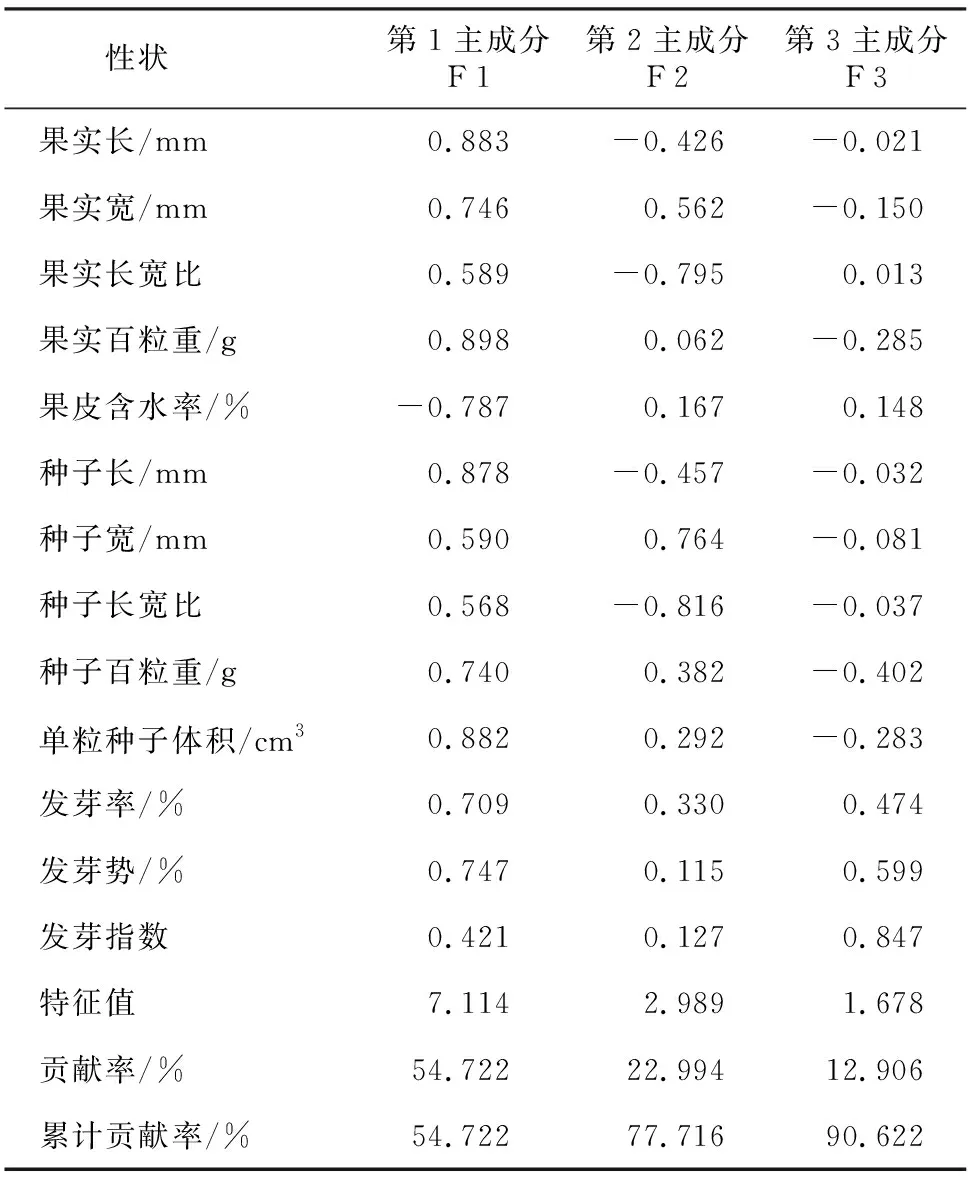
果实、种子表型性状及发芽特性之间相关分析结果(表4)表明,果实长与果实长宽比、果实百粒重、种子长、种子长宽比、种子体积均呈极显著正相关(p<0.01),果实长与果皮含水率呈极显著负相关(r=-0.709;p=0.002);果实宽与果实百粒重、种子宽、种子百粒重、种子体积均呈显著负相关(0.01 表4 楠属4种植物果实、种子表型性状及发芽特性的相关性分析 对楠属4种植物果实、种子表型性状及发芽特性进行主成分分析。由表5可见,前3个主成分累计贡献率达到90.622%。其中,第1主成分的特征值为7.114,方差贡献率为54.722%,主要反映果实长、果实百粒重、种子长、种子百粒重、种子体积的形态特征;第2主成分的特征值为2.989,方差贡献率为22.994%,主要反映果实宽、种子宽的形态特征;第3主成分的特征值为1.678,方差贡献率为12.906%,主要反映发芽率、发芽势、发芽指数的种子发芽特征。 表5 各性状主成分的因子负荷量、特征值、贡献率及累积贡献率 基于果实、种子表型性状及发芽特性,对楠属4种植物16个群体进行UPGMA聚类分析(图1)。以平均欧式距离18 cm为阈值,可将其分为两大类,第一大类为闽楠、浙江楠、紫楠3种植物的12个群体;第二大类为桢楠的4个群体PZ-YC、PZ-LF、PZ-YA、PZ-DJY。在第一大类中,浙江楠的4个群体PC-HZ、PC-QY、PC-NB、PC-KH可聚为一类,闽楠的3个群体PB-TH、PB-YX、PB-JD可聚为一类,紫楠的3个群体PS-LA、PS-JD、PS-SC可聚为一类,而PS-NJ、PB-SC则单独为一类群。 图1 基于表型性状及发芽特性的16个群体聚类结果 林木种实表型性状是相对稳定的遗传特征,是遗传变异的直接表现形式[18]。经过长期自然选择,种实性状表现出了对环境变化的适应性,因此其性状研究对林木具有重要意义[19]。本研究结果表明,种实性状在楠属4种植物种间、种内均存在明显变异,这与李因刚等[20]的研究结果一致;此外,发芽特性在种间、种内存在明显变异,这与谢英赞等[21]的研究结果相一致。楠属4种植物各性状平均变异系数由大到小依次为桢楠(11.85%)、闽楠(7.74%)、浙江楠(7.05%)、紫楠(6.64%),均属于轻度离散,性状相对稳定[22]。Li等[19]研究指出,闽楠叶片、种子表型性状平均变异系数为12.12%,远高于本研究中闽楠的7.74%,这是由于本研究仅测定了闽楠种实和发芽性状,而未测定叶片表型性状,这也进一步证实了多数植物种实性状比叶片性状更稳定[20,23]。 各性状间相关分析结果显示,楠属植物的果皮含水率与果实百粒重、种子百粒重呈显著负相关,且与果实长、种子长和种子体积均呈极显著负相关,这说明果实、种子越长或越重,其果皮含水率则越低。种子发芽率与果实宽、种子宽呈极显著正相关,且与果实百粒重、种子体积呈显著正相关,表明楠属植物种子颗粒越饱满,质量越大,发芽能力越强,这与砂生槐(Sophoramoorcroftiana)[24]、红皮云杉(Piceakoraiensis)[25]等种子发芽特性相似。然而,发芽势则与果皮含水率呈显著负相关,因此可以推断楠属植物因其果实为浆果,果皮含水越多,可能会在一定程度上降低种子活力而影响正常发芽。主成分分析表明,第1主成分、第2主成分均反映出果实、种子的形态特征,同时,聚类分析结果显示,楠属4种植物16个群体基本按照物种聚为一类,说明通过果实、种子性状对不同楠属物种进行植物学分类的可行性[26]。 楠属植物作为长江以南地区优良珍贵常绿阔叶树种,在用材、绿化等方面颇具发展前景和应用价值。其中,桢楠、闽楠、浙江楠均已被列为国家二级保护渐危种,尽管如此,因遭受乱砍滥伐、生境破坏等原因,现存楠属植物天然资源已经十分稀缺。因此,对楠属植物资源保护与合理利用刻不容缓。基于楠属植物种实性状及发芽特性变异丰富的特点,选择优良群体或个体进行采种培育,并运用扦插、嫁接、体胚等无性快繁技术保存优株;在楠属植物采种时应选择颗粒饱满,质量体积均较大的种实进行采种播种,可提高其发芽率;进一步观测本研究中楠属4种植物苗期生长状况,为良种选育和资源保护提供依据。
2.4 主成分分析
2.5 聚类分析

3 结论与讨论
猜你喜欢
中国现代医生(2022年21期)2022-08-22
天津医科大学学报(2021年1期)2021-01-26
医药前沿(2020年20期)2020-11-10
上海农业科技(2020年2期)2020-04-02
三农资讯半月报(2020年2期)2020-03-09
山西农业科学(2020年1期)2020-02-27
小学生学习指导(低年级)(2019年9期)2019-09-25
小学生优秀作文(低年级)(2018年12期)2018-12-13
农民致富之友(2018年9期)2018-06-27
中华手工(2016年4期)2016-04-20
