
小麦种质GLM 1701蓝粒性状的遗传定位分析
2021-01-16 13:07徐如宏任明见
种子 2020年12期
彭 琴, 周 军, 徐如宏, 何 方, 任明见
(贵州大学农学院/国家小麦改良中心贵州分中心, 贵阳 550025)
蓝粒小麦作为糊粉层中富含蓝色花青素的特色小麦种质,与普通小麦相比, 含有较高的蛋白质和赖氨酸,铜、铁、锰、锌4种有色微量元素,且具有很强的抗氧化性[1-4]。高建伟等研究发现,蓝粒小麦的籽粒蓝色常常与许多优良的农艺性状连锁[5];Li Z S等经过连锁遗传分析发现,粗杆、宽叶、直叶、深绿色及半冬性等许多农艺性状与蓝色胚乳性状密切相关[6]。Jia Y.等发现,蓝色花青素只在大麦、小麦和黑麦中积累,而且小麦族蓝色籽粒是一个新的性状,最近才进化出来[7]。目前蓝粒性状已被用作理论研究和小麦育种中的遗传标记,用于测量杂交频率,鉴定真正的杂种,并监测染色体变化[8-13]。
蓝粒小麦籽粒之所以呈现蓝色,是由于其糊粉层中含有蓝色色素,使种子呈深蓝色,呈现胚乳直感、显性遗传,并具有明显的剂量效应[14]。国内外学者对控制蓝粒性状的遗传规律展开了大量研究,但是不同来源的蓝粒小麦控制籽粒色素基因数目等基因源问题还存在争议。相志国等研究表明,蓝粒在遗传过程中有母本效应,类似一对基因控制的遗传行为,蓝粒性状基因的表达有明显的剂量效应,相同的蓝粒基因在与小麦中不同的染色体发生代换,其效应完全不同[15];同一基因控制着的蓝粒可划分为深、中、浅三类,在不同的环境条件下种植,也表现出一定的差别[16]。黄碧光等研究表明,蓝粒的遗传受1对基因控制,蓝粒为显性,且控制蓝粒的基因具有剂量效应[17]。Hurd等认为,蓝粒受2个互补的不完全显性基因控制[18];Keppenne等认为,蓝粒受2对呈抑制互作的基因控制[19]。兰素缺等研究发现,来源于偃麦草的D 87065和D 87089的籽粒色素基因由2对互补基因控制;来源于黑麦的92-1由2对互补基因控制;来源不明确的7083 L-16由1对基因控制[20]。目前已经定位的小麦蓝色糊粉层基因有来自长穗堰麦草的4 AgL染色体的Bal基因[21],来源于百萨堰麦草的BaThb位于4 JL[22];来源于栽培一粒和野生一粒的Ba2位于4 AL[23-24];还未定名的,源于中间偃麦草,位于4 J(FL 0.60-1.00)[25]。
BSA(Bulked Segregant Analysis)称为混合群体分离分析法,是一种通过极端性状进行功能基因挖掘的方法。该方法是用合适的分子标记对2个基因池进行分析,在2个基因池间存在多态性的标记与目标性状的基因座连锁[26-27]。BSA虽然SSR标记具有高可靠性、稳定性和良好重复性的优点,但是它们在基因定位中与BSA(例如RAPD,AFLP,SSR)结合使用有许多分子标记,它们对某种变化具有特异性。对基因进行靶向定位仍然是耗时且劳动密集的,并且难以同时进行大量料或靶基因的大规模定位筛选。随着DNA测序技术的发展,SNP标记被发现且广泛用于构建动物和植物遗传连锁图谱,SNP标记具有数量多,密度高,遗传稳定性高,易于自动检测等优点。因此,SNP正在迅速取代传统标记,例如RFLP和SSR。BSA+基因芯片方法可以快速检测2种不同表型之间的遗传差异[28-29]。SNP突变及其染色体片段比全基因组基因定位更准确。目前很多研究者通过这种方法完成了许多基因的定位,赵秋实等[30]通过BSA法结合660 K基因芯片以及SSR分子标记技术成功将矮杆基因定位于1 A染色体上。蒋宏宝等[31]利用BSA法结合小麦660 K基因芯片将小麦叶绿素缺失突变体B 23定位于7 AL染色体上。
GLM 1701是贵州大学农学院国家小麦改良中心贵州分中心通过远缘复合杂交创制的蓝粒小麦新种质。本研究通过常规杂交构建遗传群体结合660 K基因芯片技术进行遗传分析和基因定位,以期为合理有效地利用蓝粒小麦新种质GLM 1701提供科学理论依据。
1 材料与方法
1.1 供试材料
本研究所用材料为GLM 1701(蓝粒)、贵农19(白粒)、贵农麦30号(白粒)、中燕96-3(白粒)和绵麦301(白粒),均由贵州大学农学院国家小麦改良中心贵州分中心提供,并在该中心试验地种植。
1.2 遗传群体构建
蓝粒小麦品种GLM 1701分别与白粒小麦品种贵农19号、贵农麦30号、中燕96-3和绵麦301进行正反杂交构建F1群体,将收获的F1世代夏播加代繁殖,对F1世代群体单株进行套袋自交,构建F2分离群体,于2018年11月将F2群体种植在贵州大学教学实验场国家小麦改良中心贵州分中心小麦育种科研基地,于2019年3月底3叶时进行单株编号,待长到4~5叶时,每单株取1~2片叶备提取DNA,构建F2∶3群体,5月份按单株收获,并进行单株粒色统计。
1.3 遗传分析
将收获的全部F1,BC1F1、F2中群体中不能区分的蓝色籽粒和籽粒横切成两半,含胚的一半用来继续种植,另一半用来观察糊粉层颜色,统计F1、BC1F1、F2群体各个组合蓝粒、白粒比例。
1.4 基因定位及群体验证
根据F3单株粒色结果,在贵农19×GLM 1701组合中选取F2纯蓝粒和纯白粒各40个单株叶片分别组建颜色性状混池,利用贾继增等与Affymetrix联合研发的660 K基因芯片对2个亲本和BSA混池进行SNP基因分型,这部分工作主要在北京博奥生物技术有限公司完成。
根据芯片结果,对贵农19×GLM 1701组合双亲及混池进行筛选,找出具有多态性的标记对其F2∶3群体进行基因定位和验证。单株叶片DNA提取参照徐如宏等[32]改良的CTAB法提取基因组。PCR反应体系为10 μL:模板DNA(100 ng·μL-1)2.0 μL、primer(10 μmol·L-1)1.0 μL、TagE(5 U·μl-1)0.1 μL、dNTP(2.5 mmol·L-1)0.2 μL、10×Buffer(Mg2+)(5 mmol·L-1)1.2 μL、ddH2O 4.5 μL。反应程序:94 ℃预变性3 min;94 ℃变性1 min, 50~60 ℃(根据不同引物决定)退火1 min,72 ℃延伸2 min,35个循环;72 ℃延伸10 min,4 ℃保存。 PCR扩增产物采用8%的非变性的聚丙烯酰胺凝胶,银染法检测。显影后,在白炽灯下观察电泳结果,进行条带统计和照相。
2 结果与分析
2.1 遗传群体籽粒颜色性状的表现
利用蓝粒小麦种质(GLM 1701)与白粒小麦品种(贵农19号、贵农30号、中燕96-3、绵麦301)作为亲本,分别组配正反交F1、F2和BC1F1群体,对各个群体籽粒颜色性状进行观察统计(表1)。

表1 相关世代群体籽粒颜色性状的表现
由表1可知,F1世代籽粒颜色深浅与蓝粒亲本作为父本或母本有关,当蓝粒亲本GLM 1701作为母本时,F1籽粒颜色表现为中蓝色,当蓝粒亲本GLM 1701作为父本时,F1籽粒颜色表现为浅蓝色,说明蓝粒为显性,存在剂量效应。正反交F2世代籽粒的颜色均表现出蓝白粒分离;回交中正反交F1与白粒亲本进行正反回交,BC1F1都是出现蓝粒和白粒,正反交F1与蓝粒亲本进行正反回交,BC1F1只有蓝粒。结果表明,GLM 1701的蓝粒性状为显性性状,且存在剂量效应。
2.2 遗传群体F2代群体籽粒颜色的分析
利用GLM 1701与贵农19号、贵农30号、中燕96-3、绵麦301进行正反交,调查统计其正反交F2群体的籽粒颜色,通过卡方检验分析蓝粒与白粒的分离比例(表2)。

表2 F2代群体籽粒颜色的分离情况
由表2可知,各杂交组合F2发生蓝粒和白粒的分离,且蓝粒与非蓝粒的数目相当,分离比例为5∶4,蓝粒有深浅之分,体现剂量效应。
2.3 遗传群体BC1F1群体籽粒颜色的分析
利用GLM 1701与绵麦301、贵农30号、中燕96-3和贵农19号进行正反交构建F1群体,正反交F1再与双亲进行正反回交,调查统计其回交群体BC1F1的籽粒颜色(表3)。
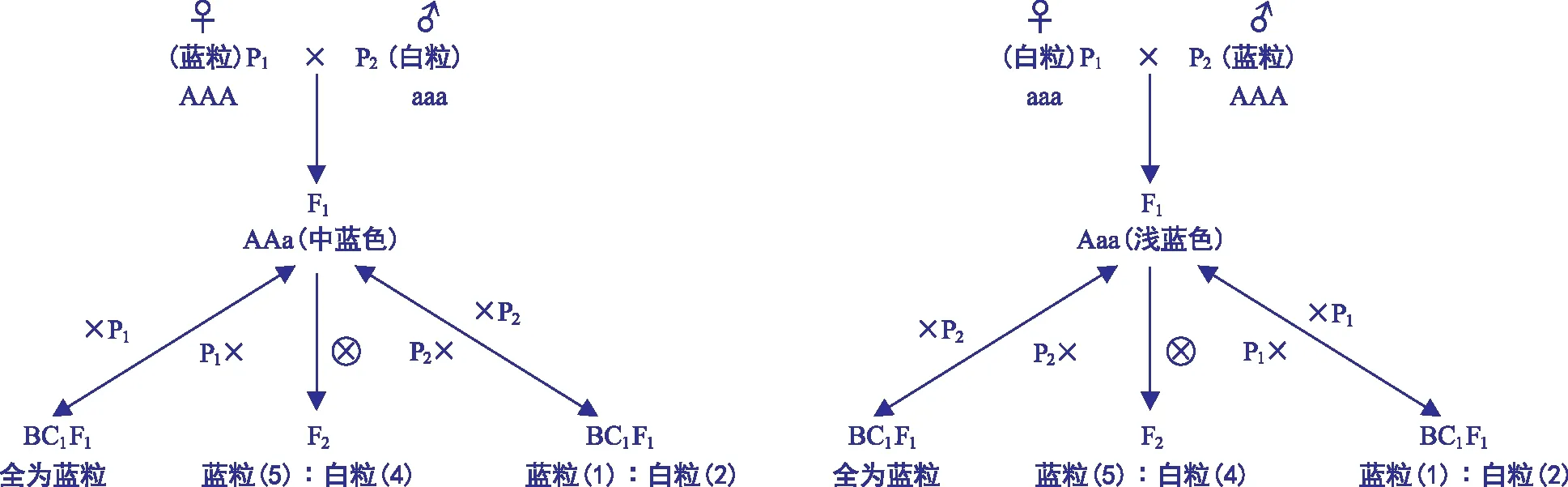
图1GLM 1701蓝粒性状胚乳基因的遗传模式
由表3可知,正反交F1与白粒亲本进行正反回交,BC1F1代群体蓝粒∶白粒为1∶2,正反交F1与GLM 1701进行正反回交,BC1F1代群体全为蓝粒。

表3 BC1F1代群体籽粒颜色的分离情况
2.4 F2分离比例的验证
综合回交群体分析表明,当正反交F1作为母本与白粒亲本回交时,BC1F1群体蓝粒与白粒的比例都是1∶2,说明含蓝粒基因的配子约占雌配子总数的33%,而理论上含蓝粒基因的雄配子占雌配子总数的50%,因此蓝粒基因通过雌配子的传递率约为67%(33%/50%)。同样当正反交F1作为父本与白粒亲本回交,BC1F1群体蓝粒与白粒的比例同是1∶2,说明蓝粒基因通过雌配子的传递率也约为67%(33%/50%)。综合分析得出,含蓝粒基因的配子约占雌或雄配子总数的33%,传递率均只有67%。
因含蓝粒基因的雌、雄配子的传递率降低,造成了F2蓝粒与非蓝粒的比例失真,不符合正常的孟德尔遗传,根据测交结果得出的蓝粒基因雌雄传递率推算F1产生2种配子在雌雄配子中的比例,据此可算出F2代各种类型的比例。用A表示蓝粒基因,a表示非蓝粒基因,蓝粒是由2 n的极核与n的精细胞结合而形成的三倍体胚乳糊粉层性状,所以用AA和aa分别表示2种雌配子,三核胚乳的基因型有4种:AAA、AAa、Aaa和aaa。从表4可看出,在F2中,A_∶aa,即蓝粒∶非蓝粒=(1+2+2)∶4 =5∶4,理论上蓝粒在F2中的比例为56%,非蓝粒为44%。这与实际中F2非蓝粒与蓝粒数目相当的情况相吻合。各组合F2蓝粒与非蓝粒的比例通过卡方验证p>0.5(表2)。说明正是含蓝粒基因雌雄配子传递率的下降,造成F2蓝与非蓝粒比例不符合3∶1。综合自交F2和回交BC1F1得出,GLM 1701的蓝粒性状是由一对主效基因控制。

表4 F1代2种配子在雌雄配子中的比例与F2代4种胚乳基因型的理论比例
假设蓝粒小麦种质GLM 1701胚乳的基因型为AAA,白粒小麦品种贵农30号和贵农19号胚乳的基因型为aaa,其胚乳遗传模式如图1所示。
2.5 蓝粒基因定位及验证
2.5.1SNP芯片测序结果分析
运用小麦660 K基因芯片分别对贵农19×Glm 1701组合2个极端混池以及双亲进行全基因组检测。将在2个混池以及双亲间均表现差异的SNP进行比较整合,基于质控后获得2个混池及2个亲本间共同表现差异的SNP共有301个。根据660 K芯片遗传整合图谱信息,将表现出差异的SNP整合到小麦21条染色体上(图2)。由图2可知,1 A、3 A、1 B、3 B、5 D五条染色体上没有差异SNP,4 D染色体上差异SNP分布达到205个,约占总SNP的68%;4 B、4 A和6 D染色体上分别分布13个差异SNP,5 A染色体上分布26个差异SNP,约占总SNP的22%,其余的差异SNP分散分布在其他染色体。推测控制蓝粒性状的基因位于4 D染色体上。

图2 差异SNP的染色体分布

注:Maker为100 bp DNA Ladder,1~48为F2群体里蓝粒单株,目标条带位置为194 bp左右。

注:Maker为100 bp DNA Ladder,1~48为F2群体里白粒单株。
2.5.2基因定位及群体验证
根据芯片结果选用位于4 D染色体上[33-34]的58对标记对贵农19×GLM 1701组合双亲及混池进行筛选,发现Xgwm 165标记能在双亲和混池中有差异。
将特异性标记Xgwm 165对贵农19×GLM 1701正反交F2群体进行PCR分析,所获结果见表5。根据表5数据,用软件Joinmap 4.0计算,Xgwm 165标记与GLM 1701遗传距离为5.8 cM。此结果证明控制GLM 1701蓝粒性状的基因位于4 D染色体。

表5 群体交换单株统计
同时,用其他各个组合的F2∶3群体对标记Xgwm 165进行验证,部分结果见图3、图4。在后代材料的检测中,除其中1株未扩增出目标条带,其它均能扩增出194 bp目标条带,与F2粒色统计结果基本一致,说明标记Xgwm 165与蓝粒基因是连锁的。
2.5.3特性标记Xgwm 165定位
对特异性标记Xgwm 165的PCR产物进行T载体克隆,在成都擎科生物技术有限公司完成。克隆得到Xgwm 165的序列为:TGCAGTGGTCAGAGTTTTCCCAGGCCACAGATGAGCAAAAGAAATCTTTTGTTGTCCAGCGTTGTCTAATCTTGCCTTGCGAATAGTACTTTTGAGCAGCTCAAGGAGAGAGAGGGAGAGAGAGAGAGAGAGAGCGCGCGCGCAAACAAAGATAGCAAAGAATTAGGCAACACTGGCGCAATCTGAAAGAAAAG,基于JBrowse网站(网址:http:202.194.139.32/jbrowse),比对中国春(IWGSC reference V1.0 all Chromosomes)版本,得到标记Xgwm 165在4 D染色体上的位置为412716481~412716679 Mb。
3 讨 论
蓝粒小麦的籽粒之所以表现出蓝色特性,主要原因是其籽粒的胚乳糊粉层中富含花色素[34-35]。胚乳是由1个精核(n)与2个极核(n+n)受精结合为胚乳核(三倍体),然后发育成胚乳。蓝粒亲本与白粒亲本杂交,F1均为中蓝(蓝粒小麦作为母本)或浅蓝色(蓝粒小麦作为父本)[16]。籽粒颜色深浅通过表型观察误差较大,所以本研究对蓝粒小麦的籽粒进行横切,含胚的一半继续种植,另外一半用来观察糊粉层颜色。
本研究以蓝粒小麦种质GLM 1701分别与白粒小麦品种贵农19号、贵农30号、中燕96-3和绵麦301进行正反交及正反回交,对籽粒糊粉层颜色的观察统计表明,F1籽粒颜色与李振声等[16]的研究结果一致;正反交F2世代蓝粒与白粒的比例为5∶4;正反交F1与白粒亲本进行正反回交,BC1F1群体中出现蓝粒∶白粒为1∶2的比例,正反交F1与蓝粒亲本进行正反回交,BC1F1群体则全为蓝粒,综合分析BC1F1群体和对自交F2群体的验证,发现含蓝粒基因的雌雄配子的传递率分别只有67%,因此造成自交F2和回交群体BC1F1偏离正常的孟德尔遗传,通过F1、F2和BC1F1世代群体遗传规律分析得出GLM 1701蓝粒性状的基因是一对基因控制,与李振声等[16]、黄碧光等[17]、兰素缺等[20]和Knievel等[36]的研究结果一致。有所不同的是,黄碧光等研究得出,含蓝粒基因的雌配子传递正常,而雄配子的传递率只有20%,自交F2为11∶9的比例。Knott等研究发现,雌蕊功能表现为正常的情况下,携带偃麦草染色体的花粉功能弱于不携带偃麦草的染色体的花粉[37]。傅体华等也认为,含蓝粒基因的配子传递率会下降[ 38];Keppenne等认为,F2符合1∶2∶1的特点[19]。控制蓝粒性状的染色体由于种间杂交可能被群体分离成碎片或含有整个供体亲本染色体,且与小麦中不同的染色体发生代换,其效应完全不同[7,15,37],且由于花粉直感和环境变化的影响,以及其他一些不确定因素影响蓝粒小麦蓝粒颜色的表型,导致蓝色种子表面形成条纹或斑点,干扰了对蓝粒性状的观察统计,从而导致出现不同的研究结果。除此之外,蓝粒基因的表达还会受到其他基因的影响[39-42]。
利用BSA混池法结合660 K基因芯片能够快速和高效地进行目标基因的定位。本试验中芯片结果显示,控制蓝粒性状的基因位于4 D染色体,不同于之前的研究结果,可能是一个新的基因,且筛选到一个特异性标记Xgwm 165,位于4 D染色体的长臂上,遗传距离为5.8 cM,本研究将控制蓝粒性状的基因命名为LM1。对Xgwm 165标记进行T载体克隆,比对中国春得到的物理位置为412716481~412716679 Mb。此结果可为GLM 1701蓝粒小麦的进一步利用提供基础。
猜你喜欢
西南农业学报(2022年5期)2022-06-06
中国农学通报(2022年13期)2022-05-31
热带亚热带植物学报(2022年2期)2022-04-14
现代畜牧科技(2021年4期)2021-12-05
广东农业科学(2021年3期)2021-04-23
山东建筑大学学报(2020年5期)2020-09-16
科学之谜(2019年3期)2019-03-28
热带农业科技(2019年1期)2019-01-14
科学之谜(2018年8期)2018-09-29
科学种养(2017年6期)2017-06-13
