
西双版纳热带植物园蝙蝠物种多样性与季节变化研究
2021-01-13 04:01杨建波鲁建美AliceHUGHES
野生动物学报 2021年1期
曹 慧 杨建波 鲁建美 Alice C.HUGHES
(1.中国科学院西双版纳热带植物园,勐腊,666303;2.中国科学院西双版纳热带植物园综合保护中心,勐腊,666303;3.中国科学院大学,北京,100049)
目前蝙蝠已知物种数超过1 400多种,且具有广泛分布[1],常作为种子和花粉传播的重要媒介,也是害虫防治的关键因子,提供重要的生态系统服务功能。蝙蝠具有较高的物种特异性,并根据自身生活史特征选择适宜的环境进行栖息、捕食、交流等活动[2]。不同物种对栖息地和捕食场所的选择有明显差异,温带地区气温较低,蝙蝠经常选择岩石缝隙和洞穴作为长久栖息地,甚至是空旷、干扰较弱的人为建筑也会成蝙蝠的固定栖息地,从而在城市中的活动水平有明显上升[3],成为城市中物种多样性最高的哺乳动物类群[4-5]。热带地区已有超过100多个物种在同一地区被记录,并在热带森林环境中物种多样性达到最高[6-7]。
热带地区喀斯特地貌上常发育长势良好的热带雨林,这对热带蝙蝠的生存至关重要。不仅为洞栖蝙蝠,如菊头蝠科(Rhinolophidae)、蹄蝠科(Hipposideridae)提供捕食场所和食物资源,也为树栖蝙蝠提供赖以生存的栖息环境。例如哈氏彩蝠(Kerivoulahardwickii)栖息在蜷缩的芭蕉叶中,扁褐颅蝠(Tylonycterisrobustula)栖息在竹洞中,狐蝠科(Pteropodidae)物种栖息在树干上,棕果蝠(Rousettusleschenaultii)栖息在棕榈树叶下。热带地区年内温度变化不大,但降水量丰富的热带区域性气候对全球气候变化起着决定性的作用,并影响森林覆盖面积[8],从而影响蝙蝠活动。例如随着旱季末期的到来,温度达到最高,墨西哥的一种大食果蝠(Artibeusintermedius)和幽暗美洲果蝠(Dermanuraphaeotis)开始进行交配,以确保在雨季来临之前完成生殖,利用雨季中丰富的食物供给自身能量需要,并抚育幼崽[9]。这种温度和降水量的变化,是季节性繁殖物种得以生存的关键因素。季节性繁殖也导致了栖息地利用的季节性变化,影响蝙蝠种群结构和物种组成[10]。生殖季节能量需求的提高,雌性蝙蝠常选择多种不同的生态环境进行活动,例如人为修建的公路、水坝、稻田都可作为蝙蝠栖息和捕食场所,生态交错区和收割之后的荒地也会由于食物资源可利用性的增加,蝙蝠种群密度有所提高[11]。因此植被覆盖程度和栖息地类型转换在蝙蝠季节性繁育过程中具有重要的生态意义。
热带地区具有最高的森林砍伐率,森林断层的不断增加严重威胁热带地区生态环境,热带雨林的破坏程度也往往被人们所忽视[12]。例如在热带雨林地区不断出现没有被记录在道路测绘网络中的小路[13],造成对森林破碎化程度的低估。同时东南亚地区的一些国家,仍然保持着刀耕火种的传统耕作方式,季节性的改变森林覆盖面积,影响当地蝙蝠物种组成。
然而目前热带地区蝙蝠物种多样性的研究甚少开展,尤其在西双版纳地区。本研究将探究西双版纳热带植物园热带雨林中蝙蝠物种组成及物种多样性的季节变化,揭示热带森林破碎化及季节变化对蝙蝠活动的影响,为今后热带地区发展和生物多样性保护提供理论依据。
1 研究区域和研究方法
1.1 研究区域
实验在中国科学院西双版纳热带植物园中进行(101°15′56″E,21°55′36″N),该地区森林植被主要为热带雨林。气候具有明显的干湿两季,年平均降水5 157 mm,旱季降水主要以浓雾的形式。年均温24℃,最热月平均温25.3℃,最冷月平均温15.6℃。
1.2 研究方法
自2019年1—12月,实验共进行201 d(表1)。选择1个被动型声学监测设备(AnaBat Express,Titley Scientific,Australia),架设高度为2 m,考虑蝙蝠常沿线形要素飞行的特点,该设备被垂直并顺着道路方向架设。通过使用温湿度监测设备HygrochronTMiButton(DS1923)(Wdsen Electronic Technology Co.,Ltd)记录环境变化。实验时间为日落前1 h至日出后2 h。通过Anabat Insight(version 1.1.5)软件分析超声波,利用以下参数:主频率、最大频率、最低频率、平均频率、持续时间以及波形,进行蝙蝠物种鉴别。由于一些物种超声波持续时间较长,如马来亚菊头蝠(Rhinolophusmalayanus),或重复在同一区域活动,可能造成单个个体的过度估计。为减小误差,将取样夜晚分为1 min时间间隔,统计每个物种被记录到的1 min时间间隔超声波次数,以表达蝙蝠物种多样性和活动强度[14]。使用R语言(version 3.5.3)中多元线性回归模型分析物种活动强度、物种多度与环境变量的相关性。并结合竖琴网和雾网的捕获情况作为探究蝙蝠物种分类的补充数据。
2 研究结果
2.1 物种组成及区系分布
本研究共记录到5科14属32种蝙蝠(表2),包括5种蹄蝠科蝙蝠,12种蝙蝠科(Vespertilionidae)蝙蝠,11种菊头蝠科蝙蝠和4种狐蝠科蝙蝠,其中哈氏彩蝠,丘彩蝠(K.papillosa),马来假吸血蝠(Megadermaspasma),毛翼管鼻蝠(Harpiocephalusharpia),圆耳管鼻蝠(Murinacyclotis),艾氏管鼻蝠(Mu.eleryi),霍氏鼠耳蝠(Myotishorsfieldi),高颅鼠耳蝠(My.siligorensis),斑蝠(Scotomaresornatus),中菊头蝠(Rh.affinis),婆罗菊头蝠(Rh.borneensi),小菊头蝠(Rh.pusillus),半岛菊头蝠(Rh.robinsoni),托式菊头蝠(Rh.thomasi),棕果蝠,安氏长舌果蝠(Macroglossussobrinus),犬蝠(Cynopterussphinx),长舌果蝠(Eonycterisspelaea),这18种蝙蝠为通过竖琴网和雾网捕捉到的记录。其余15种为声学监测设备记录到的物种,其中某种管鼻蝠(Murinasp.)只鉴别到属的水平。研究发现,32种蝙蝠不仅具有东洋界分布的特征,并且华南水鼠耳蝠(Myotislaniger),中华菊头蝠(Rh.sinicus),棕果蝠也有在古北界青藏区分布的记录[15-18]。
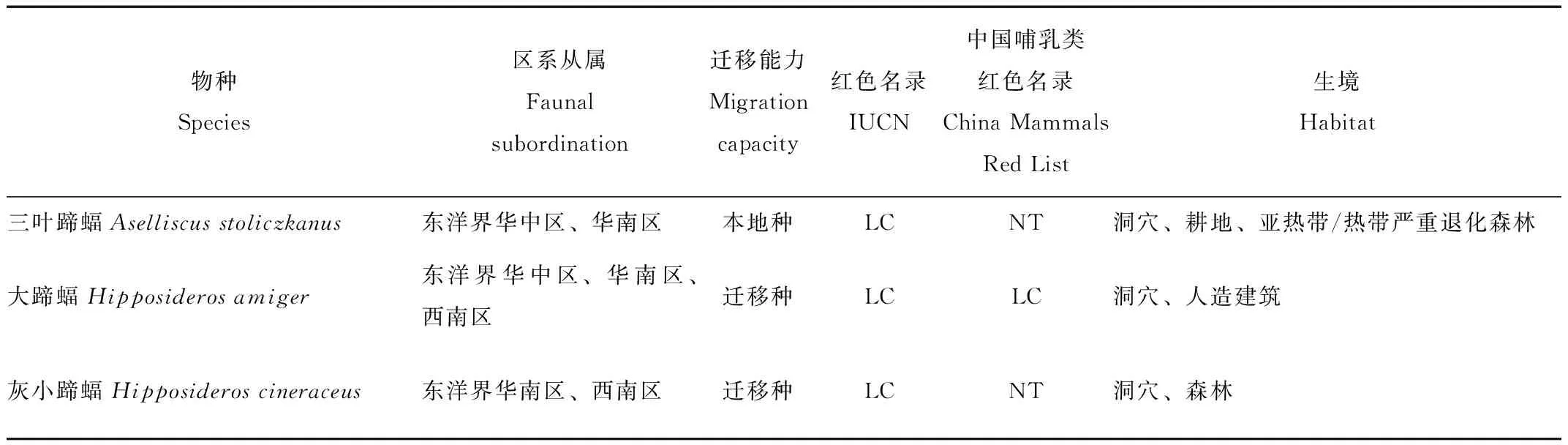
表2 蝙蝠物种分布和迁移能力Tab.2 Bats species distribution and migration capacity
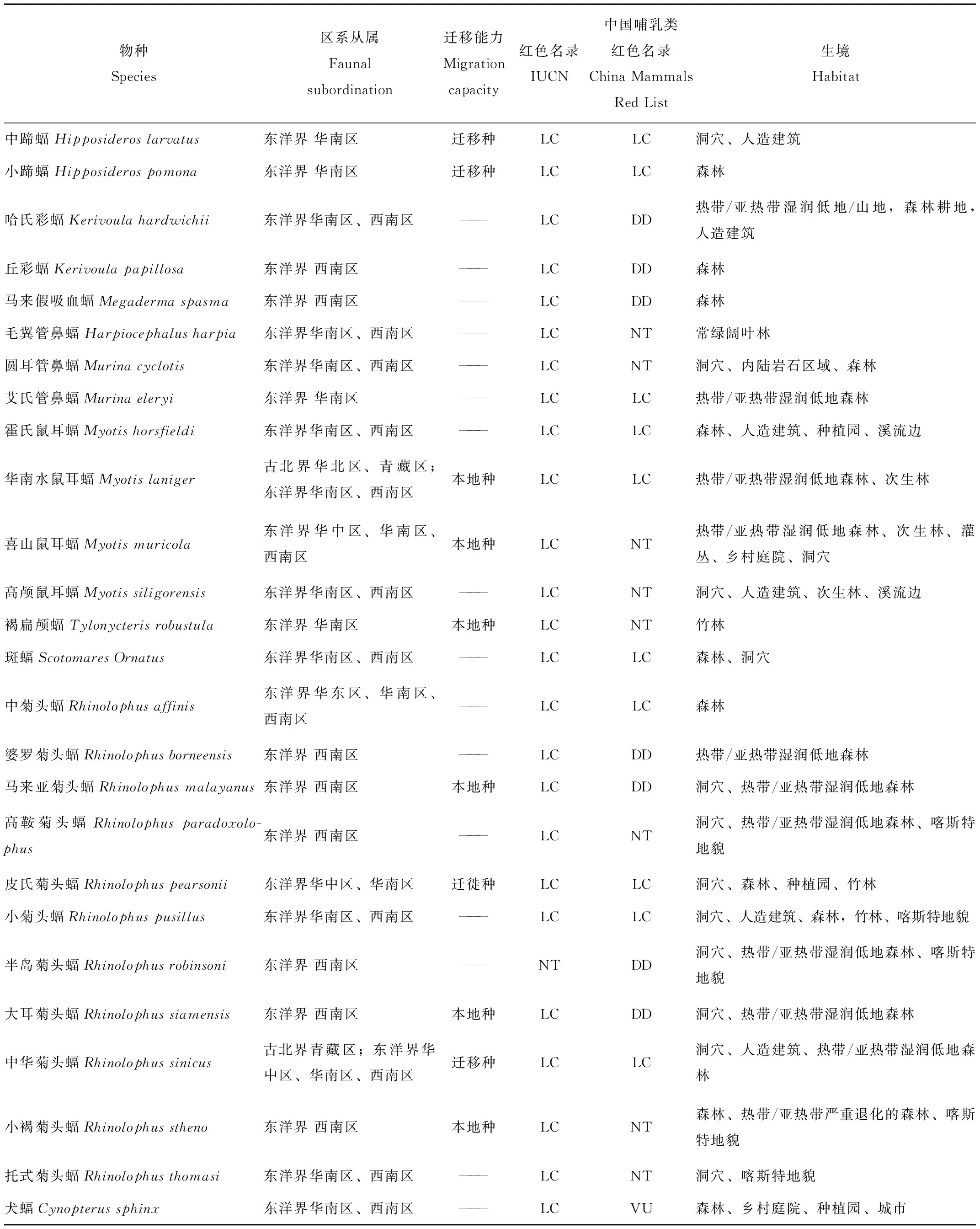
续表2
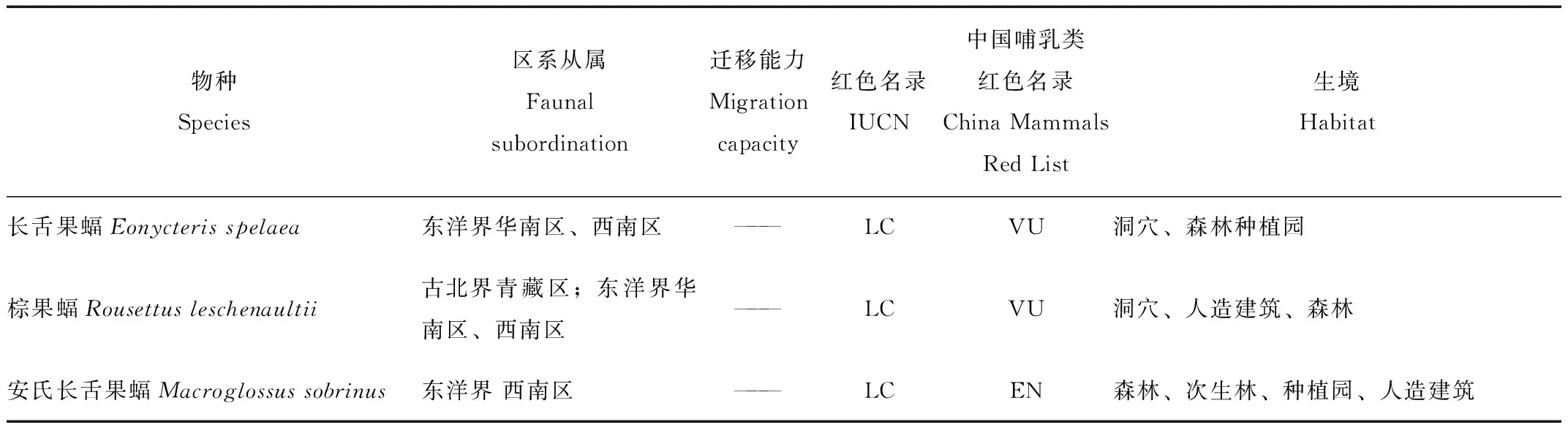
续表2
本研究将使用声学监测设备连续2个月没有探测到的物种记为迁移种,将每月均有记录的物种记为本地种。蹄蝠科中4种蝙蝠显示有迁移特性,占80%,蝙蝠科仅有管鼻蝠被记录到有迁移行为,占25%,菊头蝠科蝙蝠仅有2种蝙蝠被记录有迁移行为,占33%。
2.2 蝙蝠物种多样性与环境变化
2.2.1 蝙蝠物种多样性与活动强度
使用声学监测设备共记录到的15种蝙蝠中,马来亚菊头蝠为全年活动性最强的物种,灰小蹄蝠(Hipposideroscineraceus)活动性最弱。15种蝙蝠全年1 min时间间隔超声波次数占比见图1:马来亚菊头蝠:35.732%,中蹄蝠(Hipposideroslarvatus):20.322%,大耳菊头蝠(Rhinolophussiamensis):10.661%,小褐菊头蝠(Rhinolophusstheno):10.434%,喜山鼠耳蝠(Myotismuricola):6.606%,三叶蹄蝠(Aselliscusstoliczkanus):6.051%,高鞍菊头蝠(Rhinolophusparadoxolophus):4.435%,褐扁颅蝠(Tylonycterisrobustula):4.049%,华南水鼠耳蝠:0.870%,大蹄蝠(Hipposiderosamiger):0.216%,小蹄蝠(Hipposiderospomona):0.201%,皮氏菊头蝠(Rhinolophuspearsonii):0.175%,中华菊头蝠:0.134%,某种管鼻蝠:0.108%,灰小蹄蝠:0.005%。
2.2.2 环境变化与蝙蝠物种多样性的相关性
尽管不同蝙蝠物种活动强度对环境变化的响应不同,但大部分蝙蝠活动强度与温度和湿度的变化显著相关,而与降水量的变化相关性不显著(表3)。蝙蝠科蝙蝠的活动强度常与单一的环境变量有关,例如管鼻蝠仅与湿度变化显著相关,褐扁颅蝠仅与温度变化显著相关。但蹄蝠科和菊头蝠科蝙蝠对环境参数的变化更为敏感,不仅多个物种的活动强度与湿度和温度的变化同时显著相关,而且菊头蝠科小褐菊头蝠的活动强度是唯一对降水量变化显著相关的物种(P<0.01)。同时研究表明蝙蝠物种多度的变化仅与温度表现显著相关(P<0.05)。
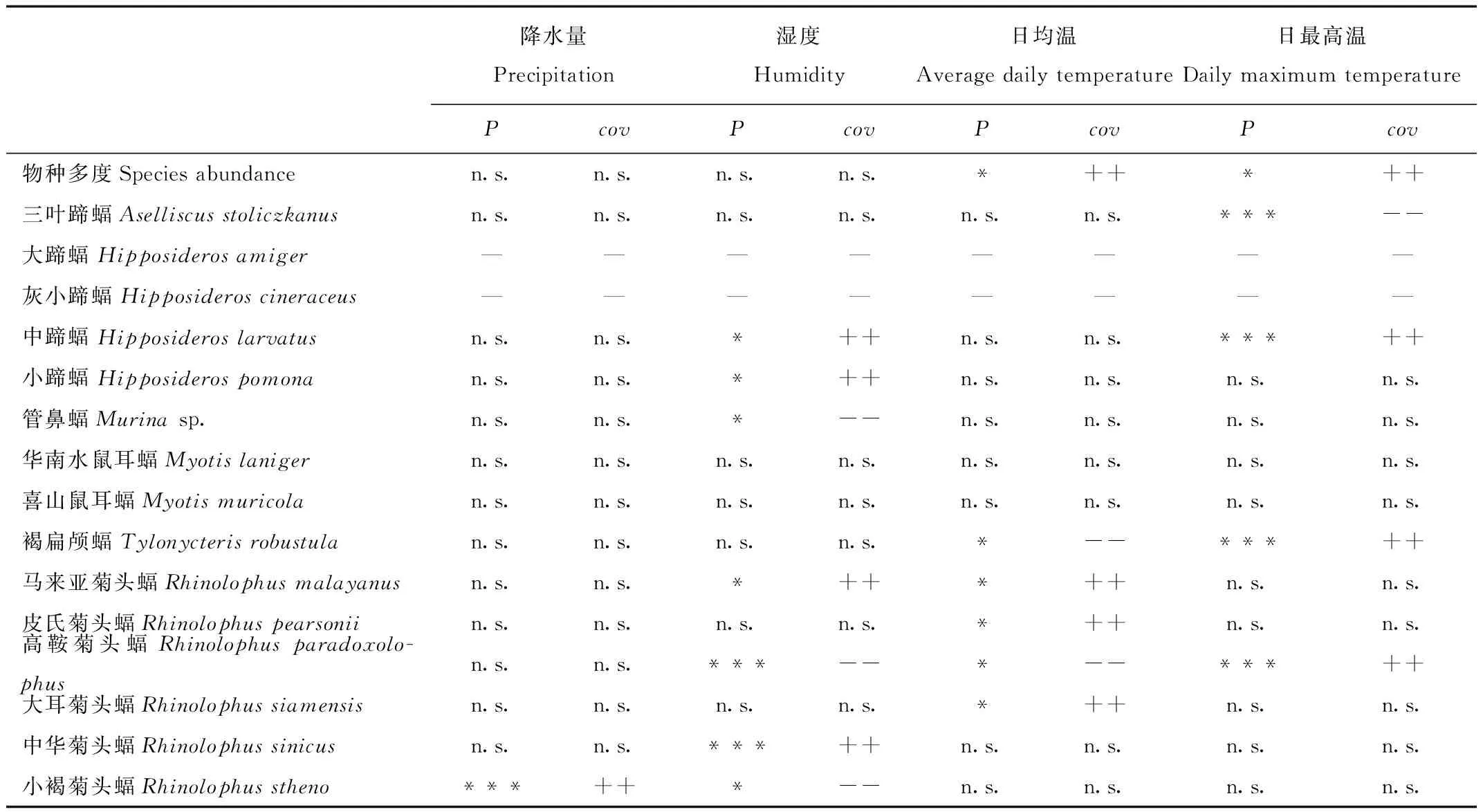
表3 蝙蝠活动强度、物种多度与环境参数的相关性Tab.3 Correlation of bat activity intensity,richness and environmental parameters
2.3 蝙蝠物种多样性的季节变化
声学监测的结果表明,蝙蝠物种多样性的季节变化具有明显的物种特异性(图2)。菊头蝠科蝙蝠物种多样性全年变化最为平稳,有长达7个月的时间物种多度保持不变(3—6月,10—12月),并在雨季前和旱季前期物种多样性较高,而在雨季中期则有相对下降。蝙蝠科蝙蝠物种多样性的季节变化虽同样趋于平稳,但在雨季过后有下降趋势(10月至次年2月)。蹄蝠科蝙蝠物种多样性的季节变化最为明显,雨季中期(7月)达到最高,旱季有明显的下降,1月和2月到达最低。
3 讨论
3.1 蝙蝠物种多样性的季节变化
通过对蝙蝠物种分布和活动强度的分析都表明该地区蝙蝠物种多样性具有明显的热带区域特有性。菊头蝠科和蹄蝠科蝙蝠作为典型的热带物种,其活动强度具有指示热带地区气候变化的属性,在该地区活动强度也相对较高。一些蝙蝠科物种古北界的区系分布特征也体现了西双版纳热带北缘地理位置的特殊性和该地区生物分布由亚热带向热带过渡的趋势。同时蝙蝠活动强度和物种多样性的变化往往更受温度的影响,而与降水量的变化并没有显示出显著相关。雨季来临前,蝙蝠活动强度、物种多样性与日均温达到高值,并随温度的降低而降低。旱季时部分蝙蝠物种有明显的迁徙行为,主要表现在蹄蝠科蝙蝠,5种蹄蝠科蝙蝠中有4种蝙蝠,如大蹄蝠、中蹄蝠、灰小蹄蝠、小蹄蝠连续2个月没有监测到活动,并在以往旱季时期云南地区洞穴考察中,也没有发现该科蝙蝠有冬眠行为。而菊头蝠科和蝙蝠科的活动中,虽没有大规模的迁移行为,但仍有短暂的冬眠行为,导致了在旱季时期蝙蝠物种多样性的下降,这不仅与温度有关,也可能由于该地区旱季时期食物资源可利用性下降,故在旱季时期,热带雨林遭到破坏对蝙蝠物种多样性的危害更大。这也更加证实了全球气候变化对蝙蝠物种多样性影响的重要性和需要对热带地区气候长期监测的必要性。
此外也要考虑声学监测设备可探测性对本研究的影响,如灰小蹄蝠超声波频率较高,不易被探测。热带雨林地区由于相对密闭的环境,也会影响探测范围,蝙蝠活动性可能被低估。
3.2 热带雨林对蝙蝠物种多样性的影响
西双版纳地区森林植被类型主要包括热带雨林、热带季节性湿润林、热带季雨林、热带山地(低山)常绿阔叶林、热带棕榈林、暖热性针叶林和竹林7种[19],其中热带雨林具有最高的乔木物种多样性[20]。热带雨林良好的更新发展对维持当地生物多样性,尤其是对生活在热带雨林中的特有种意义重大。本研究声学监测设备共记录到的15种蝙蝠,菊头蝠科和蹄蝠科的蝙蝠大部分物种为典型的热带物种,例如马来亚菊头蝠。这类蝙蝠依托热带雨林茂密的植被、狭窄的空间和夜晚极低的可见度进行栖息和捕食,“调频/恒频”交替的超声波特征使这类蝙蝠具有短距离监测猎物的能力,以适应在密闭的雨林中生存[21-22]。也正因如此,此类蝙蝠更易受到人为活动的干扰,对热带雨林的动态变化更加敏感。同时研究表明蝙蝠物种多样性在雨季中期有所减少,这可能由于生殖期的结束,蝙蝠种群数量增加,种间竞争加大,促使蝙蝠活动强度增大,扩散能力增强[23-24],继而选择更多样的栖息环境和捕食场所,故森林覆盖面积成为影响该物种组成和结构至关重要的因素。同时也应考虑雨季时由于降水量增大导致昆虫飞行效率降低,飞行成本上升,继而影响蝙蝠的捕食活动,限制捕食时间,造成物种多样性的下降。当森林断层出现时,林内的透光性增强,蝙蝠转移栖息地并回避常规的捕食路线,减少了捕食成功率,增加了捕食成本和被捕食的风险[25]。这一点也在一些研究中得以证实,城市化程度较高的区域,夜晚灯光水平的提升和强烈的人为活动干扰,菊头蝠科和蹄蝠科的蝙蝠更易遭受灭绝的风险[26-27]。并且蹄蝠科蝙蝠多具有狭食性的特点[28],选择鳞翅目(Lepidoptera)昆虫作为主要食物,使它们对生存环境的依赖性更强,对景观类型的转变更加敏感。而不同于菊头蝠科和蹄蝠科的蝙蝠具有洞穴栖息的特性,蝙蝠科的蝙蝠常选择树叶下或有遮挡的岩石缝隙作为栖息场所,从而对热带雨林的环境质量和森林覆盖面积的要求更高。
然而西双版纳地区,经济发展的加速以及夜晚灯光水平的提升,导致森林类型不断由热带雨林向橡胶林、农田和茶园转换[28-29],热带雨林面积占总植被覆盖面积的比例从1992年的22.77%减少至2016年的15.1%[20],热带次生林面积迅速增加,造成森林植被物种组成结构的单一,乔木多样性降低,这为蝙蝠物种多样性的调查和保护的工作带来诸多难题。在蝙蝠栖息地的保护中,进行就地保护,营造合理并且成熟的林分结构是保护蝙蝠物种栖息地的重要手段[30],因此未来仍需对热带雨林动态变化和蝙蝠物种多样性协同响应进行长期的监测[31],准确评估森林破碎化程度对蝙蝠物种多样性的影响,了解蝙蝠生活史特征和区域性关键生态因子的重要作用,准确定位蝙蝠受位危害的现状,制定优先保护对象,充分考虑基础设施合理规划对蝙蝠物种保护的重要性,从而维持蝙蝠物种多样性,提高生态系统服务功能。
猜你喜欢
小哥白尼(野生动物)(2022年7期)2022-09-20
女报(2020年7期)2020-08-17
小天使·四年级语数英综合(2019年4期)2019-10-08
儿童故事画报(2019年8期)2019-08-14
艺术启蒙(2018年8期)2018-08-22
小溪流(画刊)(2016年12期)2017-02-04
好孩子画报(2016年12期)2017-01-12
小火炬·智漫悦读(2015年4期)2015-06-05
微型小说选刊(2015年5期)2015-06-05
小学生·多元智能大王(2014年5期)2014-07-24
