
Effects of climate,biotic factors,and phylogeny on allometric relationships:testing the metabolic scaling theory in plantations and natural forests across China
2021-01-11 07:08HanSunXiangpingWangandDayongFan
Forest Ecosystems 2020年4期
Han Sun,Xiangping Wang* and Dayong Fan
Abstract
Keywords: Allometry, Successional stage, Climate, Plantation, Conifer, Broadleaf trees, Metabolic scaling theory
Background
Allometry relationships are not only required in estimating ecosystem carbon sources and sinks (Chave et al.2005, 2014) and in up-scaling ecological processes from the individual to the ecosystem level (Enquist and Niklas 2001; Brown et al. 2004), but also important for understanding the basic biophysical and evolutionary control of plant organs (West et al. 1999; Apol et al. 2008). Considering its critical roles, allometry has long been a focus of studies in ecology and forestry. To date, several biophysical models on allometry have been proposed (e.g.geometric, elastic, and stress similarity (Rubner 1883;McMahon and Kronauer 1976)). Among these models,metabolic scaling theory (MST) provides a series of testable predictions regarding allometric scaling relationships, based on basic biophysical constraints from the organ, individual to the ecosystem level (West et al.1997; Brown et al. 2004). Because of its broad theoretical importance, MST has attracted far more attention than other models (Muller-Landau et al. 2006; Price et al.2009; Poorter et al. 2012; Duncanson et al. 2015).
Nevertheless, MST has been debated since it was proposed, partly because the model has not explicitly taken into account the influence of environmental gradient and different among taxa. In fact, studies have shown that allometries may deviate from MST predictions and can vary as a function of abiotic and biotic factors(Wang et al. 2006; White et al. 2007; Lines et al. 2012;Hulshof et al. 2015). For instance, the height-diameter allometric scaling exponent may decrease with lower temperature so as to avoid xylem cavitation under cold and drought conditions (Wang et al. 2006; Lines et al.2012). The inclusion of climate and forest type can,therefore, markedly improve the explanatory power of allometric models of biomass allocations (Wang et al.2008). Biomass distribution scaling exponents may have significant differences in their goodness of fit results before and after species specific characteristics are considered (Wang et al. 2011), suggesting that species identity is also important in affecting scaling relationships. Although these criticisms may not be enough to reject MST in its entirety, it is clear that MST needs to be improved.
Recently, studies have proposed an explanation for why observed scaling exponents deviated from the MST predictions in forests (Rüger and Condit 2012; Duncanson et al. 2015; Sun et al. 2017a). These authors found that measured allometric exponents only agreed with MST predictions in late successional forests, because earlier successional forests were more affected by biotic and abiotic factors. For instance, Duncanson et al.(2015) demonstrated that allometries of trees in younger forests may vary quite substantially, while tree allometries in mature forests taller than 35 m were consistent with MST predictions. Likewise, Sun et al. (2017a) found that H-D allometric scaling exponents conformed to MST predictions in late-successional broadleaf and Korean pine mixed forests in northeast China, while forests in earlier successional stages revealed greater deviations.These deviations from the predictions of MST may be primarily caused by biotic factors such as light competition and recruitment limitation, while climate may indirectly affect allometric exponents through these biotic factors. However, this hypothesis was only derived from studies in temperate forests. More tests need to be conducted to determine whether it applies to other forest biomes (e.g. subtropical and warm-temperate forests),and is therefore applicable across a broad climatic scale(Duncanson et al. 2015;Sun et al. 2017a).
Previous studies have mainly focused on the allometric relationships in natural forests (Muller-Landau et al.2006; Price et al. 2009; Poorter et al. 2012). However,plantations are significantly different from natural forests in species composition and forest structure due to forest management (Luo et al. 2012). On the one hand, plantations are mostly dominated by fast-growing tree species,which is similar to early successional natural forests. On the other hand, forest management in plantations can increase the utilization of environmental resources(Binkley et al. 2004) which makes the living conditions steadier, and thus the allometries may be more similar to late successional natural forests (Duncanson et al.2015). Therefore, the allometric relationships in plantations, whose species identity and age are more comparable to early successional forests, may better fit MST predictions like in mature forests than in early successional forests. One aim of this study is to test this hypothesis.
Allometry is not only important for ecological theory,but also an important method used for carbon storage and flux estimation. One of the most widely-used methods in estimating biomass is based on the allometric relationships between biomass and diameter at breast height (Fang et al. 2006; Wang et al. 2013). In this study,we assembled data on biomass-diameter allometric relationships from the literatures on Chinese natural forests and plantations, in order to compare the scaling exponents of three important allometric relationships: leaf biomass-diameter (L-D), stem biomass-diameter (S-D),and root biomass-diameter (R-D). These allometric relationships are not only critical for forest biomass estimation, but are also important indicators of individual biomass allocation strategies for plants (Luo et al. 2015).Previous studies have shown that the leaf mass fraction increased with increased light competition and nutrients(Poorter et al. 2012). The stem biomass fraction increased significantly with stem density (Poorter et al.2012), and root biomass was found to be much less limited by growing-season precipitation compared to shoot biomass (Wang et al. 2008). These different mechanisms may also affect L-D, S-D, and R-D allometric relationships, leading to variability and deviations from MST predictions. So, as a part of this study, we examined the differences among the three allometric relationships.
Here we test three hypotheses: 1) the allometric relationships of trees support the predictions of MST on a broad scale; 2) the observed deviations from MST predictions are caused by climate, biotic factors, and/or phylogeny; 3) abiotic and biotic factors influence allometric relationships in plantations and natural forests differently, and different allometric relationships (i.e. LD, S-D, and R-D) are affected differently.
Methods
Data collection
We assembled scaling exponents from studies that reported the allometric relationships between leaf (L),stem (S) and root (R) biomass and tree diameter at breast height (D) across China. The literatures used different functions to fit the relationship between biomass and D, such as power, logarithm, linear, and quadratic functions, etc. However, only the power function is consistent with theoretic allometric models and can be used to test MST (West et al. 1997, 1999; Brown et al. 2004).Consequently, we assembled data only from literatures that used the power function (Biomass=aDb, where b is the scaling exponent). For each record, we also documented the following information whenever available:(1) geographic coordinates, including latitude and longitude of the plantation or forest; (2) forest type, forest origin, species composition, and dominant tree species;and (3) forest structure variables, including the stem density and tree height ranges of each plot measured at the exact time when these organ biomass-diameter power functions exponents were estimated. As a result,our dataset of 284 records included 201 L-D scaling exponents, 259 S-D scaling exponents, and 189 R-D scaling exponents from 143 plots spanning a latitudinal range of 18.7°-52.0° N, and a longitudinal range of 87.2°-130.9° E(Fig. 1). These plots included all the major forest biomes in China. We grouped the plots into two forest origins:natural forest and plantation. We also grouped natural forest plots into two successional stages: secondary natural forest and primary natural forest. Many literatures have described the successional stages of their plots.When this information is not explicitly described,the successional stage of some plots can be well judged by species composition and forest age, stock volume, etc., as described by the literatures. In a few cases, there was no enough information to determine whether a plot is a secondary or primary forest. These data were excluded from our dataset(see Appendix 1 and 2 for more detail).
Based on the latitude and longitude of each plot, we extracted the monthly average temperature and precipitation data (means from 1950 to 2000) for each plot from a very high resolution (1 km×1 km) global climate database using ArcGIS 10.2 (Hijmans et al. 2005). Climatic indices for each plot were calculated using these monthly climate data, including annual potential evapotranspiration (PET) as a measure for energy availability,mean temperature for the coldest month (MTCM) as a proxy for winter coldness, and moisture index (MI, calculated as PET - mean annual precipitation) which is increasingly recognized as a better metric of water availability than commonly-used mean annual precipitation (Fang et al. 2012).
Statistical analysis
To test MST’s predictions on the L-D, S-D, and R-D scaling exponents (= 2, 8/3, and 8/3, respectively), we pooled the data together and compared the means and 95% confidence intervals of the three scaling exponents to MST predictions. To test whether allometric relationships changed with successional stage, leaf phenology and leaf forms, we grouped data into three forest types(secondary, primary, or planted forest), two leaf phenology (evergreen vs. deciduous), and two leaf forms(conifer vs. broadleaf) and compared the three scaling exponents in each group with MST predictions.
To determine the effects of climate, biotic factors, and phylogeny on allometric relationships, we explained the L-D, S-D, and R-D scaling exponents with variables as follows: (1) Climate. Previous studies have shown that energy and water availability, and winter coldness can affect allometric relationships (Wang et al. 2006; Lines et al. 2012; Sun et al. 2017a), so we selected PET (energy), MTCM (coldness), and MI (water availability) for statistical analyses. Other climate indices are similar to PET, MTCM, and MI in biological meaning and thus were not used to avoid collinearity. (2) Biotic factors.Studies have shown that competition, species (functional) identity, and forest succession stage can affect allometric exponents, thus we choose stem density, forest age, biome (boreal, temperate, warm-temperate, or subtropical forest), and forest type (primary forest, secondary forest, and plantation) as biotic factors to examine their potential effects on the three scaling exponents.Stem density and forest age were log-transformed to increase normality and homoscedasticity of residuals,which is a common practice in statistical analysis (Quinn and Keough 2002; Crawley 2007; He et al. 2009; Wang et al. 2009). (3) Phylogeny. To test for a phylogenetic effect, we used family, genus, and species as explanatory terms, which have been shown to be good surrogates for phylogenetic relationships among taxa (He et al. 2009).This is because some species cannot be found in the existing phylogenetic database (Zanne et al. 2014), and thus we were not able to construct a phylogenetic tree for the species in our study.
We used bivariate analyses and multivariate mixedeffect models to explain the L-D, S-D, and R-D allometric exponents with above-mentioned abiotic and biotic factors. In mixed models, we used genus as a random effect for the fixed effect of species, while family was used as random effect for that of genus (for details, see e.g.He et al. 2009). Records missing forest age or stem density were excluded from the mixed-model analysis, and thus 125 records including 123 L-D scaling exponents,125 S-D scaling exponents, and 81 R-D scaling exponents were retained in this analysis. To identify the major factors affecting the three allometric relationship,we refined our model using the Akaike Information Criterion (AIC) to drop unnecessary variables (Johnson and Omland 2004). All statistical analyses were performed with R 3.5.1 (R Core Team 2014).
Results
When data were pooled together, the mean L-D scaling exponent was 2.01 (95% CI: 1.908-2.212), which agrees with the MST prediction of 2. The mean of the S-D and R-D scaling exponent was 2.303 (95% CI: 2.253-2.353)and 2.194 (95% CI: 2.125-2.263), respectively, both were lower than the MST predictions of 8/3 (Fig. 2).
When examined for each forest type separately(Fig.3),the L-D scaling exponent again agreed with the MST prediction in all the three forest types: secondary and primary natural forests, and plantations. However, both S-D and R-D scaling exponents showed greater deviation from MST predictions in secondary forests, and were close to MST predictions in primary forests, while plantations revealed medium exponents between secondary and primary forests. The L-D scaling exponents also agreed with the MST prediction irrespective of leaf phenology or leaf forms (Fig. 4). However, the S-D and R-D scaling exponents were lower than MST predictions when examined in each category grouped by leaf phenology or leaf forms.
Some climate, biotic factors, and phylogeny were significantly related with scaling exponents(Table 1,Figs.5,6 and Table A1). Although some r2were not very strong, significant relationships could still reflect the trends. In fact, low r2but significant relationships were often found in previous studies of allometric scaling factors at large scale (Luo et al. 2012). In climate factors,the temperature indices (PET and MTCM) were significantly and negatively correlated with the L-D scaling exponent (r2=0.02-0.03, p<0.05), while the moisture index (MI) had a significant negative relationship with the S-D scaling exponent (r2=0.02, p<0.05). As for biotic factors, stem density was significantly related with the L-D scaling exponent (r2=0.05, p<0.01), while forest type was significant in explaining the S-D scaling exponent (r2=0.03, p<0.05), and leaf form had a significant effect on the R-D scaling exponent (r2=0.02,p<0.05). In comparison, phylogeny (family, genus and species) revealed much stronger effects on the S-D and R-D scaling exponents (r2=0.25-0.58, p<0.05).
There were clear differences in the main correlates of allometric relationships between plantations and secondary forests. Scaling exponents were related to only a few factors in plantations: stem density was positively correlated with the L-D scaling exponent (r2=0.05, p<0.05),genus was significant in explaining the S-D scaling exponent (r2=0.28, p<0.05), while MTCM was negatively correlated with the R-D scaling exponent (r2=0.05, p<0.05). However, scaling exponents in the natural forests were affected by more factors: the L-D scaling exponent was affected by climate (PET and MTCM, r2=0.09-0.10,p<0.01) and biotic factors (leaf phenology, r2=0.07, p<0.05);and the S-D scaling exponent was affected by forest type (r2=0.07, p<0.01) which means that successional stage had a significant effect on scaling exponents in a natural forests.Both S-D and R-D scaling exponents were significant affected by leaf form (r2=0.06-0.08, p<0.05)and phylogeny(r2=0.45-0.82,p<0.01).

Table 1 The r2 of each climate,biotic factors and phylogeny in explaining the scaling exponent for allometric relationships of leaf,stem and root biomass with diameter at breast height(L-D, S-D,and R-D, respectively). Negative r2 represents negative correlation while others represent positive correlation. Significant relationships (p<0.05)were boldfaced. Abbreviations:PET annual potential evapotranspiration,MTCM mean temperature for the coldest month, MI moisture index

Table 1 The r2 of each climate,biotic factors and phylogeny in explaining the scaling exponent for allometric relationships of leaf,stem and root biomass with diameter at breast height(L-D, S-D,and R-D, respectively). Negative r2 represents negative correlation while others represent positive correlation. Significant relationships (p<0.05)were boldfaced. Abbreviations:PET annual potential evapotranspiration,MTCM mean temperature for the coldest month, MI moisture index (Continued)
The aforementioned results suggested that climate, biotic factors, and phylogeny all can have significant influence on allometric exponents. Indeed, many of these variables were retained in the final mixed-models and had significant explanatory power (Table 2). However,the result of mixed-models suggested that the major factors influencing the three allometric relationships were different. L-D scaling exponent was only significantly affected by MTCM, which explained 3.24% of variation(p<0.05). On the other hand, S-D and R-D scaling exponents were affected by many factors including climactic, biotic, and phylogenetic factors. The S-D scaling exponent was significantly affected by MTCM (i.e. climate index, %SS=2.76, p<0.05), forest types and stem density (i.e. biotic factors, %SS=7.32-5.86, p<0.01), and family (i.e. phylogeny, %SS=26.43, p<0.05). The R-D scaling exponent was significantly affected by forest types, stem density (%SS=8.20-8.54, p<0.01) and family (%SS=36.16, p<0.05).
Discussion
Allometries vary with successional stage in natural forests
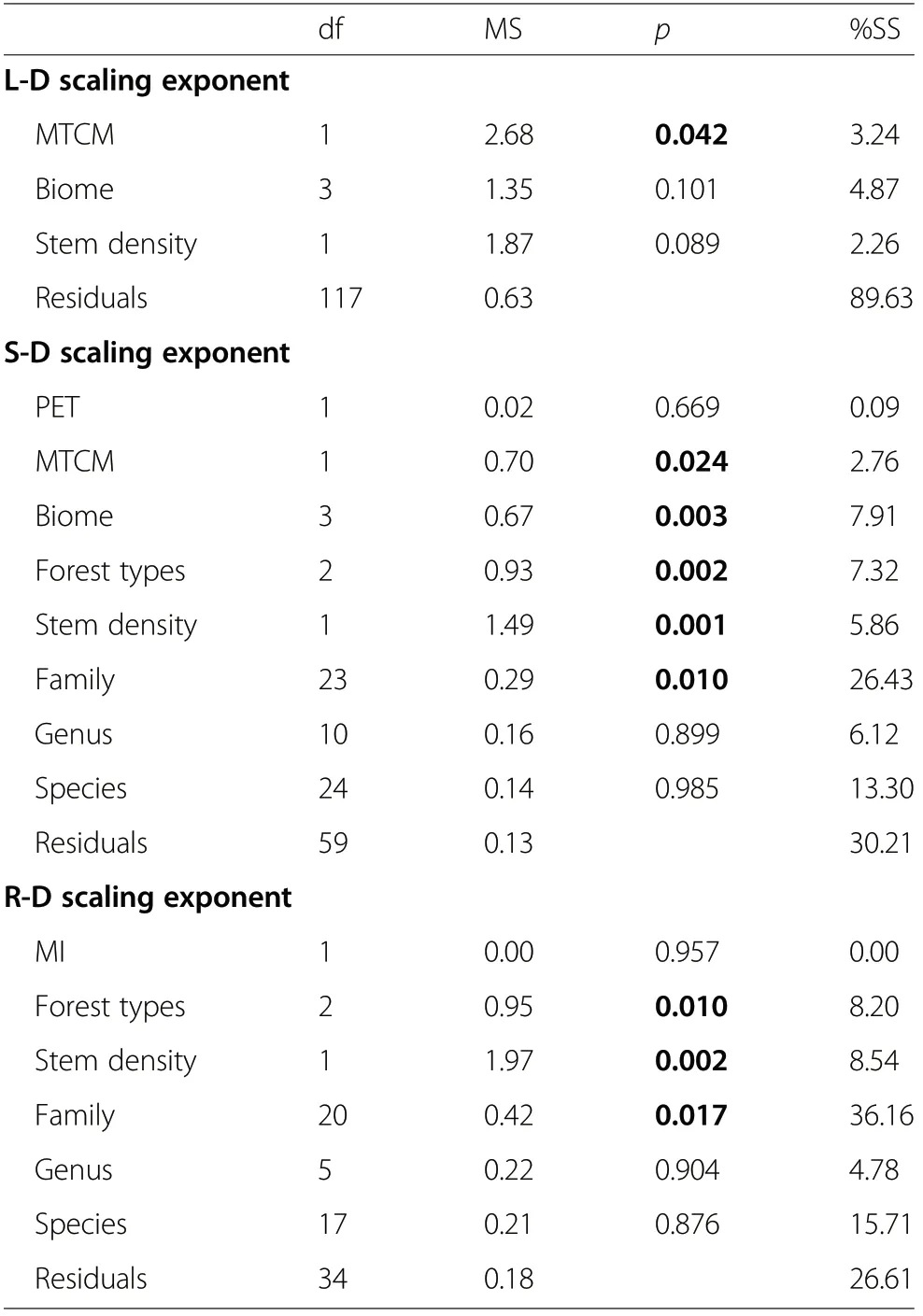
Table 2 Summary of mixed-models for the effects of climate(PET,MTCM, MI,and biome),leaf form (conifer or broadleaf), leaf phenology(evergreen or deciduous), forest types (secondary natural forest,primary natural forest,and plantation), stem density,forest age, and phylogeny groups(family, genus, and species) on L-D, S-D,and R-D allometric relationships. Bold indicates p<0.05. Unnecessary variables were dropped based on AIC. %SS,percentage of sum squares explained; df degree of freedom, MS mean square
Although according to our results, allometries in primary forests did not fully support MST predictions,which seems inconsistent with MST, it still cannot be rejected. We found that the data were not well estimated by reduced major-axis (RMA) regressions, and that when using an ordinary least squares (OLS) regression for fitting the slope (i.e. scaling exponent), the allometry exponent values were underestimated (note that OLS regression is recommended when predicting Y from X while RMA is recommended when test if the slope equals a specific value for the line best describing the relationship between Y and X (Warton et al. 2006;Price et al. 2007)). The scaling exponents of allometric relationships in primary forest would therefore be affected leading to lower statistical results. Therefore,the allometric relationships in primary forest might better support the predictions of MST if the underestimates of traditional regressions are accounted for. Thus, our results were consistent with the predictions of MST.
Differences in allometries between plantations and natural forests
Our results showed that forest origin influenced allometries. Scaling exponents deviated more in secondary natural forests and were close to theoretical predictions in primary natural forests, while plantations fell somewhere in the middle. These are consistent with the results of Wang et al. (2008), which demonstrated that root-shoot allocation scaling exponents in plantations were in between those of primary and secondary forests. However,both of our results contrast with Mokany et al. (2006)who reported no significant difference in the ratios of root to shoot biomass between plantations and natural forests.It may be because that Mokany et al. (2006) did not analyze natural forests at different stages of succession separately. Our results suggest that, if succession stages are ignored and data from early and late successional forests are pooled together, the result may be close to the intermediate value that is observed in plantations.The importance of distinguishing between successional stages when examining allometric relationships in natural forests(see also Duncanson et al. 2015; Sun et al. 2017a), as well as comparing allometries between natural forests and plantations,is clearly illustrated by our findings.
Plantations are generally established in more fertile and moister soil than natural forests, and comprehensive forest management practices can further ensure them more appropriate living conditions, compared with other types of forests. Because of sufficient soil moisture and nutrient availability in plantations, trees do not need to increase their allocation to root biomass to get more nutrients and water (Mokany et al. 2006; Wang et al.2008). Consequently, plantations generally have lower root to shoot biomass ratios than natural forests (Luo et al. 2012). Although Luo et al. (2012) did not look into age but used stands over 20 years and did not analyze the age effect, our results showed that the effect of forest age is weak. Furthermore, forest management practices in plantations such as thinning and selected harvesting affect the biotic factors that are important for tree allometries, which lead to different allometries between plantations and natural forests (Luo et al. 2012, 2014;Zhang et al. 2015). For example, thinning reduces stem density, which reduces the competition for light (Rüger and Condit 2012;Sun et al.2017a),and thus promotes the accumulation of biomass in woody tissues (Poorter et al.2012), while pruning reduces the branches and leaves of the trees which affects the accumulation of leaf biomass(but not necessarily the most active part of the tree foliage(“unproductive leaf mass”) and can also affect the diameter development if in extreme conditions.
Additionally, our result showed that phylogeny have few significant effects on allometries in plantations. This may be due to management practices involving species selection. Such species selection may make biodiversity in plantations lower than natural forests (Brockerhoff et al. 2008). This may be the reason that phylogeny did not have a significant effect in plantations.Consequently,forest management practices may lead to different patterns of allometries in plantations than those in natural forests, and it must be considered when conducting research on allometric relationships in plantations.
More importantly, the differences between allometric relationships in plantations, secondary and primary natural forests mean that the estimation of stand biomass requires the use of relevant allometric relationships measured from the same forest type. But this has not been paid attention to in biomass estimation yet, which may lead to errors in biomass estimation.
Factors determining allometric variation
In our multivariate models, MTCM, biome, forest type,stem density, and species significantly affected allometric exponents. These results, examining abiotic and biotic effects on allometries, are consistent with previous broad-scale studies (Duncanson et al. 2015).
Authors have found that climate can have a weak indirect effect on allometric relationships (Wang et al.2006;Lines et al.2012).In our study, winter temperature had a significant effect on L-D and S-D scaling exponents. Trees may adopt particular allometric relationships to survive cold temperatures in winter (Lines et al.2012). Thus, winter coldness may become an important constraint for allometric relationships. Our results are in line with previous studies showing that higher stem density intensifies light competition between trees(Getzin et al. 2006; Wang et al. 2016). Competition for light has been known to explain the highest proportion of the variations in the D-H allometric exponent(Poorter et al. 2012). Our study further confirmed that the S-D and R-D scaling exponents were linked to light competition too. However, we also found that these effects were weak, which means that biophysical constraints still strongly influence allometric relationships.This may explain why the effect of external factors was almost insignificant over a small gradient in some studies (Poorter et al. 2012; Sun et al. 2017b).
Meanwhile, among these factors, phylogeny explained the highest proportion of the variations in the S-D and R-D scaling exponents (Table 2, 26.43% and 36.16% of variation). Wang et al. (2011) found that there were significant deviations in scaling exponents when interspecies differences were considered with the stratified Bayesian method. Our results support their findings. A possible mechanism that phylogeny affects the S-D and R-D scaling exponents is that phylogeny has been shown to be associated with conduit size, i.e., recently evolved species have longer and wider conduits than ancient ones (Savage et al. 2010; Jacobsen et al. 2012), while the size of conduit largely determines the water transport efficiency it can consequently leads to the variations of the S-D and R-D scaling exponents. Therefore, theoretical models such as MST should be extended to include the effect of phylogeny.
信息与计算科学专业是数学、信息科学和计算机科学三者交叉的学科,它以数学为基础,计算机为工具,解决信息和工程计算方面的实际问题.这一专业设置较好地适应了以信息技术为核心的全球经济发展格局下的人才培养与专业发展[1-4].
Besides, we noted that the influence of phylogeny is mainly reflected in family level (Table 2). This may imply that the allometric relationships are evolutionary conservative at the family level. Previous studies have generally found that conifers had a higher D-H scaling exponent than broadleaf trees because of differences in wood density or branching architecture (Lines et al.2012; Duncanson et al. 2015; Hulshof et al. 2015). Our results are consistent with these findings.
Different variation rules among L-D, S-D and R-D allometries
Previous studies have shown that the biomass allocation and functional traits of leaves, stems, and roots were affected by various abiotic, biotic and evolutionary factors(Wang et al. 2008; Poorter et al. 2015; Wright et al.2005, 2017). According to MST, different allometric relationships between traits are connected and can be therefore be deduced from each other, however, our results showed that variations, and the major factors producing those variations, were significantly different for L-D, S-D and R-D relationships. This is consistent with many previous studies (Deng et al. 2006; Poorter et al.2012). For example, previous studies found that leaf biomass allocation was relatively insensitive, while stem and root biomasses were affected by many factors (Poorter et al. 2012). Deng et al. (2006) showed that during water shortage, between above-ground biomass and stem density of shrub communities varied significantly with water gradient, while the scaling exponent of total biomass with underground biomass was not only unaffected by moisture gradient but was close to the MST prediction of −4/3. Our results also showed that in forests, the scaling exponent of the L-D relationship supported MST prediction and had no significant deviations in plantations or secondary forests, while variations observed in the S-D and R-D scaling exponents were relatively large.The L-D scaling exponent was more stable and only weakly affected by MTCM, while the S-D and R-D scaling exponents had similar patterns of variation and were significantly affected by climate, succession, light competition, and phylogeny. Our results and Deng et al. (2006)both showed different responses for different allometric relationships to various factors. Consequently, it is necessary to study multiple allometric relationships simultaneously in order to understand how they are related and how they are influenced by various abiotic and biotic factors.
Our results indicated that L-D exponent is relatively stable,and conform to MST prediction irrespective of successional stage, forest origin, phylogeny, leaf form and phonology. To our knowledge, this difference from S-D and R-D allometries has not been reported before in large scale studies. We propose three potential explanations as follows. First, as the photosynthesizing organ, leaves may have stronger biophysical constraints and thus may be more phylogenetically conserved than the stem or root.Previous studies found that leaves prefer to change shapes and area to adapt to the environment rather than change biomass allocation (Poorter et al. 2012). Our results also showed that phylogeny had no significant effect on the LD scaling exponent,so our results all support this explanation. Second, trees can shed and regenerate leaves to adapt leaf biomass to their environment (Zanne et al.2014). This mechanism may also make leaf biomass allocation more stable and closer to theoretical predictions as they maintain optimal metabolism. Third, adding secondary xylem and gradually lost water transport function of earlier developed xylems in the stem and root during growth may change stem and root biomass fractions(Niklas and Spatz 2004). This change may cause further variation in the S-D and R-D scaling exponents. Since leaves have no secondary xylem,the L-D scaling exponent would not be altered by the same mechanism.However,which of these explanations is the major cause of the differences among L-D, S-D, and R-D scaling exponents remains unclear and needs further examination.
Conclusions
In this study, we examined effects of climate, biotic factors, and phylogeny on the allometric relationships of leaf, stem, and root biomass to diameter in plantations and natural forests across China. Our research at a broad scale, including biomes from boreal to tropical forests across China, thus provided strong support to the idea that allometric relationships are also vary with successional stage. Scaling exponents are consistent with MST predictions in primary forests and deviated greater from theoretical predictions in secondary forests. The allometric scaling exponents of plantations were in between secondary and primary natural forests even though the species composition and forest age of a plantation is closer to that of secondary forests, and were affected by fewer factors than in natural forest. These results reflect the influence of forest management practices on allometries. MST cannot explain this variation yet, however, since there is a great amount of plantation area in the world, it is important to do more research on allometries in closely managed plantations. Our study also found that the L-D scaling exponent was relatively insensitive, while the S-D and R-D scaling exponents were significantly affected by climate, biotic factors, and phylogeny. This suggested that L-D allometry is more strongly controlled by biophysical constraints compared with S-D and R-D allometry, however, the mechanisms behind this difference still needs further examination.
Supplementary information
Supplementary informationaccompanies this paper at https://doi.org/10.1186/s40663-020-00263-y.
Additional file 1.
Additional file 2.
Abbreviations
MST: metabolic scaling theory; H: height; D: diameter; L: leaf biomass;S: stem biomass; R: root biomass; PET: annual potential evapotranspiration;
MTCM: mean temperature for the coldest month; MI: moisture index;
AIC: Akaike Information Criterion; CI: confidence intervals; RMA regression: reduced major-axis regression; OLS regression: ordinary least squares regression
Authors’contributions
Han Sun, Xiangping Wang and Dayong Fan conceived the study; Han Sun performed the analyses and wrote the first draft; all authors contributed critically to successive drafts and gave final approval for publication.
Funding
This study was supported by the Fundamental Research Funds for the Central Universities (BLX201802), National Natural Science Foundation of China (31870430), and the National Key Research and Development Program of China(#2017YFC0503901).
Availability of data and materials
The datasets used and/or analyzed in during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
No applicable.
Consent for publication
No applicable.
Competing interests
The authors declare that they have no competing interests.
Received: 7 December 2019 Accepted: 2 July 2020
猜你喜欢
数学杂志(2022年3期)2022-11-27
山西大同大学学报(自然科学版)(2022年1期)2022-03-17
青年生活(2019年7期)2019-09-10
环球时报(2019-07-16)2019-07-16
速读·下旬(2017年7期)2017-08-03
东方教育(2017年7期)2017-07-03
人间(2016年28期)2016-11-10
现代经济信息(2016年24期)2016-11-09
中国信息技术教育(2015年18期)2015-11-03
中国信息技术教育(2015年11期)2015-07-20

- Forest Ecosystems的其它文章
- Evaluating the impact of sampling schemes on leaf area index measurements from digital hemispherical photography in Larix principis-rupprechtii forest plots
- Discovering forest height changes based on spaceborne lidar data of ICESat-1 in 2005 and ICESat-2 in 2019:a case study in the Beijing-Tianjin-Hebei region of China
- Comparison of the local pivotal method and systematic sampling for national forest inventories
- Effects of firewood harvesting intensity on biodiversity and ecosystem services in shrublands of northern Patagonia
- The Siberian moth(Dendrolimus sibiricus),a pest risk assessment for Norway
- Hydrological functioning of forested catchments,Central Himalayan Region,India