
放牧强度对典型草原6个物种生物量分配格局的影响
2021-01-09 06:44:28张紫薇林长存刘佳乐苏德荣平晓燕
草地学报 2021年1期
张紫薇,高 斌,林长存,王 青,刘佳乐,杨 娜,苏德荣,平晓燕
(北京林业大学草业与草原学院,草地资源与生态研究中心,北京 100083)
放牧是天然草原最重要的利用方式[1]。放牧作为草地生态系统的重要干扰因子和驱动力,会通过家畜的选择性采食、踩踏和排泄物归还等方式直接或间接的影响草地植物的群落演替进程[2]、群落组成和生产力[3]及草地生态系统的资源分配格局[4]。但过度放牧等不合理的人类活动会导致草地退化,降低草地生态系统的生产力和土壤养分含量,威胁我国草原区的生态安全,也给边疆稳定和社会经济发展带来不利影响[5]。深入理解草原的放牧退化过程和阐明草原退化过程中的物种形成机制,对恢复和管理放牧退化草原具有十分重要的意义[3,6]。而物种在放牧退化过程中的生态适应对策是理解放牧退化草原群落物种组成的关键[7]。
生物量分配格局反映植物应对环境和资源胁迫时的光合产物分配策略,被认为是草本植物适应放牧的一种重要的生态适应对策[8-9]。生物量分配或光合产物分配格局在个体水平上作为影响草本植物活力和耐牧性的重要功能性状[10],在群落水平上会影响群落的结构、功能和稳定性[11],在生态系统或区域水平是全球陆地生态系统碳循环模型中的一个重要参数[12]。草地植物的生物量分配格局受气候、地形、食草动物采食、刈割、资源有效性、种内或种间竞争等诸多因素的共同影响[13-14]。有关植物生物量或光合产物分配策略的机理性假说包括功能平衡假说(Functional Balance Hypothesis)、源汇关系假说(Source-sink Relationship Hypothesis)和相关生长关系假说(Allometric Relationship Hypothesis)等[15]。但这些假说大都针对环境变化对光合产物分配格局的影响或用以阐明光合产物的分配机理,很难解释放牧影响下植物的光合产物分配策略。
典型草原是我国温带草原中具有代表性和典型性的一种草原类型,也是欧亚草原区的典型代表,约占全国草地总面积的10.25%[16]。前期已有较多关于放牧强度对个体水平的生物量分配格局[10,17-18]、群落水平的生物量分配格局[19-20]、群落或生态系统水平的光合产物分配格局[3,21]以及个体尺度的繁殖分配格局等的影响研究[22],但大部分研究集中在群落或生态系统水平[11,14,21],对个体尺度的生物量分配格局研究也大都只关注单一物种,较少针对占据群落不同地位的物种开展生物量分配格局的研究,从而限制了我们对放牧影响下物种的生态适应策略和生物量分配格局在群落物种组成中具体作用和贡献的准确理解。因此本研究选取河北坝上地区的典型草原为研究区域,从该区域不同放牧强度样地的群落优势种、亚优势种和常见种中选择6个物种为研究对象,通过分析6个物种在不同放牧强度下的生物量分配格局,从个体水平阐明不同物种应对放牧干扰的生态适应对策,为深入理解放牧影响下该区域的物种组成机制提供理论基础和参考。
1 材料与方法
1.1 研究区概况
研究区位于河北省沽源草地生态系统国家级野外观测实验站(116°14′ E,41°37′ N,海拔1 430 m),属半干旱大陆季风气候带,年降水量350~450 mm,且多集中在6—9月,占全年降水量的79%,年蒸发量1 700~2 300 mm,年平均气温1℃左右,最冷月(1月)平均气温—18.6℃,最热月(7月)平均气温17.6℃,≥0℃年积温 2 000~2 800℃,地带性土壤类型为粟钙土,地带性植被为典型草原。研究区维管束植物354 种,分属于61 个科,196 属。其中种数最多的科是菊科(Composite)、禾本科(Gramineae)、蔷薇科(Rosaceae)、豆科(Leguminosae)和藜科(Chenopodiaceae),这 5科约占总种数的 50%以上。该区草原植被以羊草(Leymuschinense)为建群种,克氏针茅(Stipakrylovii)为次优势种,冷蒿(Artemisiafrigida)、糙隐子草(Cleistogenessqurrosa)等为常见种。
1.2 野外试验设计与方法
1.2.1野外试验设计 研究区自2009 年起连续开展了不同放牧强度(围封、轻度、中度和重度放牧)影响下典型草原生态学过程的控制试验研究。本研究基于前期的野外控制实验样地,设置了3个放牧强度:轻度放牧(1.33 羊·hm-2)、中度放牧(4.0 羊·hm-2)和重度放牧(6.67 羊·hm-2),以围封样地为对照。每个放牧强度 3次重复,共计 12 个小区,每个小区面积 1.5 hm2,所有放牧小区均为生长季(5—10月)持续放牧[23]。
1.2.2野外群落调查 在野外试验地点划分的围封区、轻度放牧区、中度放牧区和重度放牧区分别进行群落样方调查,不同放牧强度样地的基本特征见表1。其中,围封样地的群落优势种为羊草,亚优势种为线叶蒿(A.subulate);轻度放牧样地的群落优势种为羊草,亚优势种为糙隐子草和华北米蒿(A.giraldii);中度放牧样地的群落优势种为羊草,亚优势种为冷蒿和二裂委陵菜(Potentillabifurca);重度放牧样地的群落优势种为羊草,亚优势种为马蔺(Irislactea)。
1.2.3样品取样与分析方法 2016年在草本植物生长最旺盛的7月分别在围封区、轻度放牧区、中度放牧区和重度放牧区等4个不同放牧处理样地采集了马蔺、线叶蒿、糙隐子草、二裂委陵菜、华北米蒿和羊草等6种植物的单株样品,在每个放牧梯度样地,每个物种各随机选取长势均一的5株植株将其整株挖取,在挖取根系时尽量小心将全部根系挖出。在采样过程中,保证取样的随机性和植株的完整性。将所有样品带回实验室后,将植株分为地上和根系两部分,其中根系部分放入土壤筛中清洗去除杂质和土壤后,将地上和根系样品分别放入65℃烘箱中烘干48 h至恒重后用万分之一天平称量获得干重,每个处理5次重复。

表1 不同放牧强度样地的基本特征Table 1 Characteristics of grazing plots under different grazing intensities
1.3 数据处理和分析
利用R语言程序软件(3.6.1 for Windows)对不同放牧强度处理下6个物种的单株地上生物量、单株地下生物量、单株总生物量和根冠比等指标进行了单因素方差分析,同时对同一放牧强度下不同物种之间的根冠比进行了单因素方差分析,对方差分析结果显著的数据进行LSD多重比较。为深入探讨典型草原物种的生物量分配格局与群落结构或功能的关系,本研究基于Pearson相关分析对6个物种的根冠比和物种在群落中的重要值进行了相关分析。统计结果的显著性水平为P<0.05。利用Sigmaplot 12.0软件作图。
为了分析物种在群落中的地位和作用,根据群落调查结果,计算6个物种的重要值(Important value,IV(%)),计算公式如下:
IV=(相对盖度+相对密度+
相对频度+相对生物量+相对株高)/5
(1)
2 结果与分析
2.1 放牧强度对6个物种单株生物量的影响
由图1可知,放牧强度对6个物种的单株地上生物量均产生极显著影响(P<0.001),且放牧强度对不同物种的单株地上生物量的影响趋势基本相同。除线叶蒿和羊草外,马蔺、糙隐子草、二裂委陵菜、华北米蒿等物种的单株地上生物量在轻度、中度和重度放牧处理下均显著低于对照。马蔺的单株地上生物量随放牧梯度呈现显著下降趋势,在四个放牧处理下均呈显著差异。除糙隐子草外,与对照和轻度放牧相比,重度放牧显著降低了其他物种的单株地上生物量。
不同物种单株地下生物量对放牧强度的响应各不相同(图2,马蔺(IL)P=0.08;线叶蒿(AS)P>0.05;糙隐子草(CS)P=0.01;二裂委陵菜(PB)P<0.001;华北米蒿(AG)P=0.05;羊草(LC)P>0.05)。放牧强度极显著地影响了糙隐子草和二裂委陵菜的单株地下生物量。与单株地上生物量的影响相反,与对照相比,重度放牧显著提高了糙隐子草的单株地下生物量。相比对照和轻度放牧,中度和重度放牧显著增加了二裂委陵菜的单株地下生物量。

图1 放牧强度对典型草原6个物种单株地上生物量的影响Fig.1 Effects of grazing intensity on aboveground biomass of individual plant of six plant species in a typical grassland注:IL:马蔺;AS:线叶蒿;CS:糙隐子草;PB:二裂委陵菜;AG:华北米蒿;LC:羊草,图中不同小写字母表示不同放牧强度间差异显著(P<0.05),误差棒表示标准差。CK:围封对照;LG:轻度放牧;MG:中度放牧;HG:重度放牧,下同Note:IL:Iris lactea;AS:Artemisia subulate;CS:Cleistogenes squarrosa;PB:Potentilla bi furca;AG:A. giraldii;LC:Leymus chinensis. Different letters indicate significant difference at the 0.05 level,and the error bar of each treatment represents the standard deviation of five replications. CK:control;LG:light grazing;MG:moderate grazing;HG:heavy grazing,the same as below
放牧强度对6个物种的单株总生物量均产生显著或极显著影响(图3,马蔺(IL)P<0.001;线叶蒿(AS)P<0.01;糙隐子草(CS)P<0.01;二裂委陵菜

图2 放牧强度对典型草原6个物种单株地下生物量的影响Fig.2 Effects of grazing intensity on belowground biomass of individual plant of six plant species in a typical grassland
(PB)P<0.001;华北米蒿(AG)P<0.001;羊草(LC)P<0.05)。其中,除糙隐子草外,其他物种的单株总生物量随放牧强度均呈现逐渐降低的趋势。与对照相比,马蔺和线叶蒿的单株总生物量在中度和重度放牧强度下显著降低;糙隐子草的单株总生物量在轻度和中度放牧强度下显著降低;放牧显著降低了二裂委陵菜的单株总生物量,但不同放牧强度间没有显著差异。与轻度和中度放牧相比,重度放牧显著降低了华北米蒿的单株总生物量。羊草的单株总生物量仅在重度放牧样地显著降低。
2.2 放牧强度对6个物种根冠比的影响
放牧处理对6个物种的根冠比均产生显著影响,且不同物种的影响趋势基本相同(P<0.01)。与对照相比,重度放牧均显著增加了6个物种的根冠比(图4a);轻度放牧对根冠比有一定的促进作用,但影响不显著;中度放牧显著提高了马蔺和二裂委陵菜的根冠比。随放牧强度的增加,6个物种的根冠比均呈上升趋势(图4b)。在每个放牧强度处理不同物种之间的根冠比均存在极显著差异(CK:P=0.003;LG:P<0.001;MG:P<0.001;HG:P<0.001)。在围封样地(CK),糙隐子草的根冠比显著高于羊草和线叶蒿;在轻度放牧样地,华北米蒿的根冠比显著高于线叶蒿、二裂委陵菜和羊草,糙隐子草的根冠比显著高于羊草;在中度放牧样地,二裂委陵菜的根冠比显著高于其他所有物种;在重度放牧样地,二裂委陵菜的根冠比显著高于马蔺、糙隐子草和羊草,华北米蒿的根冠比显著高于糙隐子草和羊草,线叶蒿的根冠比显著高于羊草。
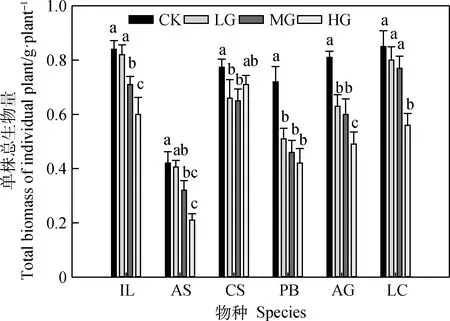
图3 放牧强度对典型草原6个物种单株总生物量的影响Fig.3 Effects of grazing intensity on total biomass of individual plant of six plant species in a typical grassland

图4 放牧强度对典型草原6个物种根冠比的影响Fig.4 Effects of grazing intensity on the R/S ratio of individual plant of six plant species in a typical grassland注:图中不同小写字母表示同一物种的根冠比在不同放牧强度间差异显著(P<0.05),图中**和***表示同一放牧强度下不同物种的根冠比间具有极显著差异(P<0.01和 P<0.001),误差棒表示标准差Note:Different letters indicate significant difference in R/S ratio among different grazing intensities at the 0.05 level,** and ***represent significant difference in R/S ratio among different plant species under the same grazing intensity at the 0.01 and 0.001 level,and the error bar of each treatment represents the standard deviation of five replications
2.3 不同放牧强度影响下物种的重要值与根冠比间的关系
羊草的根冠比与其重要值间呈极显著的负相关关系(r=-0.82,df=10,P=0.001,图5a),但其他5个物种的根冠比与重要值之间呈极显著的正相关关系(r=0.65,df=13,P=0.008,图5b)。
3 讨论
3.1 不同物种的生物量分配格局对放牧强度的响应特征
本研究结果表明随放牧强度的增加,6个物种的单株地上生物量和单株总生物量显著降低,根冠比随放牧强度显著增加,但不同物种之间的生物量分配格局又有所不同。该结果与前期众多群落水平的测定结果类似,群落地上生物量和总生物量均随放牧强度的增加而显著降低[3,20,24]。放牧促使草本植物减少茎的生物量分配比例,植株趋向小型化,从而减少家畜采食和踩踏的比例,这是草本植物面对放牧采取的有效生态适应策略[25-26]。
根系生物量随放牧强度的变化存在较大争议,在佛罗里达的亚热带草原研究结果表明,放牧会增加根系生物量、土壤细根分泌物和微生物碳含量[27]。在藏北高寒草原的研究结果表明中度放牧样地的根系生物量最高[3]。也有研究表明,放牧处理会降低不同草原类型的根系生物量和根系净初级生产力[19]。但根系生物量的分配比例随放牧梯度的显著增加得到了众多研究的支持[21,27-29]。本研究结果表明,6个物种在放牧条件下均会显著提升生物量向根系的分配比例。前期研究结果表明放牧处理会显著降低研究区土壤特别是表层土壤的含水量[30]。基于功能平衡假说,植物光合产物会优先分配至缓解环境胁迫的部位[15]。因此在放牧干扰下,研究区6种植物均通过增加生物量向根系的分配比例来缓解土壤水分胁迫。
相比前期群落或个体水平的生物量分配格局研究,本研究仅简单考虑了生物量在地上和根系两个组分的分配比例,没有综合考虑植物繁殖器官和茎叶的分配比例,这会影响对生物量分配格局这一生态适应对策的深入理解[18,22]。因此,未来的研究要关注放牧强度对植物体不同组分的生物量分配格局的影响,并分析生物量分配格局与物种功能特征间的关系,从而更好的揭示物种生物量分配格局的影响机制。
3.2 不同研究尺度根冠比测定结果间的差异
在温带典型草原和荒漠草原的研究结果表明,个体和群落水平测定的根冠比之间存在较大的差异,本研究结果也支持这一结论[17,31-32]。个体水平测定的根冠比相比群落水平显著偏低,这可能是由于个体水平的根系生物量取样更多针对植物的活根,而群落水平的测定结果关注死根与活根的总生物量,也可能与单株个体根系的取样方法有关。本研究中羊草(根茎型禾草)的根冠比值介于0.23~0.38之间,不同植株羊草个体的根茎之间可能会相互联系,但在单株植物取样时可能只关注单一植物的根系,而群落水平观测时会将单位面积的地下根系全部取样,从而使草原地下密集的根系部分被取样,导致群落水平的根系生物量测定值偏高。因此,未来的研究应该关注不同研究尺度、研究区域和干扰类型下物种的生物量分配格局,从而能更深入的理解生物量分配格局的控制因素和物种的生态适应对策。
3.3 放牧影响下物种的生态适应对策
物种在群落中的地位和作用受其竞争能力、外界因素的干扰(如放牧、刈割、火烧等)、环境条件、种间相互作用等多重因素的共同影响[5,7,11,23,33]。能够更好的适应放牧干扰的物种必将在放牧草原占据群落的优势地位[18,33]。生物量分配格局的改变是物种应对环境胁迫和外界因素干扰常采用的一种生态适应对策。羊草作为原生草原的群落建群种,其在群落中的重要值随放牧强度和根冠比逐渐减低,而糙隐子草、二裂委陵菜和马蔺等物种的重要值则随放牧强度而升高,这可能与不同物种本身的适口性、耐践踏性、株型及生活史类型等有很大关系。羊草和糙隐子草的适口性较好,但糙隐子草相比羊草,株型更小,更加适应放牧导致的干旱环境,从而使其能更好的适应轻度放牧[33]。二裂委陵菜和马蔺的适口性较差,在中度和重度放牧条件下绵羊的选择性采食使其在群落中占据了一定的地位和作用。
本研究结果表明,在不同的放牧强度下,6个物种的根冠比之间均呈现显著差异,这种生物量分配格局的差异体现出不同物种的放牧适应对策。在轻度放牧样地,糙隐子草和华北米蒿相比其他物种显著提高了生物量向根系的分配比例(根冠比显著增加,P<0.001);糙隐子草的单株根系生物量在重度放牧处理下最高,导致糙隐子草的单株总生物量在重度放牧处理下与围封之间无显著差异,这可能是糙隐子草适应高强度放牧的一种有效的生态适应对策。糙隐子草是典型草原适应性和耐牧性均较强的牧草,其作为绵羊最喜食的牧草之一,却在所有放牧强度下均维持较强的优势度[33-34]。结合前期研究和本研究结果,糙隐子草会通过调整其形态可塑性和生物量分配格局来适应放牧干扰,具体表现在,植株趋向于小型化,在减少生殖枝数量的同时,保证其营养枝数和叶片数,并增加植物向根系的生物量分配比例[25-26,33]。二裂委陵菜相比其他物种,在中度放牧样地,显著提高了其根冠比(P<0.001)。这种生物量分配格局的调整使其更能适应放牧导致的环境变化,并促使其成为中度放牧样地的群落亚优势种。因此,本研究结果证明生物量分配格局的调整可能是草地植物适应放牧干扰并占据群落优势地位的一种重要的生态适应对策。
4 结论
典型草原6个物种的单株地上生物量和单株总生物量均随放牧强度的增加而显著降低,放牧导致草本植物的小型化,但放牧强度对单株根系生物量的影响随物种变化而有所不同。放牧处理显著提高了6个物种的根冠比,且在相同放牧强度下不同物种的根冠比之间具有显著差异。糙隐子草在轻度放牧样地和二裂委陵菜在中度放牧样地的生物量分配格局作为其重要的生态适应对策,可能在其占据群落优势地位过程中起到重要作用,其具体的作用机制还有待进一步研究。
猜你喜欢
现代园艺(2021年23期)2021-12-01 07:47:44
现代畜牧科技(2021年7期)2021-07-28 06:40:30
现代畜牧科技(2021年4期)2021-07-21 06:13:04
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
中国科技纵横(2018年14期)2018-08-30 09:15:34
浙江农林大学学报(2018年4期)2018-08-22 01:54:12
现代农业科技(2017年16期)2017-09-22 14:47:46
现代畜牧科技(2016年9期)2016-02-21 12:25:04
江苏农业科学(2014年1期)2014-07-18 21:21:34
