
用Cyt b 基因分析松嫩平原区湖泊水库大银鱼的遗传多样性
2021-01-06 02:34鲁翠云陈昕那荣滨郑先虎李喆唐富江
水产学杂志 2020年5期
鲁翠云,陈昕,那荣滨,郑先虎,李喆,唐富江
(中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070)
大银鱼Protosalanx hyalocranius 隶属于鲑形目Salmoniformes 银鱼科Salangidae,自然分布于我国钱塘江河口、长江口及长江下游湖泊、黄海、渤海沿岸河口及朝鲜半岛西海岸和越南等。银鱼类均为一年生的小型鱼类,大银鱼是银鱼类群中个体较大的物种,曾作为重点经济鱼类在我国北方大、中型湖泊和水库广泛移植[1-3]。近年来,松嫩平原区湖泊水库大银鱼增殖渔业迅速发展,已成为我国最大的大银鱼产区,为渔业提质增效做出了贡献,也对大银鱼资源起到了迁地保护的作用。大银鱼的研究主要集中在分类、生物学、移植增殖生态学上[4-6],在遗传学研究方面开展得相对较少。研究表明大银鱼的同工酶和随机扩增多态性DNA(random amplified polymorphic DNA,RAPD)的多样性均低于太湖新银鱼Neosalanx taihuensis 和寡齿新银鱼 Neosalanx oligodontis[7,8]。
线粒体细胞色素b(Mitochondrial cytochrome b,Cyt b)基因进化速率快,约为16S rRNA 的四倍[9],适合于近缘物种间的系统进化研究。用Cyt b 基因序列研究银鱼科鱼类的系统分化较多。罗宏伟等[10]研究表明三峡库区大银鱼Cyt b 基因序列多态性较低。李大命等[11,12]对大银鱼Cyt b 基因分析表明太湖和洪泽湖群体均为单倍型多样性高而核苷酸多样性低的种群,即经过强捕捞遗传瓶颈效应后伴随着迅速的种群增长与突变的积累而形成。Xiao 等[13]指出淮河大银鱼线粒体Cyt b 基因多样性较高。对于移植到北方的大银鱼种群,Tang 等[14]分析了移植到黑龙江、松花江、兴凯湖等大银鱼的遗传分化,结果显示移植群体的遗传多样性较原种地太湖有大幅增加。在此基础上,本研究利用Cyt b 全序列分析了移植到北方松嫩平原区湖泊、水库的大银鱼群体的遗传分化,可为大银鱼种群的遗传管理和持续利用提供支持。
1 材料与方法
1.1 材料
大银鱼采自松嫩平原区的5 个湖泊水库,分别为黑龙江省大庆市连环湖(LHH)72 尾和龙虎泡(LHP)49 尾,吉林省松原市查干湖(CGH)54 尾、长春市新立城水库(XLC)22 尾和四平市二龙山水库(ELS)29 尾,共226 尾。分别取其尾鳍,无水乙醇固定后,-20℃冷冻保存备用。
1.2 基因组DNA 提取
采用传统的酚氯仿抽提法从大银鱼鳍条组织中提取基因组DNA[15],使用NanoDropTM8000 分光光度计检测所提取DNA 的浓度及纯度,稀释至50 ng/μL,4℃保存备用。
1.3 PCR 扩增与测序
使用引物L14321 和H15634 扩增大银鱼Cyt b基因全序列[16],L14321 引物序列为5’-CCAGTGA CTTGAAAAACCACCG-3’;H15634 引物序列为5’-CTTAGCTTTGGGAGTTAAGGGT-3’,引物由上海生工生物公司合成。PCR 反应体系为25 μL,其中模板DNA 2 μL、混合PCR 缓冲液buffer 18 μL(10 mmol/L Tris-Cl(pH8.0)、50 mmol/L KCl、1.5 mmol/L MgCl2、200 μmol/L dNTP)、上下游引物(10 μmol/L)各0.5 μL、Taq DNA 聚合酶1.5 U,其余体积用去离子水补充。扩增反应在ABI 9700 型PCR 仪上完成,反应程序为:94℃预变性7 min;94℃变性30 s,56℃退火45 s,72℃延伸1 min,30 个循环;72℃延伸5 min。PCR 产物经2%琼脂糖凝胶电泳检测后,由上海生工生物工程技术服务有限公司纯化后进行双向测序及拼接。
1.4 序列分析
测序后,将序列输入Clustal X 软件[17]进行序列的对位排列,并加以人工校对,截取相同长度的序列用于群体遗传分析。用DnaSP v5 软件[18]统计变异位点类型和数目、计算单倍型数和核苷酸多态性等;用MEGA 7.0 软件[19]分析序列的碱基组成和差异百分比、变异位点、简约信息位点数、转换/颠换值,用Kimura 2-Parameters 方法计算5 个群体间的遗传距离,分别构建基于邻接法(Neighbor-Joining,NJ)和非加权配对算术平均法(unweighted pair group method using arithmetic average,UPGMA)的群体间发生关系的聚类树,采用Bootstrap(重复数=1 000)检验分子系统树各分支的置信度。利用Arlequin 3.11软件[20]中的分子方差分析(AMOVA)计算群体间的遗传分化系数(Fst)及遗传变异组成。
2 结果与分析
2.1 核苷酸和氨基酸序列的特征
将所得序列在GenBank 中进行Blast 比对,结果确定为大银鱼线粒体Cyt b 基因的全序列[21]。所有序列经Clustal X 比对并加以人工校对后,截取1 141 bp 序列用于群体遗传分析。经MEGA 7.0 分析,此序列可编码380 个氨基酸。保守位点1 081个,变异位点60 个,其中单变异位点32 个,简约信息位点28 个。测得的序列中A、T、C、G 的碱基组成分别为21.73%、29.27%、32.33%和16.67%,其中A+T 含量(51%)略高于C+G 含量(49%),转换/颠换值为3.808。
2.2 单倍型及核苷酸多样性
5 个群体226 个样本的Cyt b 基因可分为38 个单倍型,其中单倍型Hap 4 的个体数最多(31.42%),其次为单倍型Hap 7(22.57%),且Hap 4和Hap 7 的个体分布于5 个群体中;单倍型Hap 3(8.85%)和单倍型Hap 15(3.98%)的个体分布于4个群体中,单倍型Hap 6(8.85%)的个体分布于3 个群体中,其余单倍型分布较少。各单倍型在群体中的分布见表1。
5 个大银鱼群体总体单倍型多态性(nh)为(0.833±0.016),核苷酸多样性(Pi)为(0.00209±0.00020)。每个群体的单倍型数(nh)为5~16 个,LHP最少而LHH 最多;单倍型多样性(Hd)在(0.656±0.051)和(0.939±0.028)之间,核苷酸多样性(Pi)在(0.00121±0.00010)和(0.00496±0.00150)之间,LHP 多样性最低而XLC 多样性最高。Tajima's D 中性检验值的变化范围为-1.89133~0.55966,中性检验结果显示只有XLC(P<0.05)显著不符合中性突变,其他4 个群体均不显著,符合中性突变(表2)。
2.3 群体遗传分化
Cyt b 基因的AMOVA 结果表明,各群体间的遗传差异主要来自群体内(93.81%),少数来自群体间(6.19%)(表3)。群体间遗传分化系数(Fst)为0.06185,各群体间的遗传分化系数(Fst)为0.01537~0.13572,除了查干湖与龙虎泡分化不显著外,其余群体间分化显著(P<0.05),查干湖和连环湖、新立城和龙虎泡、二龙山水库与龙虎泡和查干湖达到了极显著的遗传分化水平(表4)。
2.4 遗传距离和系统进化树的构建
利用Cytb 基因序列,采用MEGA 4.0 软件,根据Kimura 双参数模型计算各群体之间的遗传距离(D)。结果显示:XLC 和ELS 之间遗传距离最大(0.0040),LHP 和LHH 之间遗传距离最小(0.0014)(表4)。根据各群体之间的遗传距离构建的系统进化树显示,CGH 和LHP 聚为一支后与LHH 聚为一支,再与ELS 聚为一支,XLC 独立为一支(图1)。
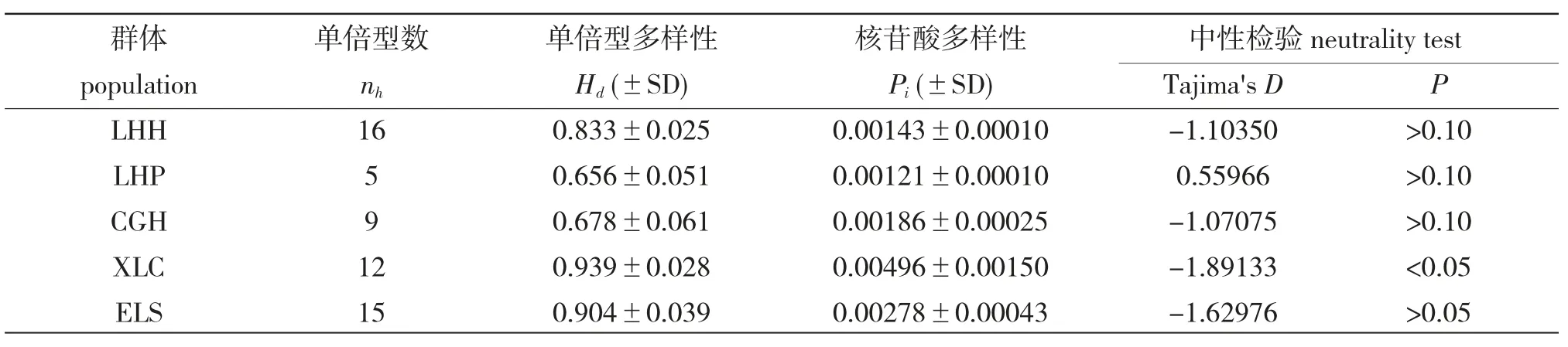
表2 基于线粒体Cyt b 基因的遗传多样性Tab.2 Genetic diversity estimates in the five populations of clearhead icefish based on Cyt b gene of mtDNA
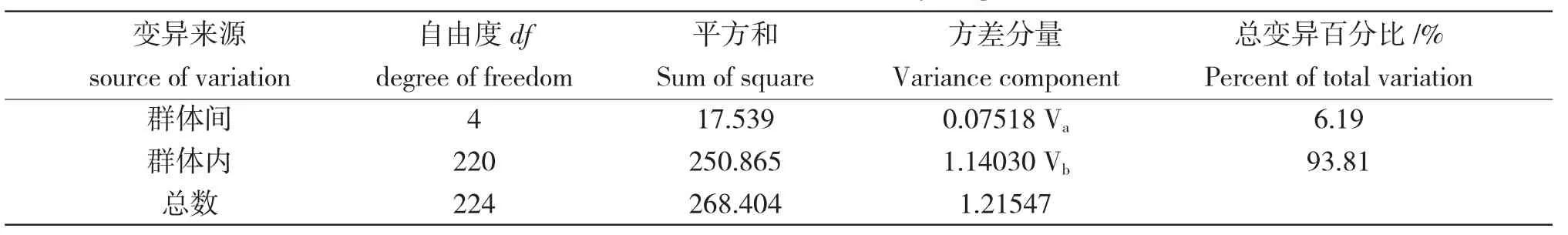
表3 基于线粒体Cyt b 基因的AMOVA 分析结果Tab.3 The result of AMOVA based on Cyt b gene of mtDNA

表4 基于Cyt b 基因的各群体间的Fst 和基于Kimura 双参数模型计算的各群体间遗传距离Tab.4 The Fst among populations based on Cyt b gene and genetic distance calculated based on Kimura 2-parameter model in each population
3 讨论
银鱼营养丰富、口感好、经济价值高,是我国重要的出口创汇水产品之一。自20 世纪90 年代以来,陆续移植到全国27 个省(市)的湖泊、水库进行养殖[23],其中大银鱼个体大、冬季繁殖的生物学特性,使其具有较强的适应能力,特别是对寒冷有较强的耐受力,而在北方的湖泊水库成功移植,在诸多省份形成了产量,获得了显著的经济效益和社会效益[2]。
线粒体DNA(mtDNA)具有严格的母系遗传、进化速率快、几乎无重组等特点,被广泛用于鱼类系统发育和群体遗传研究。其中Cyt b 基因作为重要的蛋白质编码基因,进化速率适中,能有效反映属、亚种、种群等不同群体水平的遗传信息[22],广泛应用于银鱼系统进化及遗传资源评估中[9-13]。用Cyt b 基因序列在原产地太湖和洪泽湖大银鱼群体中分别检测到12 个和7 个单倍型,单倍型多样性分别为(0.850±0.045)和(0.775±0.045),核苷酸多样性分别为(0.00296±0.00017)和(0.00129±0.00010)[11,12]。本研究中,在松嫩平原区3 个湖泊和2 个水库大银鱼群体中共检测到38 个单倍型,单倍型多样性为(nh)和核苷酸多样性(Pi)分别为(0.833±0.016)和(0.00209±0.00020),略低于太湖群体而高于洪泽湖群体,其中连环湖群体和二龙山群体的单倍型数高于太湖,新立城水库和二龙山水库的遗传多样性指标也高于太湖群体,龙虎泡和查干湖群体遗传多样性较低。连环湖是嫩江下游平原型湖泊,本研究的样本分别采自连环湖最下游的水体阿木塔泡和牙门喜泡,而所有连环湖水均经阿木塔泡后流入嫩江,因此,上游水源带来的大银鱼增加了该水体大银鱼的遗传多样性。新立城水库和二龙山水库均为第二松花江支流的水库,均采用人工投放受精卵的方式维持遗传多样性;两水库不仅与松嫩平原湖泊进行大银鱼种质交换,也与南方湖泊进行了较多的种质交换,增加了本地区其他湖泊不具备的单倍型,因此保持了较高的遗传多样性水平[23,24]。龙虎泡和查干湖均为独立的水体,与外界水体沟通较少,增加遗传多样性的措施也为投放不同来源受精卵,但本研究显示二者遗传多样性较低,应注重引进更广泛水体的大银鱼受精卵。
AMOVA 结果表明:大银鱼遗传差异主要来自群体内(93.81%),少数来自群体间(6.19%)。虽然群体间遗传分化系数较低(Fst=0.06185),但是两两群体间遗传分化均达到显著水平或极显著水平,显示出各水体不同批次移植、投放大银鱼受精卵的特征;而查干湖与龙虎泡群体分化不显著,可能是两水体常态化交换投放受精卵的结果。聚类分析结果表明松嫩平原区湖泊连环湖、查干湖、龙虎泡首先聚类,然后再与第二松花江支流的二龙山水库和新立城水库聚类,体现出嫩江下游平原碱性湖泊遗传相似度高而松花江两水库与其他各水体相似度均低的特征。
大银鱼在土著水域正面临着资源衰退的问题,其广泛移植不仅创造了很好的经济效益和社会效益,也增加了我国这一独有物种的遗传多样性,这对增强该物种的可持续利用具有重要意义。但是,大银鱼的移植增殖会明显抑制其他鱼类种群发展,特别是小型上层鱼类,如属Hemiculter sp.鱼类等。因此,认为不应在水生生物保护区水域进行大银鱼移植增殖;同时,在管理大银鱼种群时要根据水体环境容纳量确定大银鱼卵投放量和亲鱼保留量,防止过度放养和过度消耗饵料生物资源,保护其他鱼类资源是大银鱼种群可持续发展和渔业利用的基础。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
读友·少年文学(清雅版)(2020年5期)2020-09-09
读友·少年文学(清雅版)(2020年5期)2020-09-09
读友·少年文学(清雅版)(2020年5期)2020-09-09
红领巾·萌芽(2019年8期)2019-08-27
作文与考试·小学高年级版(2019年24期)2019-01-06
时代青年·视点(2017年1期)2017-02-17
农产品市场周刊(2017年2期)2017-01-16
