
中国黄渤海真江蓠(Agarophyton vermiculophyllum)群体遗传多样性研究
2021-01-04 09:57:12钟凯乐宋小含段德麟胡自民
海洋科学 2020年12期
钟凯乐, 宋小含, 段德麟, 胡自民
中国黄渤海真江蓠(Agarophyton vermiculophyllum)群体遗传多样性研究
钟凯乐1, 2, 3, 宋小含1, 2, 3, 段德麟1, 2, 胡自民1, 2
(1. 中国科学院海洋研究所 海洋大科学研究中心 实验海洋生物学重点实验室, 山东 青岛 266071; 2. 青岛 海洋科学与技术试点国家实验室 海洋生物学与生物技术功能实验室, 山东 青岛 266237; 3. 中国科学院大学, 北京 100049)
真江蓠是原产于西北太平洋的重要经济红藻。我们利用10对微卫星引物检测中国黄渤海地区真江蓠的群体遗传多样性和结构。10个微卫星位点在12个群体中共检测到65个等位基因, 每个位点的等位基因(a)为1~28, 有效等位基因(e)为1.0~9.6。每个群体的平均等位基因(a)、平均有效等位基因(e)、平均香浓指数()、平均观察杂合度(o)和平均预期杂合度(e)分别为2.4、1.6、0.419、0.133和0.227, 显示较低的群体遗传多样性。中国黄渤海12个真江蓠群体间遗传分化较大(st=0.398 7), 基因流有限(m=0.377 1), 近交系数为正(is=0.391 3,it=0.634 0), 表明可能存在近交和杂合子缺失现象。Structure和UPGMA系统进化分析一致将12个群体分为两个遗传组, 并在黑石礁群体(HS)和石岛群体(SD)中发现明显的遗传混杂现象。AMOVA分析显示遗传变异主要来自于群体内(73.27%)。该研究可为黄渤海地区真江蓠自然资源保护和管理提供科学依据。
真江蓠(); 微卫星; 遗传多样性; 基因流; 黄渤海
气候变化和近岸环境变迁是影响潮间带海洋生物群体遗传多样性和分化的重要因素。黄渤海作为太平洋西部的边缘海, 形成于第四纪更新世晚期[1], 在末次盛冰期(Last Glacial Maximum, LGM)经历了剧烈的气候和海岸环境变迁。更新世冰期导致全球海平面下降120~140 m[2], 使得西北太平洋边缘海面积缩小, 黄渤海陆架几乎完全暴露成为陆地, 东海向东缩减变为狭长的冲绳海槽[2-3]。边缘海面积的缩减导致海洋物种大量减少, 残遗物种只能在避难所生存, 因此避难所的不连续性成为不同群体之间隔离和分化的重要因素[4], 其中冲绳海槽被证实是多种海洋生物的冰期避难所[3, 5]。在更新世间冰期, 气温回暖海平面上升, 残遗物种从避难所扩展到新的栖息地。这种群体扩张可能导致来自不同避难所的群体之间的遗传交换, 消除了隔离造成的群体分化[6]。除此之外, 黄渤海的洋流系统对海洋生物的遗传多样性和分化也具有重要影响[7]。主导黄渤海海域的洋流主要是北向的黑潮分支黄海暖流和南向的中国沿岸流, 黄海暖流和中国沿岸流驱动的群体遗传均质性在鼠尾藻()[7-8]和角叉菜()[5]等多种海洋生物中已有报道。
真江蓠()隶属于红藻门(Rhodophyta), 真红藻纲(Florideophyceae), 江蓠目(Gracilariales), 江蓠科(Gracilariaceae),属, 是原产于亚洲-西北太平洋的一种食用红藻, 也是一种在世界范围内广泛分布的入侵种[9]。真江蓠不仅是提取琼胶的主要原料, 而且是重要的生态奠基物种, 因此具有重要的经济和生态价值[10-11]。除此之外, 真江蓠还可作为生物过滤器净化水质, 在海水富营养化修复和多营养层次综合水产养殖系(Integrated Multi-Trophic Aquaculture, IMTA)中都具有广泛应用[12-13]。
目前, 相对于真江蓠的入侵, 对该物种的群体遗传学研究仍然较少, 且大多基于单一的线粒体分子标记。Krueger-Hadfield等[9]基于线粒体1 (~1 200 bp)检测到黄渤海地区真江蓠群体为一个单一遗传谱系, 但该研究主要涉及朝鲜半岛西海岸群体, 中国沿海采样点极少。刘若愚等[14]基于1(641 bp)在中国黄渤海沿岸真江蓠群体中检测到10个单倍型, 为一个共同的遗传谱系, 群体间遗传分化较弱[14]。Hu等[15]基于AFLP(Amplified Fragment Length Polymorphism)发现黄渤海地区真江蓠群体为共同的遗传谱系, 并且在青岛地区群体中发现小尺度的遗传分化。
微卫星具有遗传信息丰富、多态性高、中性选择、共显性遗传等特点, 重复性好、易于检测、稳定性高, 在海藻群体遗传多样性研究中具有广泛应用[16-17]。本研究共收集中国黄渤海12个群体的196株真江蓠样品, 利用微卫星数据旨在深入分析该地区真江蓠的群体多样性特征, 同时检测是否在小地理尺度存在遗传结构和遗传变异的时空分布模式。该研究有助于理解气候变化与当代环境的相互作用对海藻遗传变异的影响, 从而为真江蓠自然资源保护和管理提供科学依据。
1 材料与方法
1.1 真江蓠样本采集
我们采集到来自辽东半岛和山东半岛潮间带12个地点的真江蓠样本(表1)。在每个采样点, 随机采集6~20个真江蓠个体, 确保个体之间距离大于10 m以最大程度避免采集到遗传一致的个体。样品清洗干净后置于硅胶中干燥保存。
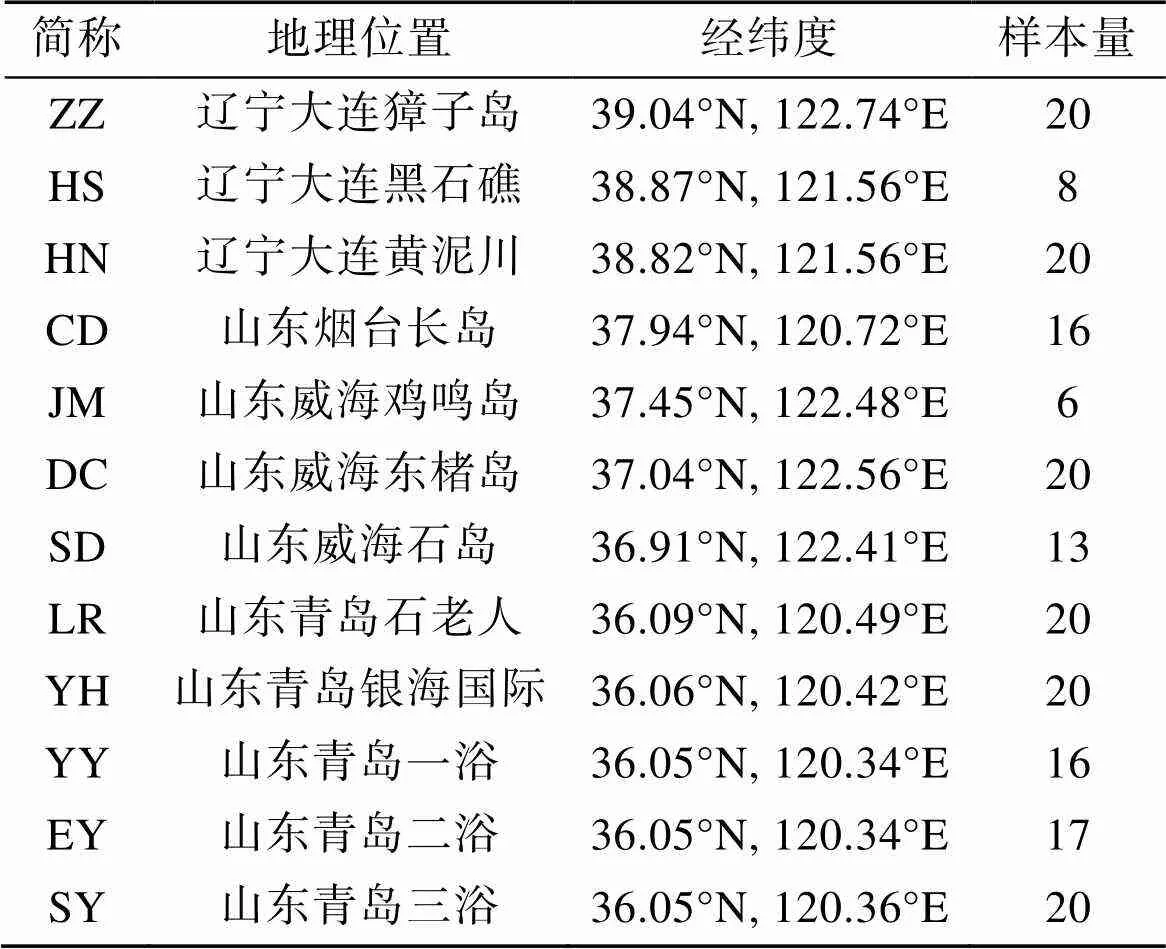
表1 黄渤海真江蓠样本采集信息
1.2 DNA提取与检测
取30 mg真江蓠样本, 在液氮中冷冻并研磨成粉末, 使用诺贝莱一步法多糖多酚植物基因组DNA抽提试剂盒DNE379(Nobelab)提取DNA, 具体操作见说明书。1%琼脂糖凝胶电泳(140 V, 30 min)检测DNA质量。
1.3 PCR扩增和电泳检测
我们使用Kollars等[18]和Krueger-Hadfield等[19]开发的10对真江蓠微卫星引物进行片段扩增(表2), 引物由北京擎科生物科技有限公司青岛分公司合成。本实验我们采用双重荧光检测, F正向引物的5′端连接荧光基团(FAM或HEX)。PCR扩增反应体系15 μL: 模板DNA 1.0 μL(约15 ng), 2*Tsingke Master mix 7.5 μL, 正向引物和反向引物各1.0 μL(10 μmol/L),灭菌水4.5 μL。PCR反应程序: 98℃预变性2 min; 98℃变性10 s, 56℃退火10 s, 72℃延伸10 s, 30个循环; 最终72℃延伸5 min。PCR扩增产物在3730型遗传分析仪(ABI, USA)上进行毛细管电泳检测(北京擎科生物科技有限公司青岛分公司)。
1.4 数据分析
利用软件Gene mapper v4.1读取数据并汇总。Popgene v1.32[20]和GenAlex v6.501[21]计算各微卫星位点和群体的遗传参数。包括等位基因数(a), 有效等位基因数(e), 香浓指数(), 观察杂合度(o), 预期杂合度(e), 哈迪-温伯格平衡偏离指数(=o–e/e),统计量(is,it和st), 基因流(m),s遗传距离和相似性。
用Poptree v2[22]构建基于遗传距离D的UPGMA聚类树, 设置自展值为(bootstrap)1 000。在Structure v2.3.4[23]中基于贝叶斯聚类法对群体遗传结构进行分析。参数设置为: Length of Burnin period为105, Number of MCMC Reps after Burnin为106, K值设置为1~5, 每个K值重复运算10次。基于Delta K=mean(|L″(K)|)/ sd(L(K)), 在Structure Harvester中确定最佳K值, 即最佳分类群数。
在Arlequin v3.5.1.3[24]中进行分子方差分析(AMOVA), 基于104次重复抽样检测群体间和群体内的遗传结构和遗传变异的分布。

表2 真江蓠10个微卫星位点的信息
2 结果
2.1 遗传参数
10个微卫星位点共检测出65个等位基因, 每个微卫星位点等位基因(a)为1~28, 有效等位基因(e)为1.000~9.614。其中Gverm 5276和Gverm 7969分别具有最高和最低的等位基因和有效等位基因数目(表2)。
每个群体在10个微卫星位点的遗传参数见表3。各群体的等位基因数(a)为1.4~3.5, 平均等位基因数为2.4; 有效等位基因数(e)为1.123~2.346, 平均有效等位基因数为1.644; 香浓指数()为0.120~ 0.649, 平均香浓指数为0.419; 观测杂合度(o)和预期杂合度(e)分别为0.012~0.276和0.073~0.349, 平均观测杂合度和预期杂合度分别为0.133和0.227, 各群体的观测杂合度均低于预期杂合度。12个群体中哈迪-温伯格偏离指数()均为负值, 表现为杂合子缺失。
2.2 群体遗传结构
12个真江蓠群体的-统计量及基因流分析(表4) 显示, 近交系数is为–0.506 4~0.855 8, 平均近交系数is为0.391 3, 其中Gverm 1803和Gverm 5276的近交系数为负值; 各位点的总群体近交系数it为–0.144 8~0.770 4, 平均值为0.634 0, Gverm 5276为负值; 度量群体间遗传差异程度的st为0.000 0~ 0.730 6, 平均值为0.398 7, 遗传分化程度较高; 基因流m为0.092 2~2.214 4, 平均值为0.377 1, 不同位点的基因流值变化较大。
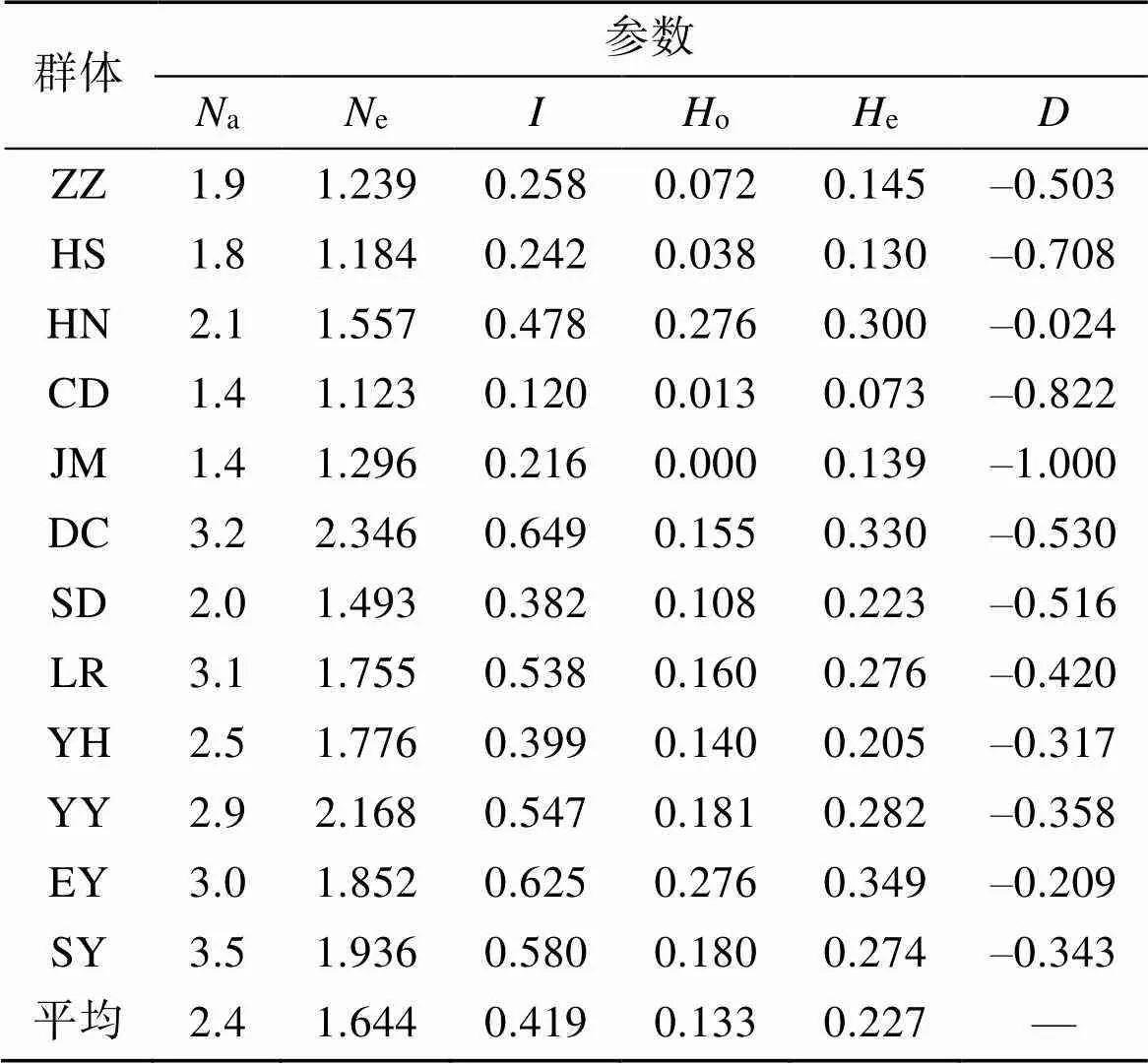
表3 10个微卫星位点在12个真江蓠群体的遗传多样性参数
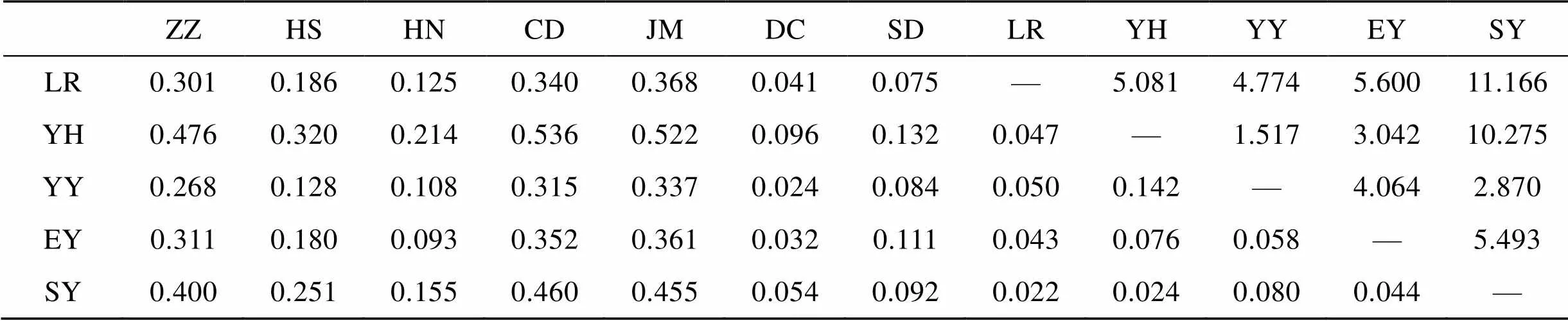
12个真江蓠群体s遗传相似性及遗传距离见表5。各群体间遗传相似性指数以银海国际(YH)和三浴(SY)之间最大(0.987), 鸡鸣岛(JM)和银海国际(YH)之间最小(0.497); 相应的,s遗传距离以鸡鸣岛(JM)和银海国际(YH)之间最大(0.699), 银海国际(YH)和三浴(SY)之间最小(0.013)。
12个真江蓠群体的遗传分化系数(st)和基因流(m)见表6。群体间最大的遗传分化在长岛(CD)和银海国际(YH)之间(0.536), 最小在石老人(LR)和三浴(SY)之间(0.022)。最大基因流发生在三浴(SY)和石老人(LR)之间(11.166), 最小基因流发生在银海国际(YH)和长岛(CD)之间(0.216)。山东半岛南侧的群体之间遗传分化较小, 基因交流频繁。

表4 F-统计量和基因流
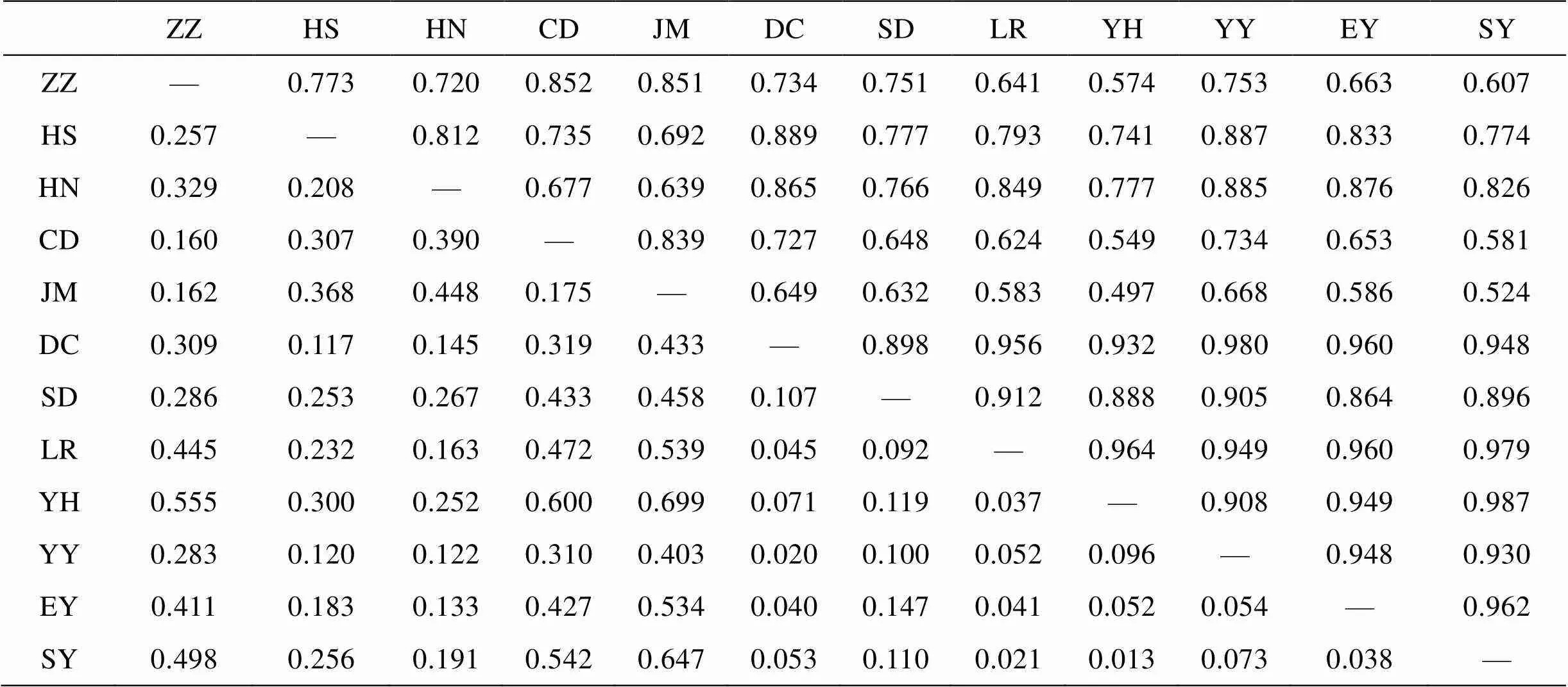
表5 12个真江蓠群体间的Nei’s遗传相似性(上三角)和遗传距离(下三角)
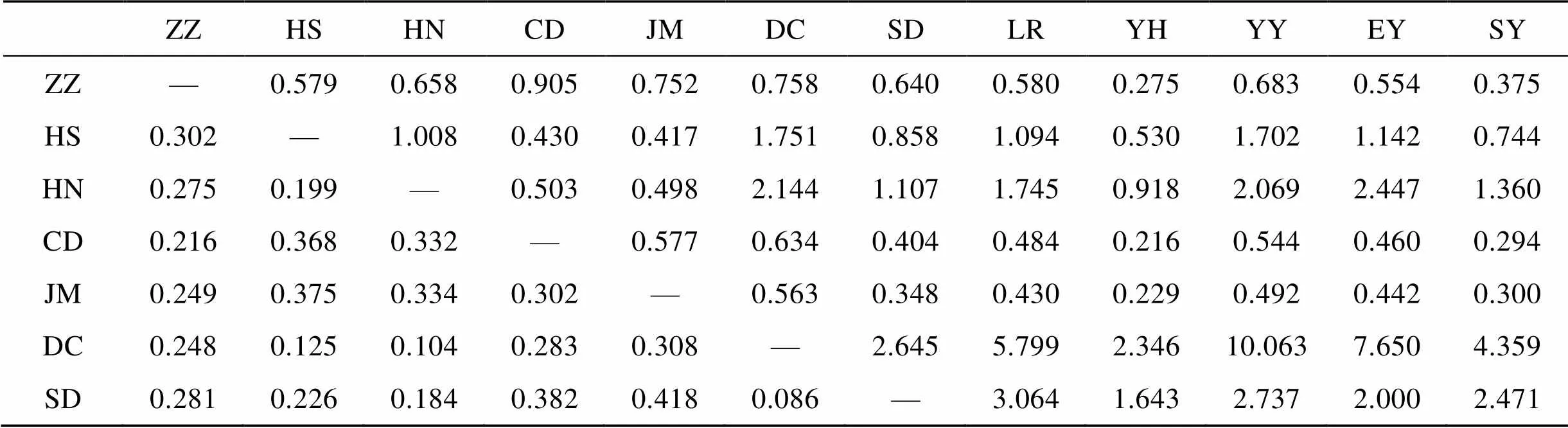
表6 12个真江蓠群体间的遗传分化系数Fst(下三角)和基因流Nm(上三角)
续表
Structure分析显示黄渤海12个真江蓠群体最佳遗传聚类数K=2(图1), 与UPGMA系统发育树结构一致(图2)。獐子岛(ZZ)、长岛(CD)和鸡鸣岛(JM)为一个遗传组, 其他群体聚为第二个遗传组, 其中黑石礁(HS)和石岛(SD)具有明显的遗传混杂现象。AMOVA分析显示群体间遗传变异占36.73%, 群体内变异占 73.27%, 表明群体内遗传变异是引起黄渤海真江蓠群体变异的主要因素(表7)。
3 讨论
等位基因、有效等位基因、杂合度等是判断群体遗传多样性的重要指标, 种群的遗传多样性越高其环境适应能力越强, 相应的进化潜力也越大。本研究中12个真江蓠群体的a(2.4),e(1.644),(0.419),o(0.133)和e(0.227)等遗传参数较低, 与刘若愚等[14]利用1(d=0.300, π=0.050×10–2)和Hu等[15]利用AFLP(Na, 0.336~1.224; Ne, 1.057~1.142; I, 0.059~0.161)检测的结果基本吻合, 一致表明中国黄渤海地区真江蓠群体遗传多样性和变异程度较低, 环境适应能力可能较差。is(0.391 3)、it(0.634 0)和哈迪-温伯格平衡偏离指数(<0)表明该地区真江蓠群体普遍存在杂合子缺失现象。这些遗传特征可能与真江蓠的繁殖方式和繁殖特点有关。在自然界, 相对于四分孢子体植株, 江蓠科海藻雌雄配子体植株数量较少[25-26]。真江蓠等红藻进行有性繁殖时, 释放的四分孢子和精子存活时间较短, 没有鞭毛结构且不具自主游动能力, 只能随着水流在近距离的雌配子果胞上完成受精[27], 这种有限距离的扩散降低了群体间遗传交换, 导致群体遗传多样性偏低。此外, 真江蓠也可通过断枝方式进行营养繁殖[28], 而营养繁殖的方式也大大降低了真江蓠的杂合度和群体遗传多样性。

图1 基于10个微卫星的12个真江蓠群体的遗传结构图(底图审图号: GS(2019)1711号)
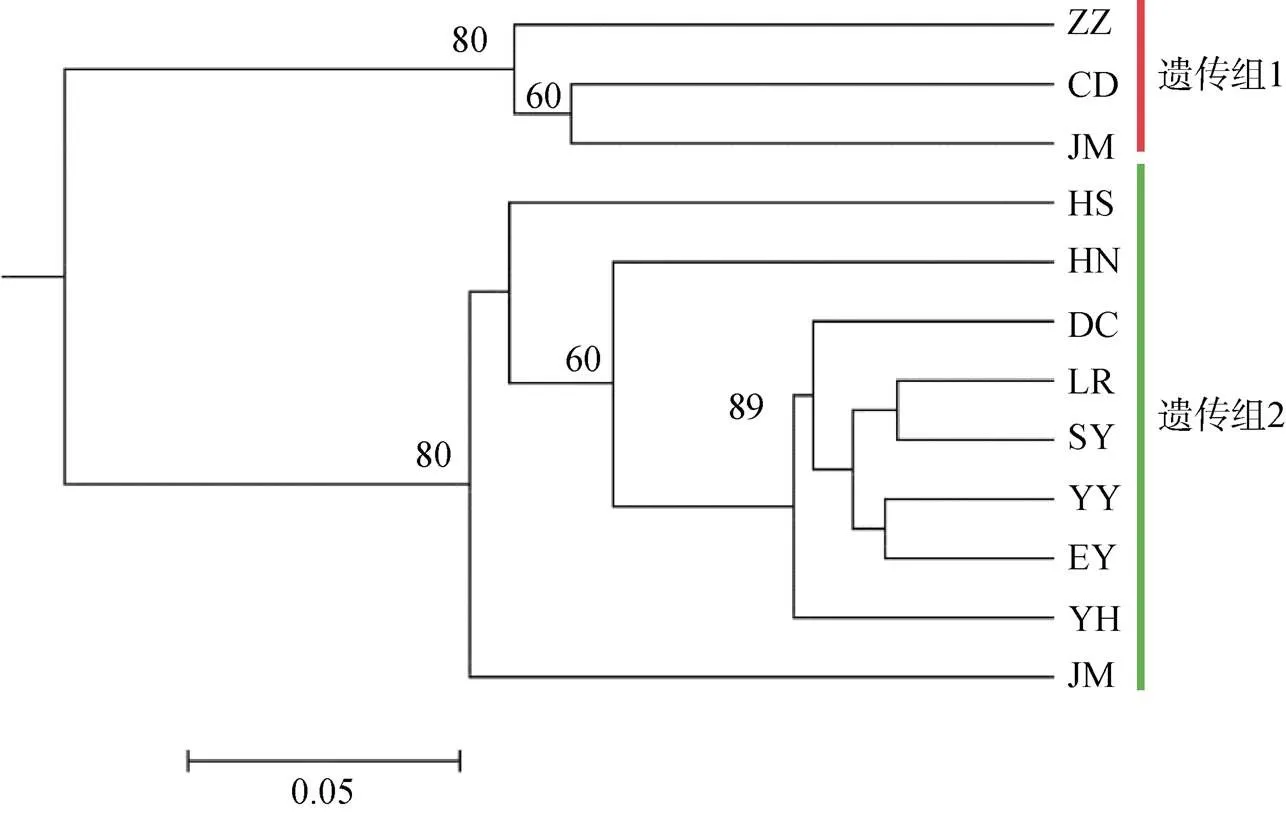
图2 基于Nei’s遗传距离构建的12真江蓠群体的UPGMA聚类树
注: 分支上的数字表示检验重复1 000 次所得的大于50%的支持率
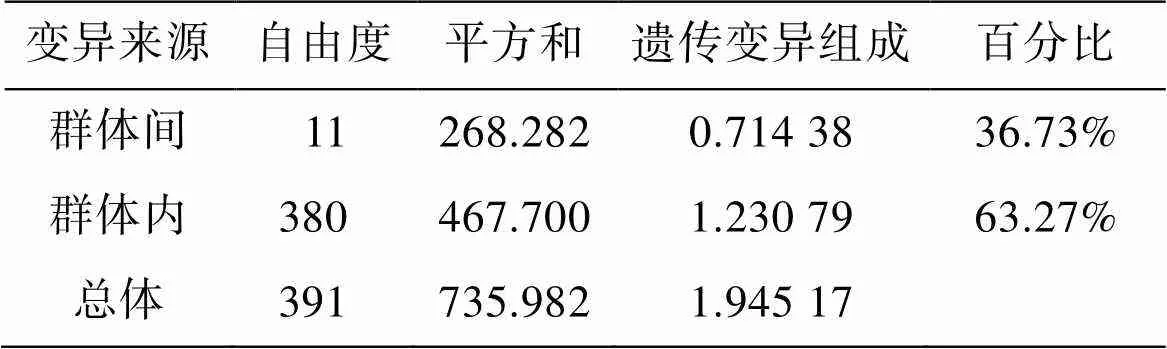
表7 12个真江蓠群体的分子方差分析
通过群体间遗传分化系数st、s遗传距离和基因流m可知黄渤海地区真江蓠群体之间遗传分化较大, 基因流有限。Krueger-Hadfield等[9]利用微卫星在日本地区真江蓠群体间检测到类似的较高的群体间遗传分化模式, 并证实遗传分化系数st与地理距离呈正相关。Hu等[15]利用AFLP也发现黄渤海真江蓠群体间较高的遗传分化, 与本研究结果一致。除了真江蓠的繁殖特点外, 群体间栖息地的不连续性造成的地理隔离可能是另一个驱动因素[29]。真江蓠可适应多种栖息环境, 在河口和海岸潮间带环境均有分布, 生境的不连续性和异质性(例如: 泥滩、沙滩、礁石和石砾)可能导致了群体水平的遗传分化。此外, 这种小地理尺度的遗传分化可能也与微地理环境差异有关[30], 例如盐度与潮汐。黄渤海不同季节和地区盐度变化较大[31], 而真江蓠可以适应5‰~ 60‰的盐度变化[32]。已有报道显示盐度梯度可对等位基因频率产生强选择(例如,)[33], 对无融合生殖和表型变异产生影响并最终导致基因型分化(例如,)[34]。山东半岛和辽东半岛毗邻黄海, 属于温带季风气候, 全年处于不规则半日潮中。高滨岸和低滨岸之间的潮汐强度也可能对真江蓠产生选择压力, 从而产生微地理尺度的遗传分化。
通过Structure遗传结构可知, 中国黄渤海12个真江蓠群体分化为2个遗传组, 但是并没有发现如Hu等[15]在青岛地区检测到的特有遗传分化现象。Hu等[15]检测到黄渤海真江蓠地理障碍发生在JM和ZZ群体间, 而本研究发现这种遗传隔离发生在山东半岛的东南部(JM和DC群体之间), 类似的现象在该地区并不常见。众多研究表明, 由于更新世冰期海平面下降, 现今黄渤海地区的海洋生物大多起源于冲绳海槽避难所, 并在间冰期随着黑潮、黄海暖流和中国沿岸流向黄渤海和东海地区扩张迁移, 故该地区的海洋生物大多属于同一遗传谱系, 具有较强的遗传均质性[5, 7-8]。因此, 我们推测这种遗传隔离可能与山东半岛形成的地理屏障和真江蓠有限传播能力共同造成的基因交流障碍有关。
刘若愚[35]检测到真江蓠的基因流方向为辽东半岛至山东半岛至青岛, 与该地区洋流运动相一致, 这可能是山东半岛南部真江蓠群体(SD)出现北部基因型(遗传组1)的重要原因。我们在辽东半岛的群体中(HS和HN)发现山东半岛南部群体的主要基因型(遗传组2), 推测这种逆洋流方向的遗传混杂可能与辽东半岛和山东半岛之间频繁的海上航运等人为因素有关。虽然真江蓠的孢子和精子运动能力有限, 但作为一个世界范围内的入侵种它可以附着在船体、缆绳、鱼竿、虾和蟹笼等硬基质上, 通过海上运输、休闲渔业和水产养殖活动等传播扩散[36]。
4 结论
本研究基于10对微卫星引物对12个黄渤海真江蓠群体进行群体遗传多样性分析, 发现该地区真江蓠群体遗传多样性较低, 杂合子缺失, 并在山东半岛东南部检测到罕见的遗传隔离。该研究对黄渤海海藻多样性形成和自然资源保护提供了科学基础。
[1] 耿秀山. 黄渤海地貌特征及形成因素探讨[J]. 地理学报, 1981, 36(4): 423-434. Geng Xiushan. The geomorphological features and forming factors of submarine relief in the Bohai Sea and the Yellow Sea[J]. Acta geographica sinica, 1981, 36(4): 423-434.
[2] Lambeck K, Esat T M, Potter E K. Links between climate and sea levels for the past three million years[J]. Nature, 2002, 419(6903): 199-206.
[3] Wang P X. Response of Western Pacific marginal seas to glacial cycles: Paleoceanographic and sedimentological features[J]. Marine Geology, 1999, 156(1-4): 5- 39.
[4] Ni G, Li Q, Kong L F, et al. Comparative phylogeography in marginal seas of the northwestern Pacific[J]. Molecular Ecology, 2014, 23(3): 534-548.
[5] Hu Z M, Li J J, Sun Z M, et al. Phylogeographic structure and deep lineage diversication of the red algaHolmes in the Northwest Pacific[J]. Molecular Ecology, 2015, 24(19): 5020-5033.
[6] Xue D X, Wang H Y, Zhang T, et al. Population genetic structure and demographic history ofbased on mitochondrial DNA and microsatellite markers[J]. PLoS One, 2014, 9: e95436.
[7] Li J J, Hu Z M, Gao X, et al. Oceanic currents drove population genetic connectivity of the brown algain the north-west Pacific[J]. Journal of Biogeography, 2017, 44: 230-242.
[8] Liu J, Zhan D, Zhang R, et al. AFLP analysis revealed a north to south genetic break in the brown algaalong the coast of China[J]. Journal of Applied Phycology, 2018, 30(4): 1-9.
[9] Krueger-Hadfield S A, Kollars N M, Strand A E, et al. Genetic identification of source and likely vector of a widespread marine invader[J]. Ecology and Evolution, 2017, 7(12): 4432-4447.
[10] 史升耀, 张燕霞, 范晓, 等. 碱处理对中国江蓠属海藻所含琼胶的作用[J]. 水产学报, 1988, 12(2): 145-155.Shi Yaosheng, Zhang Yanxia, Fan Xiao et al. The effects of alkali treatment on agars from chinese species of[J]. Journal of fisheries of China, 1988, 12(2): 145-155.
[11] Davoult D, Surget G, Stiger-Pouvreau V, et al. Multiple effects of ainvasion on estuarine mudflat functioning and diversity[J]. Marine Environmental Research, 2017, 131: 227-235.
[12] 徐姗楠, 温珊珊, 吴望星, 等. 真江蓠()对网箱养殖海区的生态修复及生态养殖匹配模式[J]. 生态学报, 2008, 28(4): 1466-1475.Xu Shannan, Wen Shanshan, Wu Wangxing, et al. Biremediation of caged fish aquaculture by the red algain an integrated multi-trophic aquaculture system[J]. Acta Ecologica Sinica, 2008, 28(4): 1466-1475.
[13] Abreu M H, Pereira R, Yarish C, et al. IMTA with: Productivity and nutrient removal performance of the seaweed in a land-based pilot scale system[J]. Aquaculture, 2011, 312(1-4): 77-87.
[14] 刘若愚, 孙忠民, 姚建亭, 等. 中国近海重要生态建群红藻真江蓠的群体遗传多样性[J]. 生物多样性, 2016, 24(7): 781-790. Liu Ruoyu, Sun Zhongmin, Yao Jianting, et al. Genetic diversity of the habitat-forming red algaalong Chinese coasts[J]. Biodiversity Science, 2016, 24(7): 781-790.
[15] Hu Z M, Liu R Y, Zhang J, et al. A unique genetic lineage at the southern coast of China in the agar-producing(Gracilariales, Florideophyceae)[J]. Algae, 2018, 33: 269-278.
[16] Tatarenkov A, Jönsson R B, Kautsky L, et al. Genetic structure in populations of(Phaeophyceae) over spatial scales from 10 m to 800 km[J]. Journal of Phycology, 2007, 43(4): 675-685.
[17] Muhlin J F, Engel C R, Stessel R, et al. The influence of coastal topography, circulation patterns, and rafting in structuring populations of an intertidal alga[J]. Molecular ecology, 2008, 17(5): 1198-1210.
[18] Kollars N M, Krueger-Hadfield S A, Byers J E, et al. Development and characterization of microsatellite loci for the haploid-diploid red seaweed[J]. PeerJ, 2015, 3: e1159.
[19] Krueger-Hadfield S A, Kollars N M, Byers J E, et al. Invasion of novel habitats uncouples haplo-diplontic life cycles[J]. Molecular Ecology, 2016, 25(16): 3801- 3816.
[20] Yeh F C, Yang R C, Boyle T. Popgene Version 1.31 Quick User Guide[M]. Canada: University of Alberta and Centre for International Forestry Research, 1999.
[21] Peakall R. Smouse P E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update[J]. Bioinformatics, 2012, 28: 2537- 2539.
[22] Takezaki N, Nei M, Tamura K. POPTREE2: Software for constructing population trees from allele frequency data and computing other population statistics with Windows interface[J]. Molecular Biology and Evolution, 2010, 27: 747-752.
[23] Pritchard J K, Stephens M, Donnely P. Inference of population structure using multilocus genotype data[J]. Genetics, 2000, 155: 945-959.
[24] Excoffier L, Laval G, Schneider S. Arlequin (ver. 3.0): An integrated software package for population genetics data analysis[J]. Evolutionary Bioinformatics online, 2005, 1: 47-50.
[25] Thornber C S, Gaines S D. Population demographics in species with biphasic life cycles[J]. Ecology, 2004, 85(6): 1661-1674.
[26] Dyck J L, Dewreede R E. Reproduction and survival in(Gigartinales, Rhodophyta)[J]. Phycologia, 2006, 45(3): 302-310.
[27] Destombe C, Godin J, Remy J M. Viability and dissemination of spermatia of(Gracilariales, Rhodophyta)[J]. Hydrobiologia, 1990, 204/205: 219-233.
[28] 任国忠, 陈美琴. 江蓠主枝不同部位的生长观察[J]. 海洋科学, 1987, 11(3): 39-42.Ren Guozhong, Chen Meiqin. Observations on the growth of different part of plant segment ofa sp. nov.[J]. Marine Sciences, 1987, 11(3): 39-42.
[29] Zardi G I, Nicastro K R, Serrão E A, et al. Closer to the rear edge: eco-logical and genetic diversity down the core-edge gradient of a marine macroalga[J]. Ecosphere, 2015, 6: 1-25.
[30] Krueger-Hadfield S A, Roze D, Mauger S, et al. Intergametophytic selfing and microgeographic genetic structure shape populations of the intertidal red seaweed[J]. Molecular Ecology, 2013, 22: 3242-3260.
[31] 黄玥. 末次冰期以来南海及日本海硅藻及其古环境变化[D]. 上海: 华东师范大学, 2009. Huang Yue. Diatom response to changes in palaeoenvironments of the South China Sea and the East Sea (Sea of Japan) since the Last Glacial Maximum[D]. Shanghai: East China Normal University, 2009.
[32] Yokoya N S, Kakita H, Obika H, et al. Effects of environmental factors and plant growth regulators on growth of the red algafrom Shikoku island, Japan[J]. Hydrobiologia, 1999, 398: 339-347.
[33] Kostamo K, Korpelainen H, Olsson S. Comparative study on the population genetics of the red algaeoccupying different salinity conditions[J]. Marine Biology, 2012(3), 159: 561-571.
[34] Ogawa T, Ohki K, Kamiya M. High heterozygosity and phenotypic variation of zoids in apomictic(Ulvophyceae) from brackish environments[J]. Aquatic Botany, 2014, 120: 185-192.
[35] 刘若愚. 中国近海重要生态建群红藻真江蓠的群体遗传多样性[D]. 青岛: 中国科学院研究院海洋研究所, 2016.Liu Ruoyu. Genetic diversity of the habitat-forming red algaalong Chinese coasts[D]. Qingdao: Institute of Oceanology, Chinese Academy of Sciences, 2016.
[36] Freshwater D W, Montgomery F, Greene J K, et al. Distribution and identification of an invasivespecies that is hampering commercial fishing operations in southeastern North Carolina, USA[J]. Biological Invasions, 2006, 8(4): 631-637.
Population genetic diversity of the red alga(Gigartinales, Rhodophyta) in the Yellow-Bohai Sea
ZHONG Kai-le1, 2, 3, SONG Xiao-han1, 2, 3, DUAN De-lin1, 2, HU Zi-min1, 2
(1. Key Laboratory of Experimental Marine Biology, Center for Ocean Mega-Science, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China)
; microsatellite; genetic diversity; gene flow; Yellow-Bohai Sea
is one of the most economically important red algae native to the northwest Pacific. We surveyed the population genetic diversity and the structure ofin the Yellow-Bohai Seausing 10 microsatellites. These microsatellites detected 65 alleles among 12 populations. The number of alleles (a) varied from 1 to 28, and the number of effective alleles (e) varied from 1.0 to 9.6. The average number of alleles (a), the average number of effective alleles (e), the average Shannon’s diversity index (), the average observed heterozygosity (o), and the average expected heterozygosity (e) were 2.4, 1.6, 0.419, 0.133, and 0.227, respectively, indicating a low population genetic diversity. There was a high genetic differentiation among the 12populations (st= 0.398 7) and a limited gene flow (m= 0.377 1). The average inbreeding coefficient was positive (is= 0.391 3,it= 0.634 0), which indicates inbreeding and heterozygotic deletion. Structure analysis and UPGMA tree analysis consistently divided the 12 populations into two genetic groups, and obvious genetic mixing was found in the Heishijiao, Dalian and Shi Island, Weihai populations. Analysis of molecular variation showed that the genetic variation in the Yellow-Bohai Sea mainly occurred at the intra-population level (73.27%). This study may provide a scientific basis for the conservation of natural resources and management ofin the Yellow-Bohai Sea.
the National Natural Science Foundation of China, No. 41761144057, No. 31971395]
Mar. 28, 2020
Q347
A
1000-3096(2020)12-0023-09
10.11759/hykx20200328001
2020-03-28;
2020-04-20
国家自然科学基金项目(41761144057, 31971395)
钟凯乐(1995-),女,汉族,山东德州人,硕士,研究方向为海藻系统进化与多样性,E-mail:18865550027@163.com;胡自民,通信作者,男,博士,研究员,研究方向为海藻系统进化与多样性,E-mail: huzm@qdio.ac.cn.
(本文编辑: 杨 悦)
猜你喜欢
中国环境科学(2023年9期)2023-09-23 12:28:40
特产研究(2022年6期)2023-01-17 05:05:06
种子(2021年3期)2021-04-12 01:42:22
农产品市场周刊(2018年35期)2018-09-13 12:43:24
农产品市场周刊(2017年27期)2017-08-09 18:50:00
四川动物(2017年4期)2017-07-31 23:54:19
科技创新导报(2016年23期)2016-12-23 16:03:35
外语教学理论与实践(2016年1期)2016-06-11 05:51:48
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:37
河北遥感(2014年3期)2014-07-10 13:16:48
