
Nrf2调控自噬对于宫颈癌发生发展的作用及机制研究
2020-12-31 07:46:02李晓兰赵骏达马俊旗
转化医学杂志 2020年6期
李晓兰,赵骏达,马俊旗
宫颈癌是女性常见的恶性肿瘤之一,其发病率和病死率居高不下,各病理类型以宫颈鳞状细胞癌最多见。其发病机制尚不明确,随着研究的深入发现,早期识别、干预宫颈癌对于其预后及治愈均有重要的临床意义[1]。而核因子E2相关因子2(nuclear factor-erythroid 2-related factor 2,Nrf2)蛋白的异常表达与自噬的相互作用可能在宫颈癌的发生发展过程中扮演着重要角色。
1 材料与方法
1.1 研究对象 宫颈癌标本及其癌旁组织各50例均来自于2018年1月-12月在新疆医科大学第一附属医院妇产科就诊的宫颈癌患者,所有的标本经两位病理医师证实,均为宫颈鳞状细胞癌,患者术前均未接受放、化疗及免疫治疗。Beclin-1、LC3、p62、Nrf2均为兔抗人多克隆抗体,购自美国ABGENT公司;DAB显色试剂盒、SP免疫组织化学试剂盒(PV-9000)购自上海碧云天生物技术有限公司。人宫颈癌SiHa细胞株购自武汉普诺赛生命科技有限公司,根据医学伦理规范,使用慢性宫颈炎患者切除的新鲜宫颈组织作为正常对照。所有患者知情同意,并获得本院伦理委员会批准。
1.2 方法
1.2.1 宫颈癌SiHa细胞细胞培养 宫颈癌SiHa细胞按常规细胞培养方式培养,将细胞接种于含10%胎牛血清、100 U/mL青霉素、100 mg/mL链霉素的DMEM培养基中,培养基置于37 ℃、5%CO2的细胞培养箱中。消化传代使用含有EDTA的0.25%胰酶,选取对数生长期细胞进行实验。
1.2.2 基因转染 取对数生长期的宫颈癌SiHa细胞经胰酶消化后,以1×105/mL密度接种于6孔板中,待细胞生长至80%融合时,取2 μg质粒加入到200 μL基础培养基中轻轻摇匀后在室温下放置5 min;取10 μL脂质体加入到200 μL基础培养基中轻轻摇匀后再在室温下放置5 min;然后混合稀释的脂质体和质粒轻轻摇匀,室温孵育30 min,最后将混合液轻轻加入到待转染的细胞中,4~6 h后换液。
1.2.3 免疫组化 采用免疫组织化学超敏两步法(SP法)染色,一抗均采用工作液(Beclin1、p62为1∶100稀释;LC3为1∶50稀释),PBS为对照,操作步骤严格按照说明书进行,经SP法染色,DAB显色,苏木素复染、脱水、透明、封片。
1.2.4 Western Blot检测蛋白表达量 在宫颈癌SiHa细胞株内分别转染Nrf2 siRNA和p62 siRNA 48 h后处理细胞,提取总蛋白,4 ℃离心25 min后,用BCA测定蛋白含量,SDS-PAGE分离样品并转膜,4%BSA封闭1 h后分别加入Nrf2、p62、Bcl2、Bax抗体,抗体浓度均为(1∶1 000),4 ℃过夜后,二抗(1∶2 000)4 ℃孵育1 h,ECL发光显影。
1.2.5 MTT法检测细胞增殖 在宫颈癌SiHa细胞株内分别转染Nrf2 siRNA和p62 siRNA处理细胞48 h后,继续培养,MTT法检测细胞增殖改变情况。
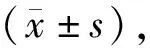
2 结果
2.1 宫颈癌组织及癌旁组织中Beclin1、LC3、p62和Nrf2的表达 免疫组化结果显示,Beclin1、LC3、p62和Nrf2的阳性染色呈棕黑色颗粒,Beclin1阳性定位于细胞膜及细胞浆,LC3阳性定位于细胞浆及部分细胞核,p62阳性定位于细胞浆,Nrf2阳性定位于细胞核。Beclin1在宫颈癌组织及癌旁组织中的阳性表达率分别为18.0%(9/50)、94.0%(47/50);LC3在宫颈癌组织及癌旁组织中的阳性表达率分别为20.0%(10/50)、92.0%(46/50);p62在宫颈癌组织及癌旁组织中的阳性表达率分别为78.0%(39/50)、14.0%(7/50);Nrf2在宫颈癌组织及癌旁组织中的阳性表达率分别为86.0%(43/50)、22.0%(11/50)。4种蛋白在宫颈癌组织及癌旁组织中表达水平均有统计学意义(P<0.05,表1),提示在宫颈癌细胞内Nrf2和p62蛋白显著高表达,自噬表达水平较低。
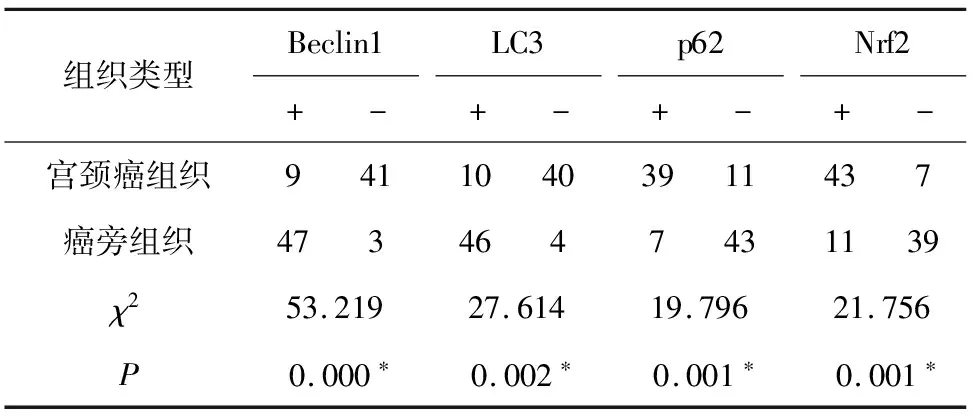
表1 宫颈癌组织及癌旁组织中Beclin1、LC3、p62和Nrf2的表达组织
2.2 在宫颈癌SiHa细胞株内分别转染Nrf2 siRNA和p62 siRNA后细胞凋亡改变情况 在宫颈癌细胞内,Nrf2与p62均呈现高表达状态(P<0.05,图1);在宫颈癌SiHa细胞株内转染Nrf2 siRNA及其阴性对照物后,细胞内p62表达显著下降,且抗凋亡蛋白Bcl2显著下降,凋亡蛋白Bax表达上升,差异有统计学意义(P<0.05);在宫颈癌SiHa细胞株内转染p62 siRNA及其阴性对照物后,细胞内Nrf2表达显著下降,且抗凋亡蛋白Bcl2表达显著下降,凋亡蛋白Bax表达上升,差异有统计学意义(P<0.05,图2);提示在宫颈癌SiHa细胞内,p62与Nrf2可成环调控,且两者相互作用可显著促进细胞凋亡。

图1 宫颈癌组织及癌旁组织中Beclin1、LC3、p62和Nrf2的免疫组化Beclin1蛋白在宫颈癌组织和癌旁组织中的表达(×400)A:癌组织;B:癌旁组织;LC3蛋白在宫颈癌组织和癌旁组织中的表达(×400)C:癌组织;D:癌旁组织;p62蛋白在宫颈癌组织和癌旁组织中的表达(×400)E:癌组织;F:癌旁组织;Nrf2蛋白在宫颈癌组织和癌旁组织中的表达(×400)G:癌组织;H:癌旁组织
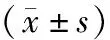
表2 转染后宫颈癌SiHa细胞株Nrf2、p62基因表达

图2 宫颈癌SiHa细胞株内Nrf2和p62表达情况与阴性对照组比较,*P<0.05
2.3 在宫颈癌SiHa细胞株内分别转染Nrf2 siRNA和p62 siRNA后细胞增殖改变情况 分别在宫颈癌SiHa细胞株内转染Nrf2 siRNA及其阴性对照物、p62 siRNA及其阴性对照物后,继续培养细胞48 h和72 h,结果显示,细胞增殖在Nrf2 siRNA和p62 siRNA组显著降低,差异有统计学意义(P<0.05,图3),提示在宫颈癌SiHa细胞内,抑制Nrf2和p62的表达可以显著抑制细胞增殖,具有时间依赖效应,图4。

图3 在宫颈癌SiHa细胞株内分别转染Nrf2 siRNA和p62 siRNA后细胞凋亡改变情况

图4 宫颈癌SiHa细胞株内分别转染Nrf2 siRNA和p62 siRNA后细胞增殖改变情况
3 讨论
宫颈癌在全球各地均有发生,是人体最常见的恶性肿瘤之一,居于女性恶性肿瘤的首位。临床流行病学结果显示,从一般的宫颈癌首病变诱发为宫颈癌约10年时间,提示宫颈癌的早期发现可能使其成为一种可以预防和治愈的疾病[2-4]。因此,研究宫颈癌发生发展的潜在分子机制对于宫颈癌的治疗具有重要的意义。
自噬是一种高度保守的真核细胞内分解代谢过程,其特点是在细胞质内形成双层膜结构的自噬小体吞噬细胞内受损的细胞器[5]。自噬由压力信号-哺乳动物雷帕霉素复合物1(mammalian target of rapamycin complex 1,mTORC1)始动,mTORC1 激活激酶ULK1(ATG1),继而形成ATG13和ATG17复合物[7]。mTORC1活性受到抑制后,自噬小体开始形成,其形成还需要激活由Ⅲ型磷脂酰肌醇3-激酶与Beclin1所形成的复合物。LC3II通过ATG7和ATG3进入自噬小体膜内。LC3II蛋白还需要一些支架蛋白的协助,如p62和NIX,通过它们可以识别并结合聚集的蛋白和受损的细胞器,将其带入自噬小体内部进行降解[6-9]。自噬小体随后与溶酶体形成自噬溶酶体,自噬小体内的物质受到许多不同的溶酶体蛋白的降解,为细胞代谢和细胞器的更新提供能量和原料[10-12]。大多数的相关实验研究发现,自噬在健康细胞内可以起到抑制肿瘤的作用,而在多种肿瘤中均异常存在[13-15]。本研究发现,在宫颈癌组织内,自噬蛋白呈现低表达状态,自噬行程标志性蛋白Beclin1、LC3表达显著低于癌旁对照组组织,同时自噬降解关键蛋白p62表达异常升高,本研究结果印证了前期研究结论,进一步证实在宫颈癌的发生发展过程中,自噬确实发挥着重要作用。
氧化应激是指由于氧自由基过量生成和/或细胞内抗氧化防御系统受损,导致氧自由基及其相关代谢产物过量聚集,对细胞产生多种毒性作用的病理状态。Nrf2是外源性有毒物质和氧化应激的感受器,在参与细胞抗氧化应激和外源性有毒物质诱导的主要防御机制中发挥重要作用[16-17]。本研究结果显示,在宫颈癌组织中,Nrf2异常高表达,自噬表达水平较低,提示在宫颈癌的发生发展过程中,Nrf2与自噬的共同作用可能影响细胞的增殖与凋亡。
本研究结果显示,宫颈癌细胞SiHa转染Nrf2 siRNA及p62 siRNA后,Nrf2与p62的表达呈正相关,均显著下调细胞增殖,促进细胞凋亡。表明在宫颈癌发生发展过程中,Nrf2、p62反馈环的存在是自噬与氧化应激联系的关键,可以进一步调控宫颈癌细胞的增殖与凋亡,该研究结果与有关报道相一致。据报道p62可以通过Keap1降解而激活Nrf2信号通路发挥细胞保护作用,预防软脂酸诱导的脂中毒及防止羰基氰化物氯苯腙诱导的细胞凋亡。有研究表明,氧化应激发生时,Nrf2的过度表达可以增加p62的mRNA水平,Nrf2结合到下游抗氧化元件p62启动子区可以促进p62的生成[18-20]。提示p62与Nrf2之间可能存在正性反馈环。与本文的研究结果相一致。本研究还发现在宫颈癌SiHa细胞内,抑制Nrf2和p62的表达可以显著抑制细胞增殖,重要的是具有时间依赖效应。
本研究证实在宫颈癌细胞内,Nrf2与p62的负向相互作用可以显著促进细胞凋亡,抑制细胞增殖。下一步将对两分子的具体作用机制展开研究。
猜你喜欢
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:05:56
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:43
西南军医(2016年6期)2016-01-23 02:21:19
医学研究杂志(2015年11期)2015-06-10 06:44:03
西南军医(2015年3期)2015-04-23 07:28:32
中国当代医药(2015年16期)2015-03-01 02:03:11
中国医药导报(2015年27期)2015-02-28 22:08:02
国际心血管病杂志(2015年5期)2015-02-27 12:11:34
西南军医(2015年2期)2015-01-22 09:09:37
现代检验医学杂志(2014年6期)2014-03-03 02:14:24
