
维氏气单胞菌acrR敲除株的构建
2020-12-21 07:24:40张伊动唐燕琼
生物学杂志 2020年6期
胡 康, 张伊动, 唐燕琼, 李 宏, 马 香, 刘 柱
(海南大学 生命科学与药学院 热带生物资源教育部重点实验室, 海口 570228)
AcrR蛋白是转录抑制因子TetR家族的成员,是一种负调控因子[1]。在大肠杆菌中已经对AcrR蛋白开展了广泛研究,其N末端形成典型的螺旋-转角-螺旋结构,是其与DNA链的结合位点。被诱导时,AcrR会抑制acrAB的表达[2]。研究发现四环素-镁复合物与AcrR结合能降低AcrR与DNA结合的亲和力,并使多药物外排复合物AcrAB转录活性增强[3]。AcrAB-TolC作为大肠杆菌中最主要的多药外排泵,包括外膜通道蛋白TolC、周质融合蛋白AcrA和药物质转运子AcrB。最新研究表明,在植物乳杆菌中,转录调节因子AcrR可通过调节脂肪酸合成提高乙醇耐受性,还可通过调控醛-醇脱氢酶(AdhE)控制植物乳杆菌对甘露醇和山梨醇的利用[4-5]。在假单胞菌中,AcrR可以调节铁清除剂的合成,提高细菌在环境中的竞争力[6]。在流感嗜血杆菌中,AcrR与细菌在热应激反应中对亚胺培南抗性增强相关[7]。另外AcrR突变会导致大肠杆菌对苯酚的抗药性增强[8]。多药物外排泵能外排有机溶剂、氧化剂及细菌产生的有害代谢物,并且能外排多种抗生素[9]。研究表明acrR基因的缺失会造成细菌致病性、运动性、耐药性以及生物膜等表型变化,表明在细菌生长代谢中AcrR蛋白具有多重功能[10]。
维氏气单胞菌(Aeromonasveronii)隶属弧菌科气单胞菌属,是革兰氏阴性杆菌,其普遍存在于自然环境中[11]。作为一种新型水生动物病原菌,维氏气单胞菌主要感染诸如锦鲤、草鱼等经济养殖鱼类,可导致其大量死亡[12]。这些年有关其感染人的病例也有逐年增多的趋势,人食用了维氏气单胞菌菌株污染的水产品、蔬菜时可引发腹泻、败血症、心内膜炎等[13]。本研究以AcrAB-TolC多药外排泵为切入点,构建acrR基因敲除菌株,对其生长特性及生物膜成膜特性进行初步研究,试图解析维氏气单胞菌的致病机制及耐药机理。研究结果对于防治重要水产病害、促进水产经济发展具有重要意义。
1 材料与方法
1.1 材料
1.1.1 质粒、菌株及引物
自杀质粒pRE112以及大肠杆菌WM3064菌株(Escherichiacoli,WM3064)、维氏气单胞菌C4菌株(A.veronii, C4)均由本实验室提供。本实验用到的验证及扩增引物序列信息如表1所示,由上海生物工程股份有限公司提供测序及引物合成。

表1 本实验使用的引物序列
1.1.2 试剂盒、酶及药品
细菌基因组DNA提取试剂盒购自上海生工生物工程有限公司;2×PCR Mix、质粒提取试剂盒及DNA/胶纯化试剂盒购自南京诺唯赞生物科技有限公司;T4连接酶、T4连接酶Buffer、BstX I、KpnI-HF和SacI-HF限制性内切酶购自NEB公司;氯霉素(Cam)、氨卞青霉素(Amp)、二氨基庚二酸(Dap)及琼脂粉购自索莱宝科技有限公司;蛋白胨、酵母提取物及氯化钠均购自西陇科学股份有限公司。
1.2 方法
1.2.1acrR基因上、下游同源序列扩增
用相应试剂盒提取维氏气单胞菌C4的基因组DNA,随后以其为模板,分别用acrR F1/R1和acrR F2/R2扩增目的片段。按照基因组DNA 1 μL,2×PCR Mix 13 μL,上或下游引物各1 μL,ddH2O 10 μL的体系配制好溶液后,再进行如下PCR程序:95 ℃ 预变性 8 min;95 ℃ 变性 30 s,55 ℃ 退火 30 s,72 ℃ 延伸 60 s,25 cycles;72 ℃ 再延伸 10 min。
1.2.2 pRE112-ΔacrR载体构建
首先用质粒提取试剂盒获得自杀质粒pRE112,在37 ℃的培养箱中孵育经KpnⅠ-HF和ScaⅠ-HF内切酶处理过的酶切产物30 min,随后用纯化试剂盒回收酶切产物。在37 ℃的培养箱中孵育经BstXⅠ内切酶处理过的酶切产物30 min,再用纯化试剂盒回收酶切产物。按以下连接体系(10 μL):pRE112载体酶切产物(12 ng/μL) 5.0 μL、acrR上/下游同源臂酶切产物(60 ng/μL)各1 μL,10×T4 Buffer 1 μL,T4连接酶1 μL,ddH2O 1 μL,将质粒酶切产物与片段酶切产物在22 ℃下体外连接2 h。吸取1 μL上述连接产物电转化到E.coliWM3064感受态细胞中,在37 ℃下于含有Dap和Cam的抗性平板中培养,隔天挑菌验证阳性克隆。
1.2.3 双亲接合及acrR基因敲除菌株的筛选
将维氏气单胞菌C4于含Amp的LB液体培养基中过夜培养,并将携带有pRE112-ΔacrR质粒的大肠杆菌WM3064于含Dap和Cam的LB液体培养基中过夜培养。将过夜培养的菌分别转接培养至OD600为0.4~0.6,将维氏气单胞菌C4和大肠杆菌WM3064按1∶1、1∶3和1∶7的比例混合至终体积为800 μL。6 000 r/min离心3 min 后除去大部分上清液,将剩余菌液混匀后滴于含有DAP的LB平板上,30 ℃ 倒置培养24 h后,在平板上滴加1 mL液体LB培养基并用涂布棒将长出的菌落刮下,取100 mL菌液涂布于含有Cam和Amp的双抗平板上,待菌落长出后PCR验证接合结果。将接合成功的维氏气单胞菌C4于含有Amp的LB液体培养基中过夜培养,取100 μL涂布于含有22%蔗糖和Amp的LB固体培养基上30 ℃倒置培养16 h。菌落PCR验证敲除菌株。
1.2.4 生长曲线测定
挑取野生型维氏气单胞菌C4和ΔacrR菌株的单菌落,在含Amp的LB液体培养基中30 ℃,150 r/min振荡过夜培养,次日转接菌液于含Amp的新鲜LB培养基中至终浓度OD600为0.02,每间隔1 h取样,分别测定培养菌的OD600数值,然后绘制野生型维氏气单胞菌C4和ΔacrR菌株的生长曲线。
1.2.5 生物膜测定
在24孔聚苯乙烯培养板中每孔添加1 mL含Amp的LB液体培养基,按OD600为0.01分别接种过夜培养的野生型维氏气单胞菌C4及ΔacrR的菌液,30 ℃静置孵育24 h;将培养液吸出,每孔加入1 mL无菌PBS缓冲液冲洗;再向每孔中加入1 mL甲醇处理20 min,随后将培养孔中残余的甲醇吸出;向每孔中加入1 mL 0.5%结晶紫溶液,染色10 min;吸出培养孔中的结晶紫染色液后,用流水把残余的染料冲净;随后将培养板倒置在滤纸上以便除去残余的水分,并于37 ℃烘箱中彻底烘干水分;再向每孔加入1 mL 33%醋酸溶液,在37 ℃下反应30 min以溶解结晶紫;用酶标仪测定培养孔中溶液的OD570值。
2 结果与分析
2.1 pRE112-ΔacrR载体的构建
以野生型维氏气单胞菌C4(WT)基因组DNA为模板,PCR扩增得到acrR上、下游同源臂片段。凝胶电泳结果显示,acrR上游同源臂和下游同源臂的PCR产物均在300 bp左右,而且PCR产物条带单一、明亮(图1)。acrR上游同源臂长度为304 bp,acrR下游同源臂片段长度为311 bp,表明目的产物扩增成功。

M: DL5000 DNA Marker; 1: acrR上游同源臂; 2: acrR下游同源臂
将提取的pRE112质粒以及PCR扩增得到的acrR上、下游同源臂片段分别进行酶切,然后用T4连接酶22 ℃孵育2 h,通过电转将连接产物转入大肠杆菌WM3064感受态细胞,并涂布于氯霉素抗性板中用于筛选,次日随机挑选单菌落,然后使用载体验证引物pRE112F/R进行菌落PCR扩增验证,并用空载体pRE112为模板作为阳性对照,以筛选阳性克隆。结果显示,空载体pRE112的PCR产物长度约为500 bp, 验证片段大小约为1 100 bp (图2),说明pRE112-ΔacrR载体构建成功。

M: DL5000 DNA Marker; 1~6: pRE112-ΔacrR基因敲除质粒菌落PCR产物; 7: 阳性对照; 8: 阴性对照
2.2 双亲接合及ΔacrR菌株的筛选
将上述转入大肠杆菌WM3064中的重组载体pRE112-ΔacrR通过双亲接合实验转入到野生型维氏气单胞菌中,在含Cam和Amp的双抗平板上筛选阳性菌株,并用载体验证引物pRE112F/R进行菌落PCR扩增验证。结果显示,扩增条带大约为1 100 bp,pRE112-ΔacrR载体质粒已经成功接合至维氏气单胞菌中(图 3)。然后利用22%蔗糖压力筛选以获得阳性突变菌株。使用acrR-F0/R0引物进行菌落PCR,选取野生型维氏气单胞菌C4基因组PCR产物为阳性对照。结果显示,突变株菌落PCR扩增条带明显低于野生型维氏气单胞菌C4的扩增条带。挑取3个阳性突变株进行PCR扩增,回收PCR产物并送测序,测序结果显示ΔacrR菌株构建成功(图 4)。
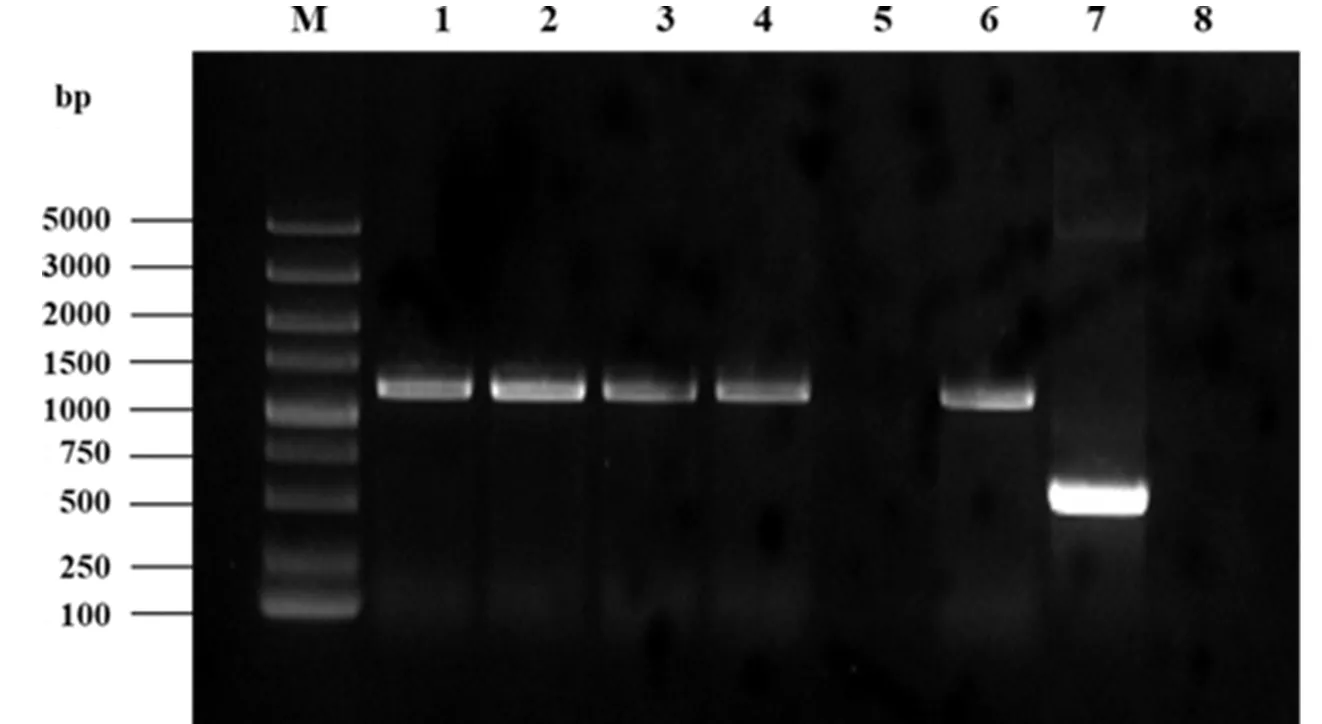
M: DL5000 DNA Marker; 1~6: PCR验证产物; 7: 阳性对照; 8: 阴性对照
2.3 ΔacrR敲除对生长的影响
为了探究acrR基因的敲除是否会影响维氏气单胞菌的生长,分别在LB培养基中培养野生型维氏气单胞菌C4和ΔacrR菌株,并每隔1 h对OD600值进行测定。结果显示ΔacrR菌株同野生型菌株生长速率一致,即acrR基因的敲除不会影响维氏气单胞菌的生长(图5)。

M: DL5000 DNA Marker; 1~6: 敲除株验证PCR产物; 7: 阳性对照; 8: 阴性对照

图5 野生型维氏气单胞菌C4及acrR敲除株生长曲线测定
2.4 ΔacrR敲除对生物膜形成的影响
为了探究acrR基因敲除对维氏气单胞菌生物膜形成的影响,将在24孔板中培养24 h后的野生型维氏气单胞菌C4及ΔacrR菌株的培养液吸出,经PBS洗涤、甲醇固定、0.5%结晶紫染色、PBS洗涤及33%醋酸染色后,用酶标仪测定培养孔中溶液的OD570值。结果显示ΔacrR菌株测出的OD570值显著高于野生型菌株,即敲除acrR基因后,细菌形成生物膜的能力显著增强(图 6)。
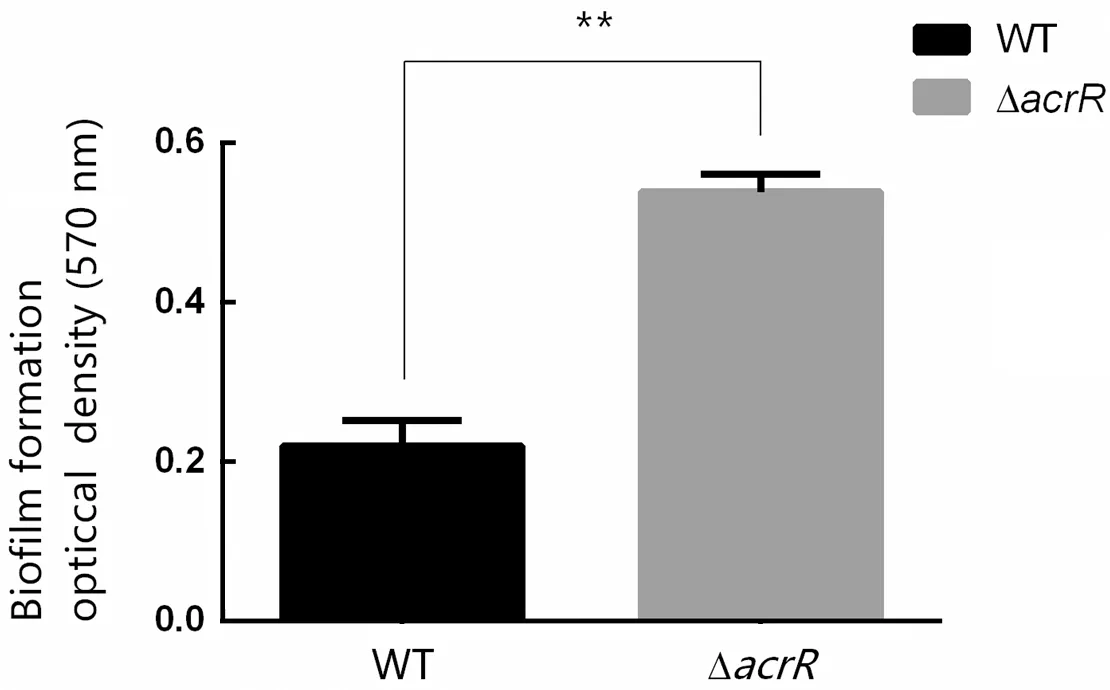
图6 野生型维氏气单胞菌C4及acrR敲除株生物膜形成测定
3 结论
本研究鉴定了AcrR蛋白对于维氏气单胞菌生长和成膜能力的影响。通过对野生型菌株及ΔacrR菌株的生长特性分析发现,AcrR蛋白的缺失并不影响维氏气单胞菌的生长,维氏气单胞菌株中敲除acrR基因导致菌株形成生物膜的能力显著增加,这一结果与医院不动杆菌acrR基因敲除株的表型一致,暗示AcrR蛋白通过调控AcrAB外排泵而参与生物膜的形成[10]。本研究结果为抵抗生物膜形成相关的病原菌提供了新的研究思路和作用靶标。
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:32
——紫 苏
河南农业(2024年1期)2024-01-19 01:56:54
华人时刊(2023年1期)2023-03-14 06:43:36
现代临床医学(2021年5期)2021-11-02 05:21:18
汉字汉语研究(2021年2期)2021-08-30 08:58:46
昆明医科大学学报(2021年4期)2021-07-23 01:22:08
洛阳师范学院学报(2017年10期)2017-03-12 08:53:58
中国兽医杂志(2016年5期)2016-06-27 01:45:02
河北书画研究(2016年3期)2016-04-28 08:55:35
中华皮肤科杂志(2014年4期)2014-12-19 12:55:59
