
老年人整体运动敏感性降低的脑灰质体积变化基础 *
2020-12-21 08:59朱子良严世振梁紫平艾克旦艾斯卡尔尹建忠姜云鹏
心理与行为研究 2020年5期
金 花 朱子良 严世振 梁紫平 艾克旦 · 艾斯卡尔 尹建忠 姜云鹏 林 琳
(1 教育部人文 社会科学重点研究 基地天津师范大学 心理与行为研究院,天津 300387)(2 天津师范大学心理 学部,天津 300387) (3 学生 心理发展与学习天 津市高校社会科学 实验室,天津 300387)(4 天津市第一 中心医院放射科,天津 300192)
1 问题提出
整体运动知觉(global motion preception, GMP)是指在一定的视觉场景中,个体将各个局部移动元素的轨迹整合成一个更有全局性的大刺激,并从中获取刺激运动速度和方向等信息(Cai, Chen,Zhou, Thompson, & Fang, 2014; Narasimhan & Giaschi,2012)。随着年龄的增长,健康成人的GMP 会逐渐衰退,即GMP 老化,表现为对整体运动的敏感性下降(Biehl, Andersen, Waiter, & Pilz, 2017; Billino &Pilz, 2019)。GMP 老化可导致老年人对动态信息的知觉障碍,引发他们和周围动态环境的交互能力的下降。
目前,已有研究证明GMP 的老化独立于视觉系统其他功能的老化。如Ball 和Sekuleryr(1986)的研究发现,老年被试的整体运动敏感性(global motion sensitivity, GMS)显著低于年轻被试,但这两组被试的视觉健康状态相同。Willis 和Anderson(2000)发现,成人区分整体运动信息的能力随年龄增长而下降,但检测静止模式的能力不受年龄影响;且老年青光眼患者、年龄匹配对照间的GMS没有显著差异。
运动信息加工主要依赖于视觉背侧通路。与观看静止刺激相比,观看运动刺激会诱发从V1(visual area 1)、V2(visual area 2)、V3(visual area 3)区经内侧颞叶V5/MT(visual area 5/ middle temporal gyrus)投射至顶叶的视觉背侧通路中许多区域更强的激活。但V5/MT、V3 区与运动信息加工的相关性更高;而V1 和V2 区虽然也对运动刺激产生反应,但它们主要应对运动刺激中蕴含的局部空间和时间结构(Grill-Spector & Malach,2004)。Braddick 和Qian(2001)发现,被试观看一致运动的随机点时,V5 和V3a 区产生更强的激活,而观看噪声运动则引起V1 区更强的激活。然而,GMP 的老化是否与脑灰质体积的退行性萎缩有关,负责GMP 的脑功能区灰质结构是否也存在类似的特异性老化,目前尚不清楚。但已有众多关于脑灰质结构老化和认知老化间关系的研究结果提示着上述特异性老化存在的可能性(Oschwald et al., 2019)。如Leong 等(2017)发现老年人海马萎缩和脑室扩张均与言语记忆和执行功能的更大下降有关;Chang 等(2015)发现老年人V3 区域面积显著小于年轻人,并且老年人V3 区域更小的面积与其在纹理辨别任务中更差的行为表现有关。
本研究拟应用多体素模式分析(multi-voxel pattern analysis, MVPA)探讨老年人GMS 下降与其脑灰质体积变化的相关性。研究先采用心理物理法获取被试的运动一致性阈值(即GMS 行为指标,下文简称“阈值”);再通过基于体素的形态学分析(voxel-based morphometry, VBM)得到灰质体积存在显著年龄效应的脑区;最后,以这些区域为兴趣区(region of interest, ROI),分析两组被试ROI 灰质体积信号形成的空间模式对个体GMS 的预测性。
结合前人关于脑老化与认知老化、GMP 神经基础的研究结果,本研究推测:(1)老年人阈值显著高于年轻人;(2)老年人视觉背侧通路特别是V5/MT区和V3 区的灰质体积显著小于年轻人,且这些区域的灰质体积信号对GMS 有预测作用;(3)结合认知与脑老化的“去分化”假设(Farràs-Permanyer et al., 2019; Seidler et al., 2010),老年人脑功能模块化程度降低,其可能需要募集更多的神经资源用于整体运动加工(Ward, Morison, Simmers, & Shahani,2018),因而老年人GMS 下降可能还与全脑范围内大量脑区灰质体积的下降有关。
2 研究方法
2.1 被试
通过广告招募到年轻组有效被试43 名(男19 名,21.85±2.01 岁)、老年组有效被试39 名(男13 名,65.20±3.84 岁),分别为天津某高校学生和天津本地居住老年人。被试入选标准:右利手,视力或矫正视力正常,无白内障等眼部疾病,无任何神经系统精神疾病症状及体征,无颅脑手术史或外伤史,无酒精滥用。老年组简易精神智能量表得分大于24 分;无MRI 检查禁忌,且脑常规MRI 检查未见因病所致的大脑结构变化。所有被试均在实验前签署知情同意书。研究获得了天津师范大学伦理委员会和天津市第一中心医院伦理委员会的批准。
2.2 程序
被试首先进行GMP 行为测试(详见2.3),并接受人口学等问卷调查,然后在确认安全后进行脑结构像扫描。
2.3 GMP 行为数据获取
通过经典的随机点阵范式获取GMP 行为数据(金花, 刘婷, 尹德圣, 邵梦灵, 朱子良, 2019)。测试程序使用基于Matlab 的PsychToolbox(http://psychtoolbox.org/)编制,刺激呈现屏的刷新率为60 Hz,分辨率为1920×1080 像素,背景为黑色(参见图1)。点阵刺激为包含100 个白色圆点(密度为0.88 点/cm2,大小为2 像素)且直径为12 cm 的白色圆形孔径;其中一定比例的信号点一致地向左或右运动,其余为噪声点,无规则地朝各个方向运动。点阵运动方向的顺序随机、概率均等。被试眼睛距屏幕60 cm,视角为5.71°。测试时先呈现注视点提醒被试实验开始,接着呈现点阵,之后呈现白色圆点(被试做出反应后持续200 ms)。要求被试尽可能准确地判断点阵的运动方向并做出相应的按键反应。
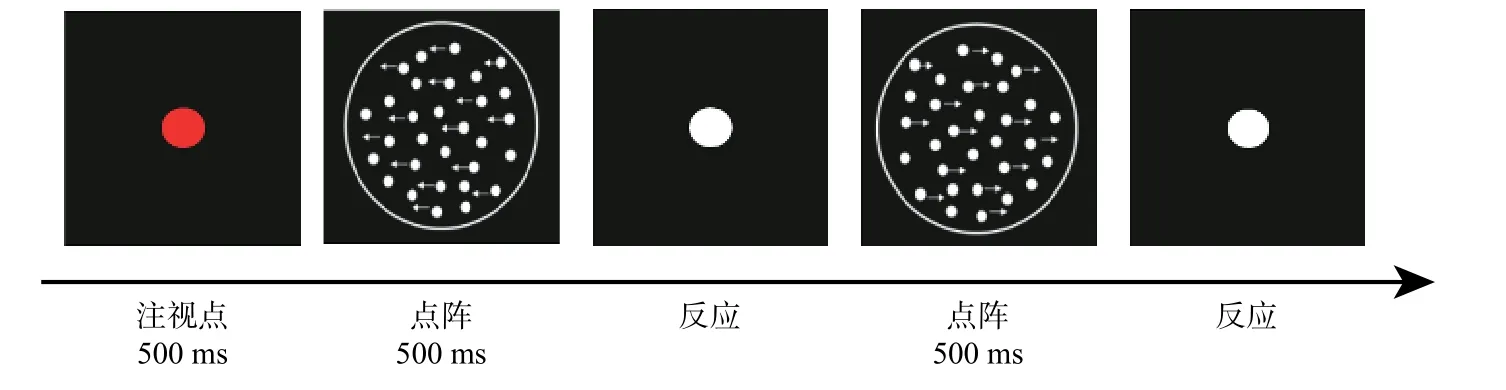
图1 整体运动知觉测试流程图
鉴于以往研究发现点速度和正确率水平在年龄相关的GMS 变化中存在不确定性(Ward et al.,2018),本研究设计了10 个水平的点速度:1°/s、1.4°/s、1.8°/s、2.4°/s、3°/s、3.8°/s、4.8°/s、6°/s、7.5°/s 和9.5°/s。采用2 下1 上(70.71%正确率)和3 下1 上(79.37%正确率)两种适应性阶梯来控制运动信号的一致性水平(Lu & Dosher, 1999)。每种条件下各完成8 次反转,后6 次反转步长为5%的数据纳入后继分析。阈值即个体判断点阵整体运动方向所需最少信号点占总点数的比例。阈值越高,GMS 越低。
测试共20 个block,约1200 个trial,分2 个session 进行。正式测试前有20 个trial 的练习让被试熟悉实验程序。被试可自主控制block 间的休息时间,整个测试约需50 分钟。
2.4 脑成像数据采集
采用西门子Prisma 3.0T 磁共振扫描仪和64通道头线圈采集T1 结构图像。扫描时被试平躺在机器中,使用软垫将被试头部固定在头线圈中以减少头部运动,并使用耳塞保护听力。T1 结构图像采集使用3D 快速磁化预备梯度回波序列(magnetization-prepared rapid acquisition gradient echo, MPRAGE),TE 为2.98 ms,反转角为7°,层厚1.0 mm,无间隔扫描,视野256 mm×256 mm,矩阵256×256,192 层。年轻组和老年组数据分别在天津师范大学脑功能成像中心和天津市第一中心医院采集,TR 分别为2530 ms 和2300 ms,成像时间分别为363s 和330s。
2.5 数据处理
2.5.1 结构像预处理
对结构像的VBM 预处理在基于Matlab 的CAT 12.6 工具包(http://www.neuro.uni-jena.de)中完成。预处理步骤包括:(1)将T1 原始数据转化成NIfTI 格式;(2)使用组织概率图(tissue probability maps, TPM)将每名被试的T1 像分割为灰质、白质、脑脊液等成分;(3)使用DARTEL方法进行空间标准化,然后进行调制;(4)检查标准化后图像的质量;(5)在spm12 中将调制后的灰质分割图进行8 mm×8 mm×8 mm 的空间平滑,用于接下来的统计分析。
2.5.2 灰质体积信号组间差异及对行为表现预测性的MVPA 分析
在spm12(http://www.fil.ion.ucl.ac.uk/spm/)中进行组水平的统计分析,把可能干扰结果的全脑体积(total intracranial volume, TIV)和性别作为协变量,进行独立样本t检验,显著水平定义为体素水平上的FWE 矫正后p<0.05,体素数量k>100。比较两组在灰质体积上的差异,并将老年人在灰质体积上显著小于年轻人的团块体积信号提取出来进行MVPA 分析。
官方机构的泄题是否涉及进一步的利益纠葛?这个问题我们不得而知,在事件未明朗前也不便展开讨论。但来自培训机构本身的人员的专业意识不到位,让学员蒙受不白之冤,这样的事确有发生,理应引起关注。
在The Decoding Toolbox(https://sites.google.com/site/tdtdecodingtoolbox/)中进行回归任务,即分别使用ROI 的灰质体积信号模式来预测被试的GMS。机器学习算法为支持向量回归(support vector regression, SVR),线性内核,惩罚系数c=1,损失函数中的epsil=p=0.1。使用留一法(leave-one-run-out)在该回归任务中进行交叉验证(cross-validation)。将预测得到的行为数据与实际的行为数据进行皮尔逊积差相关分析,该相关系数即为预测准确率,对相关系数进行1000 次置换检验,显著性水平定义为p<0.05。
3 结果
3.1 行为结果
取被试全部速度条件和正确率条件下的阈值均值作为该被试的阈值,进行后续分析。独立样本t检验显示两组阈值差异显著[t(80)=3.93,p<0.001, Cohen’sd=0.87],老年人阈值(43.77±21.04)显著高于年轻人(28.57±13.48),且年龄与阈值显著正相关(r=0.40,p<0.001),说明GMP 存在老化现象。其他人口学变量分析显示,男性的阈值显著低于女性[t(80)=2.03,p=0.046, Cohen’sd=0.46],提示有必要在后继分析中将性别作为协变量。
3.2 灰质体积信号组间差异及其对阈值预测性的MVPA 分析结果
老年人TIV 显著小于年轻人[t(80)=3.33,p=0.001, Cohen’sd=0.74],TIV 与阈值显著负相关(r=-0.22,p=0.04),提示有必要在后继分析中将TIV 作为协变量。
首先,对两组灰质体积进行全脑比较。在控制性别和TIV 后,发现右颞中回(V5/MT)、右枕中回(V3)、左舌回(V3)等14 个区域存在显著的组间差异(参见表1 和图2),老年人在上述脑区的灰质体积显著小于年轻人。MVPA 发现,分别使用右中央前回(r=0.27,p=0.013)、右顶下缘角回(r=0.23,p=0.034)、右颞中回(V5/MT,r=0.47,p<0.001)、右角回(r=0.32,p=0.003)、右梭状回(r=0.27,p=0.013)、右枕中回(V3,r=0.35,p=0.001)和右小脑前叶(r=0.23,p=0.036),左舌回(V3,r=0.37,p<0.001)和左颞下回(r=0.31,p=0.005)区域灰质体积信号得到的预测值与实际阈值显著正相关,但仅左舌回(V3)的结果通过了置换检验(参见图3A 和3B)。最后,将左舌回的灰质体积与阈值进行相关分析,发现该区域灰质体积与阈值显著负相关(r=-0.38,p<0.001),表明该区灰质体积下降可以有效预测个体阈值的升高(参见图3C)。
在上述MVPA 结果中,没有观察到V5/MT 及右侧V3 区对阈值的预测性,这可能与它们被检测到年龄效应的体素数量较少有关(分别为196 和148)。体素数量太少导致用于训练分类器的特征较少,使得预测准确率不高。因此,参考多数研究的方法,本研究直接将双侧V3 和V5/MT 作为ROI 进行MVPA 分析(V3 和V5/MT 分别定义为Brodmann 模板的19 和21 区)。结果显示,通过右V5/MT(r=0.28,p=0.01)、左V3(r=0.49,p<0.001)区灰质体积信号预测的阈值与实际阈值显著正相关(参见图4),而右V3 区和左V5/MT区则没有显著的预测作用。将右V5/MT、左V3 的灰质体积提取出来进行独立样本t检验,发现老年人这两个区域的体积显著小于年轻人[右V5/MT:t(80)=7.73,p<0.001, Cohen’sd=1.71; 左V3:t(80)=6.46,p<0.001, Cohen’sd=1.43]。为了进一步说明这些区域体积信号改变与个体阈值之间关系的方向,本研究将被试性别和TIV 作为控制变量,分别将这两个区域的灰质体积与阈值做偏相关分析,结果显示这两个区域的体积均与阈值呈显著负相关(右V5/MT:r=-0.38,p<0.001; 左V3:r=-0.30,p=0.007),表明老年人GMS 降低与这些区域的灰质体积减少有关。
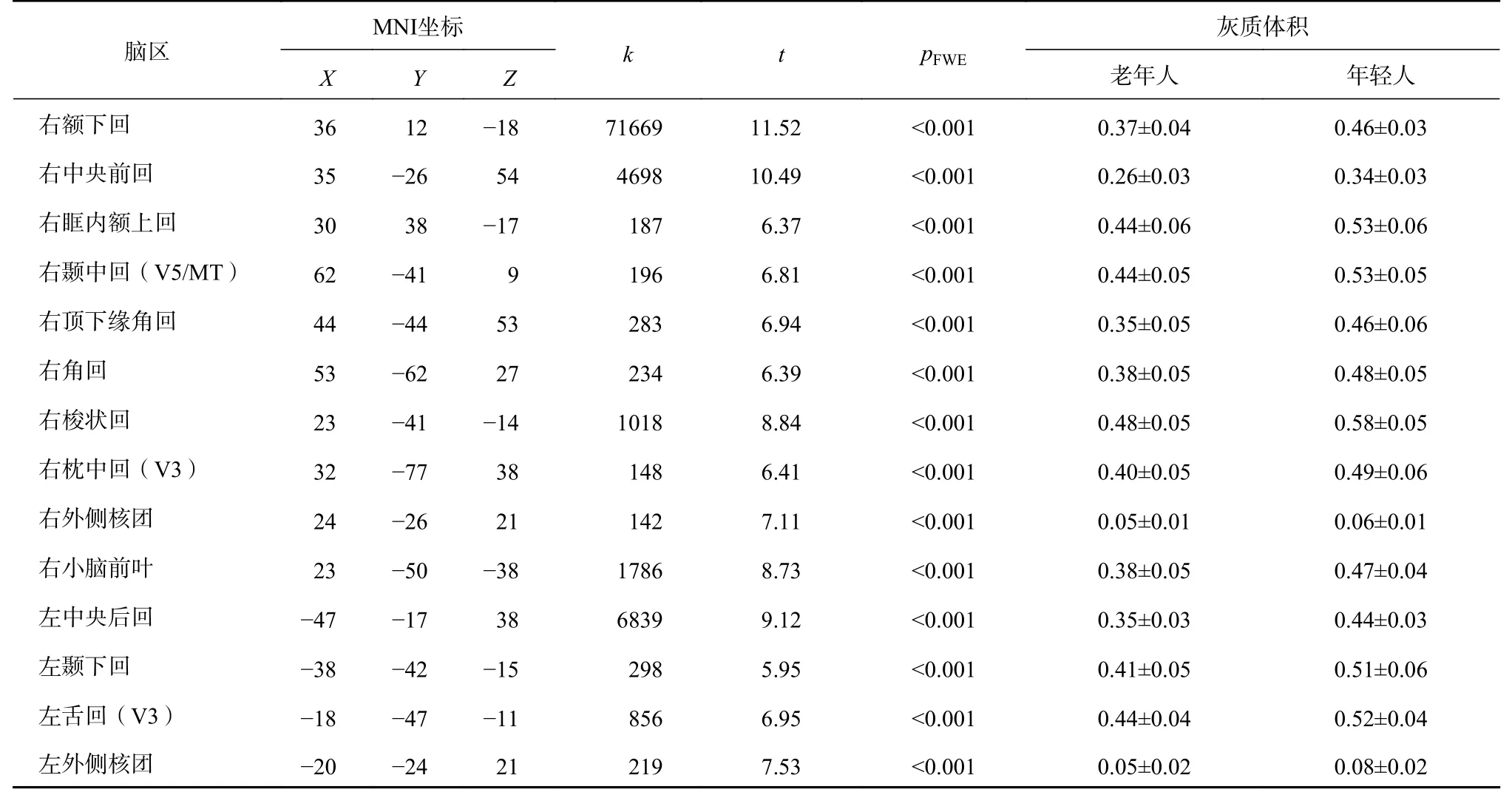
表1 老年人在灰质体积上显著小于年轻人的脑区

图2 老年人灰质体积显著小于年轻人脑区的表面映射图
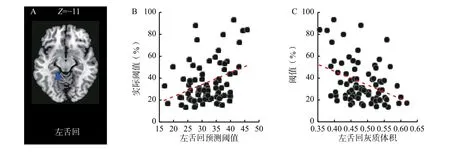
图3 左舌回(V3)灰质体积信号对阈值的预测作用(A)左舌回水平面图(B)左舌回灰质体积信号预测的阈值与实际阈值的相关(C)左舌回灰质体积与阈值的相关
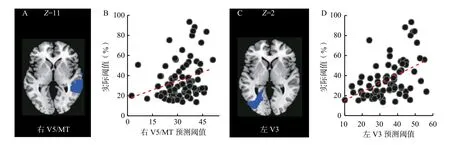
图4 右V5/MT 和左V3 灰质体积信号对阈值的预测作用(A)右V5/MT 水平面图(B)右V5/MT 灰质体积信号预测的阈值与实际阈值的相关(C)左V3 水平面图(D)左V3 灰质体积信号预测的阈值与实际阈值的相关
4 讨论
本研究运用多变量方法探讨了GMP 老化相关的灰质体积变化。行为结果验证了GMP 的老化现象,即老年人的阈值显著高于年轻人,老年人的GMS低于年轻人。MVPA 结果发现,右侧V5/MT 和左侧V3 区的灰质体积信号能够显著预测被试的阈值;控制性别和TIV 的偏相关分析发现,这些区域的灰质体积与阈值负相关;且老年人这些区域的灰质体积显著小于年轻人。另外,研究也发现了老年人视觉背侧通路外更大范围内灰质体积的下降,包括右中央前回、右梭状回、右小脑前叶、右角回和左颞下回等,它们的灰质体积信号对被试阈值具有预测性,但没有通过置换检验。结果提示,V5/MT 和V3 区,特别是V3 区的灰质体积下降可能是老年人GMS 下降的主要原因;而全脑范围内其余脑区的灰质体积下降可能也在一定程度上与老年人GMS 下降有关。
对于V5/MT 区在GMP 中的核心作用,以猴和人为对象的研究均已得到一致的肯定结果(Braddick &Qian, 2001; Cai et al., 2014; Chen, Cai, Zhou,Thompson, & Fang, 2016; Grill-Spector & Malach,2004)。Cai 等使用经颅磁刺激,以健康成人为研究对象的结果为V5/MT 区功能与GMP 之间的因果关系提供了重要的直接证据。该研究在实验中通过磁刺激短暂、可逆地改变单侧V5/MT 区域的神经活动,并在刺激前后要求被试判断随机点阵的整体运动方向,结果显示,刺激削弱了被试对整体运动方向的辨别能力。而且,GMP 老化的神经基础研究也发现,老年人的V5/MT 区在GMP 任务中的激活大于年轻人(Pilz, Miller, & Agnew, 2017;Ward et al., 2018)。本研究得到了右侧V5/MT 区域的灰质体积对个体GMS 的预测作用和老年人该脑区灰质体积显著减少的结果,提示老年人GMS 的下降可能与其右侧V5/MT 区灰质体积的减少紧密相关。研究结果从大脑灰质结构变化的角度,为V5/MT 区与GMP 老化的关联性提供了新的证据。
V3 区在GMP 中的作用存在一定的争议。Rees,Friston 和Koch(2000)应用随机点阵范式(点阵的运动一致性水平分别为0、6.25%、12.5%、25%、50%和100%),借助fMRI 技术探讨人和猴V5/MT区功能特征间的关系时发现,随机点阵方向判断任务均激活了V5/MT 和V3 区,但两个脑区的响应模式不同。V5/MT 区的响应为线性模式;而V3 区的响应则呈现为以25%为拐点的曲线模式,当点阵的一致性水平低于25%时随一致性水平增加而降低,而当点阵一致性水平高于25%时随一致性水平的增加而增加。Harvey,Braddick 和Cowey(2010)的结果却提示着V3 区参与整体运动加工的不确定性,该研究在实验中向被试呈现包含一个整体运动小点阵的大随机点阵,要求被试判断小点阵出现在屏幕上方还是下方,或小点阵是旋转还是径向运动。结果发现接受V5/MT 区经颅磁刺激的4 位被试在不同任务中均表现出了行为能力下降;但仅一位被试在接受V3 区经颅磁刺激后在全部任务中表现出了行为能力的下降。而前述Cai 等(2014)的研究结果也提示V3 区可能不参与GMP 的加工。Cai 等发现,经颅磁刺激V3a 区损害被试对局部运动而非整体运动的方向区分能力。综合上面的研究结果,本研究推测V3 区可能参与GMP 的加工,但非GMP 的必要功能区。更重要的是,Chen 等(2016)发现V3 区在功能上具有很强的可塑性。该研究发现施加于健康成人V3a 区的经颅磁刺激可使被试在100%一致运动中的行为表现(局部运动知觉)显著变差,但不影响被试在40% 一致运动中的行为表现(GMP);而刺激V5/MT 区特异性地损害了40%一致运动的加工。被试进行五天100%纯运动信号分辨任务练习后,刺激被试V3a 区不仅特异性地损害了对100%一致运动的加工,还损害了对40%一致运动的加工,而刺激V5/MT 区不再对40%一致运动的加工发生影响。V3 区在GMP 中的参与性和它的这种功能可塑性可能是其在老年人GMS 下降中起重要作用的直接原因。老年脑表现为各脑区的功能模块化程度降低,神经表征的特异性下降,刺激加工的选择性下降等(Biehl et al., 2017; Ward et al.,2018)。“去分化”假设认为,老年脑的这种变化可能反映了大脑功能去分化过程或补偿机制(Farràs-Permanyer et al., 2019; Seidler et al., 2010)。研究也发现,相比年轻人,老年人在加工整体运动信号时激活更多额外的脑区(Biehl et al., 2017;Ward et al., 2018)。Biehl 等发现,被动地观看镶嵌在噪声中的径向运动点阵时,相对于年轻人,老年人不仅在V5/MT 中有更强的激活,还额外地募集了右额下回。本研究推测,伴随V5/MT 区的老化,包含V3a 的V3 区可能因其在运动知觉中独特的功能可塑性而被优先募集于补偿V5/MT 区的老化,从而使其体积变化与老年人的GMS 下降发生更显著的相关性。
根据本研究的调查结果,目前尚没有直接探讨脑视觉区结构变化与老年人GMS 下降关系的研究。但相似主题的研究为本研究结果提供了间接的有力支撑。一方面,已有研究发现,人脑灰质整体体积随年龄的增长而下降,但不同脑区的下降速度不同(彭飞, 2013; Raz, Rodrigue, Kennedy, &Acker, 2004; Zhao et al., 2019)。如Raz 等发现外侧前额叶灰质体积表现出最强的年龄相关差异,视觉联合区的灰质体积表现出弱年龄效应,初级视皮层、扣带前部和顶下皮层没有表现出年龄效应。Zhao 等也发现,大部分脑区随年龄的增加而发生灰质体积减少、皮层厚度变薄等变化;前额叶和外侧颞叶变化最为显著,60 岁是变化加速的年龄拐点。另一方面,不少研究已证实老年人认知功能的衰退与其脑灰质形态结构改变存在相关关系(Chang et al., 2015; Leong et al., 2017; Oschwald et al., 2019)。Leong 等通过对111 名中国老年人8 年的追踪,考察了东亚人脑与认知老化的关系,发现整体认知的更快下降与大脑总体积、海马和灰质体积减少有关。Chang 等基于ROI 的单变量分析研究结果也为视知觉功能老化与脑视觉区灰质体积减少间的相关关系提供了更为直接的支撑。
本研究中,全脑分析得到的灰质体积年龄相关变化的结果符合研究假设,也符合上述认知和脑老化领域内的研究结果。本研究也发现,老年人TIV 显著小于年轻人,其额叶、颞叶、顶叶和小脑中部分脑区灰质体积也显著小于年轻人;但也没有发现初级视皮层灰质体积和年龄相关的变化。更重要的是,这些视觉背侧通路外脑区的灰质体积与阈值显著负相关。基于“去分化假设”,本研究结果提示,老年人GMS 的下降,除了与视背侧通路上的V5/MT 和V3 区的灰质体积下降有密切关系外,还可能与脑的整体老化存在一定的相关性。
本研究使用MVPA 方法,首次探索了老年人GMS 下降和其脑灰质体积下降的关系,结果从大脑灰质结构变化的角度,为V5/MT 区和V3 区与GMP 老化的关联性提供了新的证据。但本研究还存在一定的局限性。如本研究采用横断设计,在被试的年龄段选择上较为单一;今后应尝试招募不同年龄段的老年人进一步探讨老年人GMS 下降和其脑灰质体积下降的年龄相关效应,或者同步追踪老年人随着年龄增加而发生的GMS 下降和其脑灰质体积下降关系的动态性。另外,Robertson等(2014)基于ROI 的研究发现,在执行同样的随机点阵任务中,15~27 岁的自闭症患者不仅颞中回的激活小于对照组,V1 区的激活也显著小于对照组,说明自闭症患者GMP 障碍不仅与颞中回的功能异常有关,还与初级视皮层的功能有关。基于此,本研究推测,不同原因诱发的GMS 下降可能存在不同的脑灰质结构异常。今后的研究可拓展探讨其余非老化引起的GMS 下降,如阅读障碍儿童(Joo, Donnelly, & Yeatman, 2017)、自闭症患者(李开云, 陈功香, 傅小兰, 2018; van der Hallen,Manning, Evers, & Wagemans, 2019)的脑灰质 基础,比较相同行为损害在神经层面上损害的异同性,为不同GMS 障碍人群的有效康复提供线索。
5 结论
老年人GMS 显著低于年轻人,老年人V5/MT、V3 和额顶等脑区体积显著小于年轻人。V5/MT 区和V3 区,特别是V3 区的灰质体积下降可能是老年人GMS 下降的主要原因;而全脑范围内其余脑区的灰质体积下降而引发的“补偿”困难可能也在一定程度上与老年人GMS 下降有关。
致谢本研究得到了国家自然科学基金的资助(31971021)。数据的采集和分析过程得到了魏士琳、陈俊涛、阴晓娟、张羽萍、贾丽娜、赵光、王正光、章鹏、张琪涵、李骋诗、刘丽庆、汪强、邵梦灵、彭少灵、刘拓的帮助和指导,在此一并致谢;诚挚的感谢还致予参与本研究的所有被试。
猜你喜欢
艺术生活-福州大学厦门工艺美术学院学报(2022年4期)2022-09-22
心理学报(2021年8期)2021-08-11
党的生活(黑龙江)(2020年10期)2020-12-18
中学科技(2018年9期)2018-12-19
健康管理(2017年3期)2017-04-20
智富时代(2016年12期)2016-12-01
智富时代(2016年12期)2016-12-01
Coco薇(2016年2期)2016-03-22
Coco薇(2015年3期)2015-12-24
中国新闻周刊(2004年28期)2004-08-14
