
石山苣苔属四种(含一变种)植物的染色体数目和倍性研究
2020-12-14 03:59覃信梅盘波卢永彬沈利娜张强梁燕妮
广西植物 2020年10期
覃信梅 盘波 卢永彬 沈利娜 张强 梁燕妮
摘 要: 石山苣苔屬(苦苣苔科)约41种,主要分布于我国西南石灰岩地区。到目前为止,仅其中四种的染色体数目被研究和报道,其余绝大多数物种的染色体数目和倍性尚不清楚,染色体数目和倍性在该属及其姐妹属报春苣苔属中的演变历史及其对两属物种多样性分化的影响亦不清楚。该文以叶片水培生根法获取的四种(含一变种)石山苣苔属植物 [即石山苣苔原变种(Petrocodon dealbatus var. dealbatus)、齿缘石山苣苔(Petrocodon dealbatus var. denticulatus)、弄岗石山苣苔(Petrocodon longangensis)、石山苣苔未定名种(Petrocodon sp.)]的根尖细胞为材料开展染色体实验,探索了多种不同的实验条件对染色体制片效果的影响并获取染色体数目,在石山苣苔属和报春苣苔属的系统树上追踪了染色体数目和倍性的演变历史,同时探讨染色体数目尤其是倍性变化是否对两属物种多样性分化存在影响。结果表明:(1)长度为1~1.5 cm的根尖,0.002 mol·L-1 8-羟基喹啉溶液预处理5 h,解离4 min为较适宜的染色体制备条件。(2)四种(含一变种)石山苣苔属植物染色体数目一致,均为二倍体(2n=2x=36)。(3)两属之间及两属各自的最近共同祖先染色体数目尚不能确定,除个别物种染色体条数或倍性有变化以外,其余已知染色体数目的物种均为2n=2x=36,在两属中高度一致,石山苣苔属与报春苣苔属物种多样性分化尤其两属物种多样性巨大差异与染色体数目和基因组倍性变化无关。综上结果为石山苣苔属植物及其近缘类群染色体制备提供了参考,也为进一步对该类群的分类、系统演化和物种形成等方面的研究提供了基础数据和启示。
关键词: 石山苣苔属, 染色体数目, 基因组倍性, 物种分化
中图分类号: Q949
文献标识码: A
文章编号: 1000-3142(2020)10-1466-11
Chromosome numbers and ploidy of four species (including one variety) in Petrocodon Hance
QIN Xinmei1, PAN Bo1, LU Yongbin1, SHEN Lina3, ZHANG Qiang1, LIANG Yanni2*
( 1. Guangxi Key Laboratory of Plant Conservation and Restoration Ecology in Karst Terrain, Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, Guangxi, China; 2. Wuzhou University, Wuzhou 543000, Guangxi, China; 3. Key Laboratory of Karst Dynamics, MNR, Key Laboratory of Karst Ecosystem and Treatment of Rocky Desertification, MNR, Institute of Karst Geology, Chinese Academy of Geological Sciences, Guilin 541004, Guangxi, China )
Abstract: Petrocodon Hance (Gesneriaceae) consists of 41 species and is mainly distributed in the limestone region of South China. So far, the chromosome numbers of only four species have been reported, with the rest species in the genus remained uninvestigated yet. Furthermore, the evolutionary history of the chromosome number and ploidy in Petrocodon as well as in its sister genus Primulina remain unexplored, and particularly whether it affected their diversifications into extant species diversity remain unknown. In this study, the root tip cells of four species (including one variety) of Petrocodon (i.e. Petrocodon dealbatus var. dealbatus, Petrocodon dealbatus var. denticulatus, Petrocodon longangensis and Petrocodon sp.) which were generated by hydroponic culture method for the cutting leaves, were used for the chromosome experiments. First, the effects of various experimental treating conditions on the quality of chromosome squashing were explored and the chromosome numbers were counted. Then, the evolutionary history of chromosome numbers in Petrocodon and Primulina were traced based on the molecular phylogenetic tree, and whether the diversifications of the species diversity for the two genera were impacted by changes of the chromosome number or especially the ploidy or not was discussed. The results were as follows: (1) The root tip of 1-1.5 cm length, pretreatment for 5 h with 0.002 mol·L-1 8-hydroxyquinoline solution and dissociation for 4 min was a suitable chromosome preparation condition. (2) All the four species (including one variety) investigated were diploid and the chromosome numbers were the same (2n=2x=36). (3) Although the reconstruction of the ancestral state of the chromosome number of Petrocodon and Primulina failed to give any resolution for the most recent common ancestors of the two genera (generic stem node) and each of the genera (generic crown nodes), the chromosome number of Petrocodon and Primulina was highly conserved with most taxa possessing 2n=2x=36, except for the ploidy or chromosome number changes in few species, suggesting the disproportional diversification of species diversity between Petrocodon and Primulina was not correlated to the change of the chromosome number or ploidy. This study provides a reference for the chromosome preparation of Petrocodon and its allies, and also provides basic data and implications for further research on the classification, systematic evolution and speciation of Petrocodon.
Key words: Petrocodon, chromosome numbers, genome ploidy, species diversification
石山苣苔属(Petrocodon Hance)于1883年建立,隶属于苦苣苔科(Gesneriaceae)长蒴苣苔族(Trib. Didymocarpeae Endl.)。该属早期仅包含1种和1变种,即石山苣苔(Petrocodon dealbatus)和齿缘石山苣苔(Petrocodon dealbatus var. denticulatus)(Wang et al.,1998;李振宇和王印政,2005;韦毅刚等,2010)。近年来,通过分子系统发育研究,结合进一步的形态特征比较,对石山苣苔属进行了分类修订:原石山苣苔属(Petrocodon Hance)、朱红苣苔属(Calcareoboea Wu ex Li)、方鼎苣苔属(Paralagarosolen Wei)、世纬苣苔属(Tengia Chun)、长檐苣苔属(Dolicholoma Fang & Wang)和细筒苣苔属(Lagarosolen Wang)的所有种,以及长蒴苣苔属(Didymocarpus Wallich) [柔毛长蒴苣苔(D. mollifolius)、绵毛长蒴苣苔(D. niveolanosus)和东南长蒴苣苔(D. hancei)]、文采苣苔属(Wentsaiboea Fang & Qin) [天等文采苣苔(W. tiandengensis)]和报春苣苔属(Primulina Hance) [广西报春苣苔(Primulina guangxiensis)]的部分种类都被并入广义石山苣苔属内(Weber et al.,2011;Xu et al.,2014)。这一分类修订,加之随后该属一些新物种的不断发表 (Chen et al.,2014;Hong et al.,2014;Xu et al.,2014;Yu et al.,2015;Li & Wang,2015;Middleton et al.,2015;Guo et al.,2016;Lu et al.,2017;Cen et al.,2017;Zhang et al.,2018;陈力等,2019;Zhang et al.,2019;Li et al.,2019;苏兰英等,2019),使得现今广义石山苣苔 属增加,约41种(含1变种)(http://www.IPNI.org)。
广义石山苣苔属因并入了多个其他属的部分或全部种类而使其形态尤其花部形态复杂多样,可能是旧世界苦苣苔科植物花形态最为复杂多样的属之一(Weber et al.,2011)。卢永彬等(2017)对广义石山苣苔属的形态演化研究表明,多数花部形态特征尤其以往属的分类界定特征,在演化过程中变化频繁且发生了高度同塑性演化(homoplastic evolution),是导致传统形态分类不自然的关键因素。虽然新界定的石山苣苔属在分子系统树上为单系类群,但是它的一些物种反而在形态上与其姐妹属报春苣苔属的一些物种最为相似(如Petrocodon tiandenensis与Primulina renifolia;Petrocodon guangxiensis与Primulina tabacum)。目前,还未能找到石山苣苔属与其姐妹属报春苣苔属各自的共衍征(synapomorphy)能够用于确凿地区分此两属。另外,广义石山苣苔属约41种,而作为它的姐妹属报春苣苔属有约233种(http://www.IPNI.org),导致两姐妹属间物种多样性极不对称的原因亦缺乏研究。这些问题的回答,不仅要进一步对广义石山苣苔属植物形态、分子系统发育开展更廣泛、更深入的研究,而且更需要从其他层面或角度开展研究。
染色体是遗传信息的载体,通过染色体数目、倍性和核型等指标,可分析并揭示染色体结构差异及遗传多样性,为植物的分类和系统演化提供重要依据。目前已报道了报春苣苔属百余种(约占总种数三分之二)植物的染色体数目,而石山苣苔属植物染色体数目和倍性的研究却寥寥无几,仅见Petrocodon hancei(2n=20)、Petrocodon jingxiensis、Petrocodon hechiensis和Petrocodon niveolanosus(2n=36)四种石山苣苔的染色体数目有所报道(曹丽敏等,2003;刘瑞瑞等,2014)。由此可见,石山苣苔属不仅绝大多数物种的染色体数目和倍性尚不清楚,而且染色体数目可能存在很大差别,仍需对更多石山苣苔属植物的染色体数目进行研究。与其他许多苦苣苔科植物类似,石山苣苔属为小型染色体,染色体制片不易,需探索不同的实验条件对染色体制片效果的影响(MOller & Kiehn,2004;刘瑞瑞,2013)。因此,本研究在多种不同的实验条件下对四种(含一变种)石山苣苔属植物进行染色体制片和观察,探索石山苣苔属染色体制片的适宜条件并对染色体计数,重建染色体数目和倍性在该属及其姐妹属报春苣苔属中的演变历史,以及探讨其是否与两属物种多样性分化存在关联,旨在为石山苣苔属植物及其近缘类群的染色体制备方法提供参考,更为进一步研究该类群的分类、系统演化和物种形成等提供细胞学方面的证据和启示。
1 材料与方法
1.1 材料
四种(含一变种)石山苣苔属植物(表1),栽培于广西壮族自治区中国科学院广西植物研究所种质资源圃中。
1.2 实验方法
1.2.1 石山苣苔属染色体制片条件优化
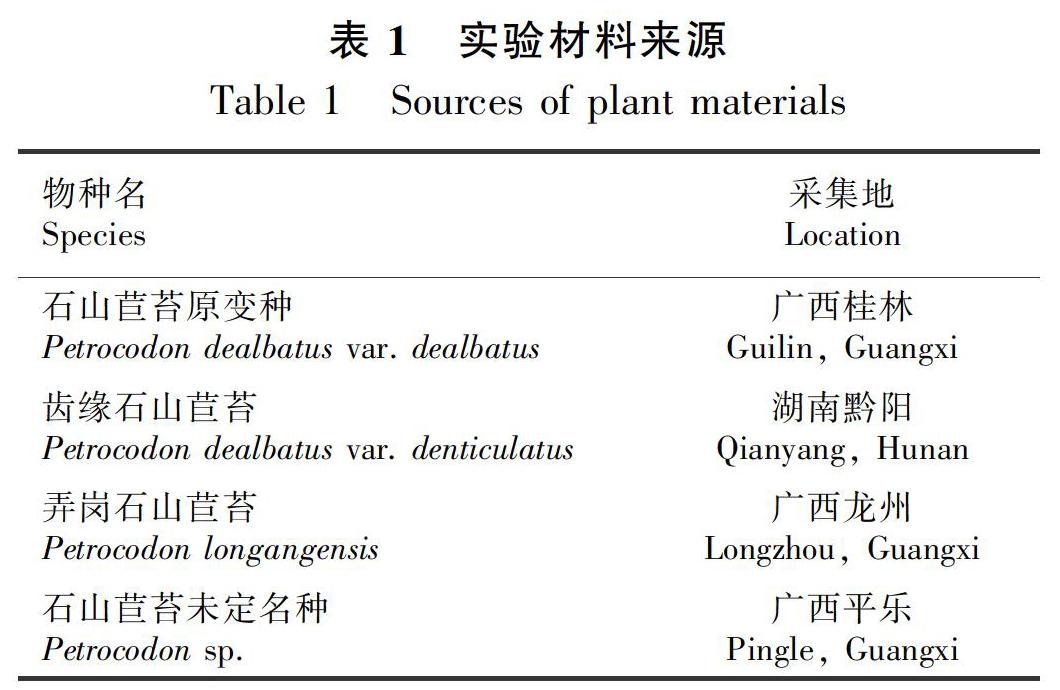
1.2.1.1 取材 取四种(含一变种)石山苣苔属植物的健康叶片,室温下用蒸馏水培养20 d左右,2~3 d换一次水,待叶片生根。对生长良好的叶片植物根尖,在8:30—9:00、20:00—20:30两个时间段内,分别对0.5、1~1.5、2~4 cm的根尖进行取材。
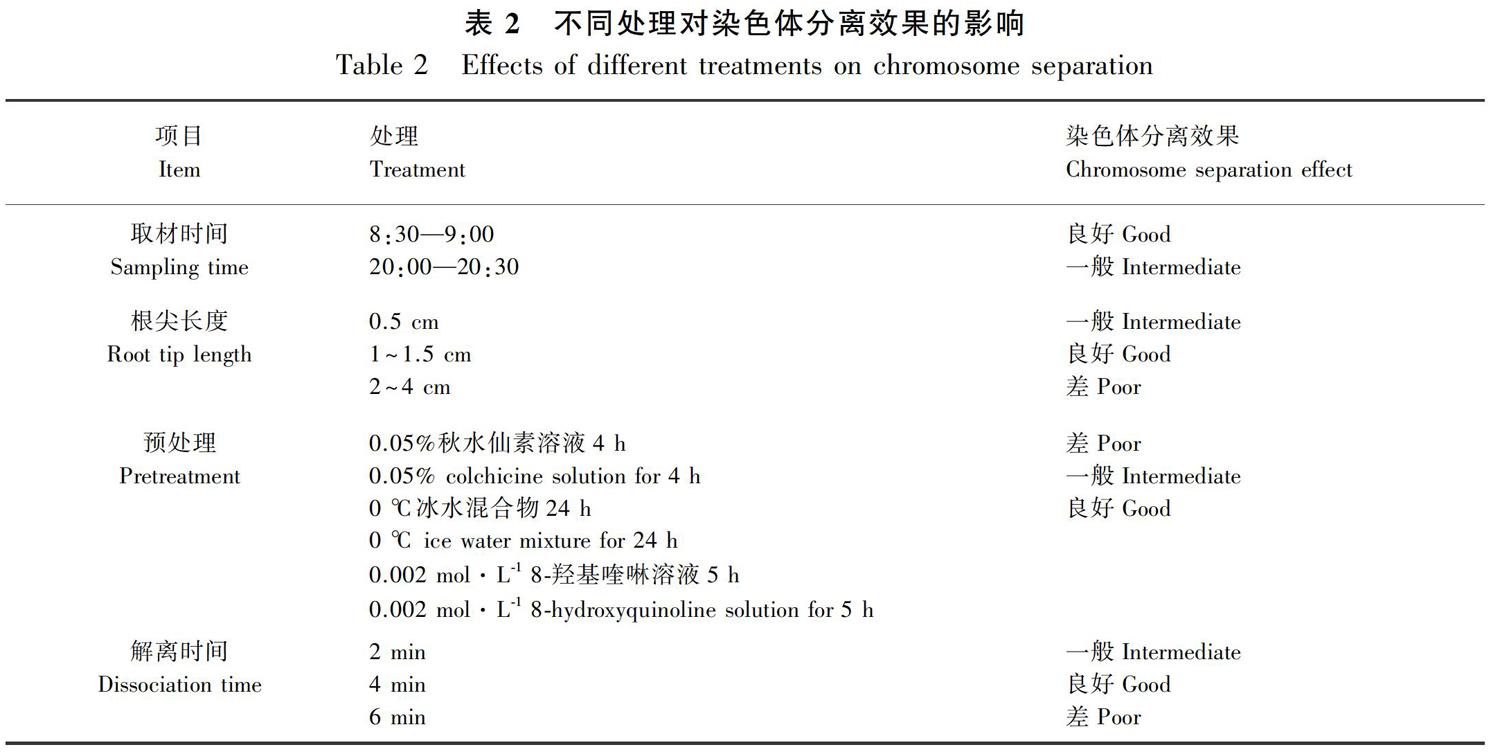
1.2.1.2 预处理 设置三种预处理方法:① 0.05%秋水仙素溶液,预处理4 h;② 0 ℃冰水混合物,预处理24 h;③ 0.002 mol·L-1 8-羟基喹啉溶液,预处理5 h。
1.2.1.3 固定 将处理后的根尖材料用蒸馏水清洗3~5次,转到卡诺固定液(乙醇∶冰醋酸=3∶1)中,在4 ℃下固定过夜。
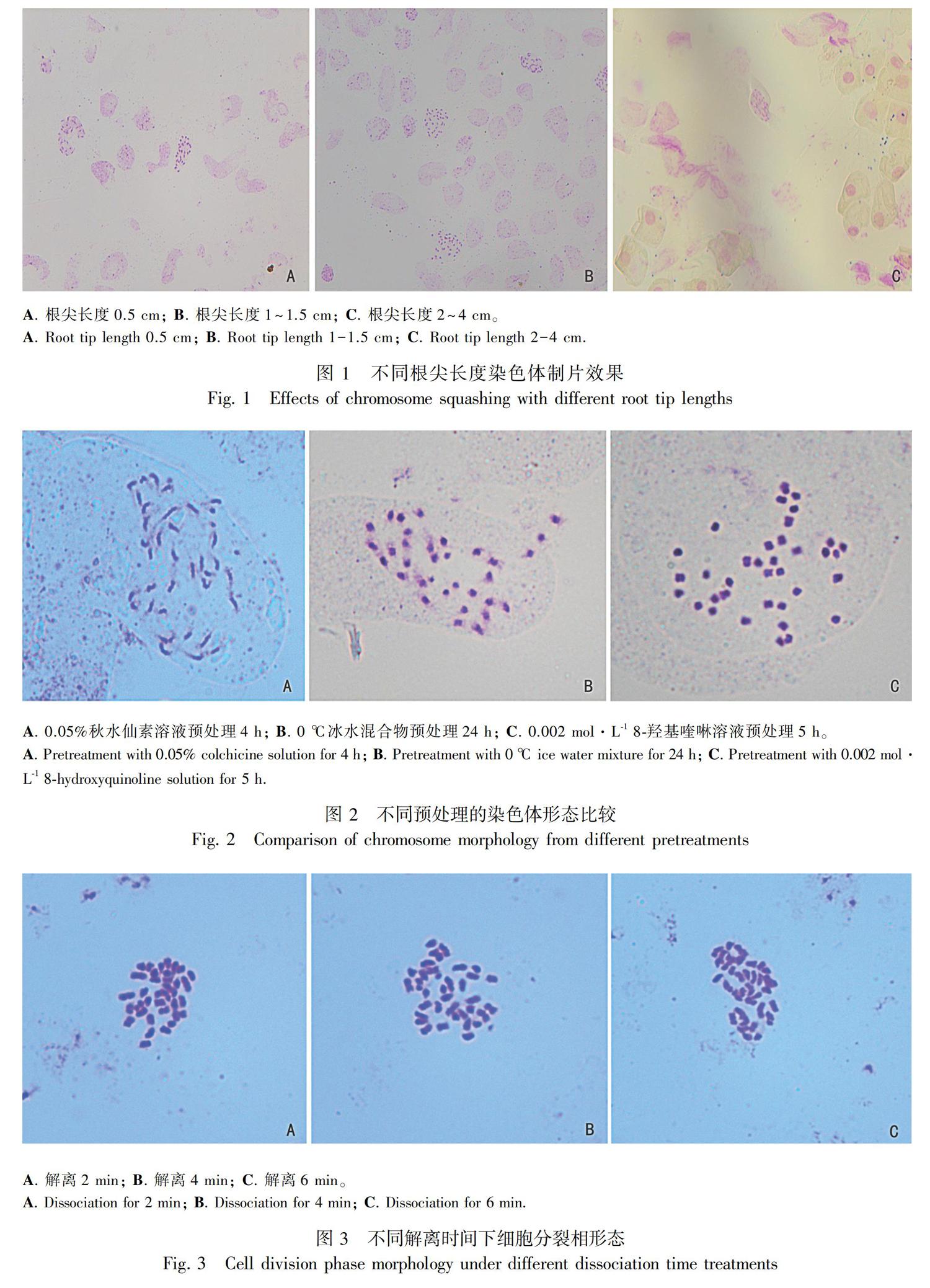
1.2.1.4 解离 将固定好的根尖用蒸馏水清洗3~5次,转到解离液(1 mol·L-1 HCl∶45%冰醋酸=2∶1)中,在60 ℃干式恒温加热器中分别解离2、4、6 min。
1.2.1.5 染色与压片 将解离后的根尖用蒸馏水充分清洗后置于载玻片上,用刀片切取前端乳白色的分生区,滴加少量改良卡宝品红染色液,染色1~2 min后进行常规压片。
1.2.1.6 镜检及染色体计数 将制好的片子放在显微镜Lecia2500下检测,挑选具有有丝分裂中期分裂相、染色体分散和平展良好的细胞于100倍油镜下进行观察和拍照。染色体计数方法参照李懋学和陈瑞阳(1985)的植物核型分析标准。
1.2.2 石山苣苔属和报春苣苔属染色体数目的祖先状态重建 通过查阅文献及染色体数目查询网站Chromosome Counts Database (CCDB)(http://ccdb.tau.ac.il/about/ #IPCN online),对石山苣苔属和报春苣苔属染色体数目已报道的数据进行收集和整理,并结合本研究报道的四种(含一变种)石山苣苔属植物染色体数目,将染色体数目进行编码,在核糖体ITS系统发育树上,用Mesquite软件(Maddison & Maddison,2018)中的MP简约法,通过Trace Character History选项对石山苣苔属和报春苣苔属物种进行染色体数目祖先状态重建,从而追溯染色体数目的演化历史。
核糖体ITS系统发育树的构建:下载NCBI数据库中石山苣苔属和报春苣苔属物种的ITS序列,采用最大似然法(maximum likelihood,ML),碱基替换模型设定为GTRGAMMA(General time reversible model of nucleotide subistution with the gamma model of rate heterogeneity),用随机树作为起始树,矩阵重复抽样1 000次计算支持率(bootstrap support)。具体运行参数设置为raxmlHPC-f a-x 123-p 334-# 1000-s a.phy-m GTRGAMMA-n b。
2 结果与分析
2.1 石山苣苔属染色体标本制备方法的优化
2.1.1 取样时间和根尖长度的优化 比较石山苣苔属植物根尖不同取样时间、取样长度获得的中期分裂相细胞,分析石山苣苔属根尖细胞分裂高峰期,结果见表2。由表2可知,8:30—9:00是根尖细胞分裂旺盛的时间,染色体分离效果良好;20:00—20:30取样则分离效果较差。从图1可以看出,根尖长为1~1.5 cm时,分裂最旺盛,染色体清晰可见且处于中期分裂相的细胞最多,即分离效果最好;根尖长为0.5 cm时,分裂还不够旺盛,观察到的细胞偏少且形态不够好,效果一般;根尖长为2~4 cm时,纤维过多,导致分离效果差。
2.1.2 预处理的优化 采用三种方法进行预处理的优化。表2和图2结果表明,0.05 %秋水仙素溶液预处理4 h后,根尖染色体不够浓缩,形态不好,有拖带现象;0 ℃冰水预处理24 h后,染色体制片效果因物种而异,部分物种(如弄岗石山苣苔)不适用,有拖带现象,分离效果一般;0.002 mol·L-1 8-羟基喹啉水溶液预处理5 h后,制片效果相对最好,染色体呈分散状态,该方法适用于石山苣苔属植物的根尖预处理。
2.1.3 解离时间的优化 比较不同酸解时间下染色体分离的效果。表2和图3结果表明,解离2 min时,时间稍微偏短,染色体相对聚集,不够分散,分离效果一般;解离4 min时,染色体相对分散,分离效果良好;解离6 min时,时间偏长,染色不如2 min和4 min时深,有的细胞破裂,染色體黏连,分离效果差。
2.2 石山苣苔属染色体数目和倍性分析
根据石山苣苔属染色体标本制备方法的优化结果,制备了四种(含一变种)石山苣苔属植物的染色体图(图4)。从图4可以看出,它们的染色体数目一致,均为二倍体2n=2x=36(表3)。
2.3 石山苣苔属和报春苣苔属染色体数目的演变
根据石山苣苔属和报春苣苔属染色体数目的祖先状态重建结果(图5),还不能确定石山苣苔属祖先、报春苣苔属祖先和两属的最近共同祖先的染色体条数。在石山苣苔属内部,出现一次染色体数目由2n=36到2n=20的演化。在报春苣苔属的内部,虽然祖先状态重建未能直接确定唯一在弄岗报春苣苔中出现的2n=72与其余所有2n=36之间的演化方向,但根据常理判断,前者显然由后者经历基因组加倍形成。
3 讨论与结论
3.1 石山苣苔属染色体标本制备条件的优化
石山苣苔属植物为小型染色体,染色体较小的植物根尖组织压片后染色体不易分散且难以观察到清晰的染色体形态, 因此对其取样时间和根尖长度、预处理方法及酸解时间进行了优化研究。
染色体制片若要获得较多的分裂相,需保证植物细胞处于旺盛的有丝分裂期,因此需要找到最佳的取材时间点,同时控制好根尖长度。前人研究表明,大多数植物在8:00—11:00内的某个时间段内取材,中期分裂相较多,染色体制片成功率较高,如橡胶草(Taraxacum kok-saghyz)(杨玉双等,2017)、白刺花(Sophora davidii)(雷文英等,2017)、黄花芥蓝 [yellow-flower Chinese kale(Brassica alboglabra)](夏雪等,2016)等。本研究中,石山苣苔属植物取材时间为8:30—9:00时,中期分裂相较多,这与大多数植物常规取材时间一致。此外,根尖长度对分裂相的多少也有明显影响。前人研究表明,北柴胡(Bupleurum chinense)的根尖长度为0.5~1 cm时,中期分裂相较多(何倚涟等,2018);青花菜(Brassica oleracea var. italica)的根尖长度为1.3~1.5 cm时,中期分裂相最多(张红梅等,2009);不结球白菜(B. campestris ssp. chinensis)的根尖长度为1~2 cm时,中期分裂相数目较多(郑金双等,2011)。本研究结果表明,石山苣苔属根尖长度为1~1.5 cm时,中期分裂相较多。
预处理是染色体制片过程的关键环节,由于植物自身存在的差异,在实验过程中预处理液及预处理时间也存在着差别(刘丹等,2015)。本研究对比不同预处理液的制片效果后认为,四种(含一变种)石山苣苔属植物采用0.002 mol·L-1 8-羟基喹啉溶液作为预处理液制片效果最好。这与李 懋学和张赞平(1996)提出的多种小染色体植物如大豆(Glycine max)、水稻(Oryza sativa)等宜用0.002 mol·L-1 8-羟基喹啉溶液作预处理液是一致的。韩杰等(2018)研究染色体非常小的薄壳山核桃(Carya illinoinensis )时认为,用0.01%的秋水仙素溶液进行预处理效果较好,而用0.002 mol·L-1 8-羟基喹啉溶液效果较差;但本研究中,用0.05%的秋水仙素溶液作为预处理液效果较差,而用0.002 mol·L-1 8-羟基喹啉溶液效果较好。这可能是由于秋水仙素浓度的高低或不同植物特性导致的预处理效果不同。本研究中,0 ℃冰水作为预处理液对部分物种适用,以往测定的三种石山苣苔属植物染色体数目也是采用0 ℃冰水作为预处理液(刘瑞瑞等,2014)。但是,本研究中弄岗石山苣苔相对于其他物种,不宜采用0 ℃冰水进行预处理。
酸解过程中,时间过短,细胞壁难以被破坏,染色体不易分散;时间过长,容易造成染色体断裂或缺失。因此,在实验过程中要准确地控制好各种材料的解离时间。本研究中,四种(含一变种)石山苣苔属植物适宜的解离时间为4 min,这与以往报春苣苔属和石山苣苔属染色体酸解时间3 min较接近(Liu et al.,2012;刘瑞瑞等,2014)。
总的来说,不同物种之间的制片方法存在差异,需要通过多次实验并结合植株本身特性才能得到最佳的染色體制片方案。本研究对石山苣苔属植物叶片根尖染色体制片技术进行了初步探索,可为石山苣苔属及其近缘属植物的染色体制片提供借鉴。
3.2 石山苣苔属染色体数目和倍性分析
本文首次对该四种(含一变种)石山苣苔属植物的染色体数目开展研究。四种(含一变种)植物染色体数目一致,均为二倍体(2n=2x=36),结合已报道的Petrocodon hancei染色体数目2n=20(曹丽敏等,2003)可知,目前该属染色体数目有2n=20、2n=36两种类型,染色体基数有x=10、x=18两种类型。在研究的四个物种(含一变种)中,石山苣苔未定名种的ITS、trnL-F序列与Petrocodon hancei一些个体的序列无差异,在分子系统树上表现为与Petrocodon hancei的一些个体聚在一起。但是,石山苣苔未定名种与Petrocodon hancei有较明显的形态差别,如叶片形状和大小、花冠颜色和下唇内侧条纹等都有所差别,推测其可作为石山苣苔属的变种。已报道的Petrocodon hancei的染色体数目为2n=20(曹丽敏等,2003),但与其亲缘关系最近的石山苣苔未定名种染色体数目为2n=36,其他已知的石山苣苔属植物染色体数目也均为2n=36。因此,在今后的研究中需进一步验证Petrocodon hancei的染色体数目,同时需要对更多的石山苣苔属植物的染色体数目进行研究,从而明确该属物种的染色体数目类型。
由对石山苣苔属和其姐妹属报春苣苔属的染色体数目和倍性分析可知,石山苣苔属除Petrocodon hancei以外,其余物种染色体数目一致,均为二倍体;报春苣苔属除弄岗报春苣苔有四倍体个体以外,其余物种染色体数目和倍性与石山苣苔属一致(Liu et al.,2012;刘瑞瑞等,2014;Kang et al.,2014)。染色体数目、倍性的一致性印证了两属间存在很近的亲缘关系,这与分子系统发育的研究结果相一致。
3.3 石山苣苔属和报春苣苔属染色体数目的演化与两属间物种多样性分化
对石山苣苔属和报春苣苔属染色体数目已报道的数据进行收集和整理,从在分子系统树上追踪染色体数目的变化可以看出,虽然两属之间及两属各自的最近共同祖先染色体数目都不明确,但除了个别物种染色体数目有变化外,两属的染色体数目高度一致(均为2n=36),且已知染色体数目的物种在分子系统树各分支上均有代表,从而推测两属之间、两属各自的最近共同祖先及两属内部各枝系的祖先染色体数目为2n=36的可能性最大。
多倍体植物在自然界中普遍存在,多倍化是植物演化和物种形成过程中最重要的细胞遗传学机制之一(Soltis & Soltis,1999)。前人研究表明,多倍化可能显著增加物种的多样性,增强植株对环境的适应性(Van de Peer et al.,2009;Soltis et al.,2015)。反之,前人也有研究表明,多倍化能减慢物种分化进而呈现较低多样性(Mayrose et al.,2011)。在物种数量上,石山苣苔属约41种,而报春苣苔属有200余种,两属物种数量相差5~6倍。报春苣苔属植物适应性强,不同种类所适应的生境有时截然不同,多数种类生长在阴湿的林下石壁或岩溶洞穴中,大都具有扁平叶;另一些种类则适应了干旱、高温环境,生长在毫无遮阴、阳光直射的裸露岩石石缝或岩石表面上,叶片变为肥厚、多汁的棒状,与生长在荒漠中的仙人掌科或景天科的多肉植物接近(王莉芳等,2009;韦毅刚等,2010)。虽然Christie et al.(2012)推断广义报春苣苔属的染色体数目2n=36起初可能是由祖先具有染色体数目为2n=18直接加倍而来,但是报春苣苔属和石山苣苔属植物中除了一个多倍体外,其他都没有出现多倍化且染色体数目极其稳定,仅个别物种中存在基因组加倍个体。显然,两属物种多样性差异及物种环境适应性转变与基因组加倍事件无关。
植物在演化过程中,除了多倍化以外,还出现了染色体的断裂或融合,使得细胞丢失或增加一条或若干条染色体,即非整倍化(Olson & Gorelick,2011)。在石山苣苔属和报春苣苔属中,除了个别物种外,几乎所有种类染色体条数高度一致(2n=36),因此多倍化和非整倍化在两姐妹属植物演化和物种形成过程中的作用很小。但是,值得关注的是,Petrocodon hancei的染色体数目2n=20与其他2n=36(72)不呈倍数关系。如果该种染色体数目确实无误,那么其演变机制以及染色体数目的减少对植物适应性有何意义,值得深入研究。石山苣苔属和报春苣苔属有的物种具有1个或2个随体,有的物种并无随体(刘瑞瑞,2013)。染色体结构是否存在其他变化,如染色体重排、互相转座和臂间倒位等,以及它们在石山苣苔属和报春苣苔属植物物种分化过程中是否起作用,还有待于进一步研究。
参考文献:
CA O LM, CAO M, TANG XL, et al., 2003. Chromosome numbers of 4 species in the Gesneriaceae from Guangxi [J]. Guihaia, 23(4): 331-333. [曹丽敏, 曹明, 唐咸来, 等, 2003. 广西苦苣苔科四种植物的染色体数目报道 [J]. 广西植物, 23(4):331-333.]
CEN HF, FU LF, WEN F, 2017.Petrocodon urceolatus sp. nov. (Gesneriaceae) from Zhangjiajie City, northwestern Hunan Province, China [J]. Nord J Bot, 35(3): 336-338.
CHEN WH, MOLLER M, SHUI YM, et al., 2014. Tree new species of Petrocodon (Gesneriaceae), endemic to the limestone areas of Southwest China, and preliminary insights into the diversifcation patterns of the genus [J]. Syst Bot, 39(1): 316-330.
CHEN L, CHEN WH, GUO SW, et al., 2019. Petrocodon tenuitubus (Gesneriaceae), a new species from SE Yunnan,China [J]. Guihaia, 39(5): 574-580. [陳力, 陈文红, 郭世伟, 等, 2019. 云南东南部石山苣苔属一新种——细管石山苣苔(英文) [J]. 广西植物, 39(5):574-580.
CHRISTIE F, BARBER S, MOLLER M, 2012. New chromosome counts in Old World Gesneriaceae: numbers for species hitherto regarded as Chirita, and their systematic and evolutionary significance [J]. Edinb J Bot, 69(2): 363-365.
GUO J, MENG T, ZHANG Q, et al., 2016.Petrocodon retroexus sp. nov. (Gesneriaceae) from a karst cave in Guizhou, China [J]. Nord J Bot, 34(2): 159-164.
HAN J, KONG DY, PENG FR, 2018. Optimization of chromosome mounting technique and karyotype analysis of Carya illinoinensis [J]. Mol Plant Breed, 16(16): 5240-5246. [韩杰, 孔德仪, 彭方仁, 2018. 薄壳山核桃染色体制片技术的优化与核型分析 [J]. 分子植物育种, 16(16):5240-5246.]
HE YL, LIU HW,YANG B, et al., 2018. Study on chromosomes preparation on technique of root tip of Buplerum chinese DC [J]. Seed, 37(8): 68-71. [何倚涟, 刘宏伟, 杨彬, 等, 2018. 北柴胡根尖染色体制片技术研究 [J]. 种子, 37(8):68-71.]
HONG X, ZHOU SB, WEN F, 2014. Petrocodon villosus (Gesneriaceae), a new species from Guangxi, China [J]. Blumea, 59(1): 33-36.
KANG M, TAO J, WANG J, et al., 2014.Adaptive and nonadaptive genome size evolution in karst endemic flora of China [J]. New Phytol, 202(4): 1371-1381.
LEI WY, ZHAO LL, JIAN ZL, et al., 2017. Optimization of chromosome preparation and karyotype analysis of Sophora davidii [J]. Seed, 36(8): 5-9. [雷文英, 赵丽丽, 简忠领, 等, 2017. 白刺花染色体压片技术优化及核型分析 [J]. 种子, 36(8): 5-9.]
LI CR, LIU FP, GADAGKAR SR, et al., 2019. Petrocodon longitubus (Gesneriaceae), a new species from Guizhou, China [J]. Phytotaxa, 408 (4): 267-275.
LI MX, CHEN RY, 1985. A suggestion on the standardization of karyotype annalysis in plants [J]. Acta Bot Wuhan, 3(4): 297-302. [李懋学, 陈瑞阳, 1985. 关于植物核型分析的标准化问题 [J]. 武汉植物学研究, 3(4):297-302.]
LI ZY, WANG YZ, 2005. Plants of Gesneriaceae in China [M]. Henan: Henan Science and Technology Publishing House. [李振宇, 王印政, 2005. 中国苦苣苔科植物 [M]. 河南:河南科学技术出版社.]
LI HQ, WANG YQ, 2015. Petrocodon confertiorus (Gesne-riaceae), a new species from Guangdong, China [J]. Ann Bot Fenn, 52: 27-36.
LI MX, ZHANG ZP, 1996. Crop chromosomes and their research techniques [M]. Beijing: China Agriculture Press: 1-40. [李懋学, 张赞平, 1996. 作物染色体及其研究技术 [M]. 北京:中国农业出版社:1-40.]
LIU RR, PAN B, ZHOU TJ, et al., 2012. Cytological studies on Primulina taxa (Gesneriaceae) from limestone karsts in Guangxi Province, China [J]. Caryologia, 65(4): 295-303.
LIU RR, 2013. Cytological characteristics of Primulina sensu lato (Gesneriaceae) and their systematic implications [D]. Beijing: University of Chinese Academy of Sciences. [刘瑞瑞, 2013. 广义报春苣苔属(苦苣苔科)的细胞学特征及其系统学意义 [D]. 北京:中国科学院大学.]
LIU RR, SKOG LE, LIAO JP, et al., 2014. New chromosome counts and their taxonomic implications in Primulina sensu lato ( Gesneriaceae) [J]. Plant Divers Resour, 36(1): 13-21. [刘瑞瑞, SKOG LE, 廖景平, 等, 2014. 廣义报春苣苔属(苦苣苔科)的染色体新计数及其分类学意义(英文) [J]. 植物分类与资源学报, 36(1):13-21.]
LIU D, XIA X, WU YM, et al., 2015. Analysis of factors affecting chromosome squashing effect in plant [J]. J Zhejiang Agric Sci, 56(10): 1654-1657. [刘丹, 夏雪, 吴益梅, 等, 2015. 植物染色体制片效果影响因素的解析 [J]. 浙江农业科学, 56(10):1654-1657.]
LU YB, HUANG YS, XU WB, et al., 2017. Floral evolutionary trend of Petrocodon (Gesneriaceae) and its significance in classification [J]. Guihaia, 37(10): 1227-1239. [卢永彬, 黄俞淞, 许为斌, 等, 2017. 石山苣苔属(苦苣苔科)花形态演化及分类学意义(英文) [J]. 广西植物, 37(10):1227-1239.]
LU YB, PAN BO, HUANG J, et al., 2017. Petrocodon pulchriflorus sp. nov. (Gesneriaceae) from Guangxi, China [J]. Phytotaxa, 291(1): 81-88.
MADDISON WP, MADDISON DR, 2018. Mesquite: A modular system for evolutionary analysis [EB/OL]. Version 3.51, http://www.mesquiteproject.org.
MAYROSE I, ZHAN SH, ROTHFELS CJ, et al., 2011. Recently formed polyploid plants diversify at lower rates [J]. Science, 333(6047): 1257-1257.
MIDDLETON DJ, SANGVIROTJANAPAT S, LA-ONGSRI W, 2015. A new species of Petrocodon (Gesneriaceae) from Thailand [J]. Thai For Bull Bot, 43: 15-17.
MOLLER M, KIEHN M, 2004. A synopsis of cytological studies in Gesneriaceae [J]. Edinb J Bot, 60(3): 425-447.
OLSON K, GORELICK R, 2011. Chromosomal fission accounts for small-scale radiations in Zamia (Zamiaceae; Cycadales) [J]. Bot J Linn Soc, 165(2): 168-185.
SOLTIS PS, MARCHANT DB, VAN DE PEER Y, et al., 2015. Polyploidy and genome evolution in plants [J]. Sci Direct, 35: 119-125.
SOLTIS DE, SOLTIS PS, 1999. Polyploidy: recurrent formation and genome evolution [J]. Trends Ecol Evol, 14(9): 348-352.
SU LY, PENG T, ZHAO ZG, et al., 2019. Petrocodon chongqingensis, a new species of Gesneriaceae from Chongqing City, China [J]. Guihaia, 39(8): 997-1006. [苏兰英, 彭涛, 赵志国, 等, 2019. 苦苣苔科石山苣苔属一新种——重庆石山苣苔 [J]. 广西植物, 39(8): 997-1006.]
VAN DE PEER Y, MAERE S, MEYER A, 2009. The evolutionary significance of ancient genome duplications [J]. Nat Rev Genet, 10(10): 725-736.
WANG LF, HUANG SX, DENG T, et al., 2009. The resources of ornamental plants and landscape application in Chirita in Guangxi [J]. N Hortic, (4):174-177. [王莉芳, 黄仕训, 邓涛, 等, 2009. 广西唇柱苣苔属植物及其园林应用 [J]. 北方园艺, (4):174-177.]
WANG WT, PAN KY, LI ZY, et al., 1998. Gesneriaceae [M]//WU ZY, RAVEN PH. Flora of China. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press, 18: 244-401.
WEBER A, WEI YG, PUGLISI C, et al., 2011.A new definition of the genus Petrocodon (Gesneriaeae) [J]. Phytotaxa, 23: 49-67.
WEI YG, WEN F, MOLLER M, et al., 2010. Gesneriaceae of South China [M]. Nanning: Guangxi Science & Technology Publishing House. [韦毅刚, 温放,
MOLLER M, 等, 2010. 华南苦苣苔科植物 [M]. 南宁:广西科学技术出版社.]
XIA X, GU JH, LIU D, et al., 2016. Optimization of chromosome preparation and karyotype analysis of yellow-flower Chinese kale [J]. J Zhejiang Univ (Agric & Life Sci), 42(5): 527-534. [夏雪, 辜金花, 劉丹, 等, 2016. 黄花芥蓝染色体制片优化及核型分析 [J]. 浙江大学学报(农业与生命科学版), 42(5):527-534.]
XU WB, MENG T, ZHANG Q, et al., 2014.Petrocodon (Gesneriaceae) in the limestone karsts of Guangxi, China: Three new species and a new combination based on morphological and molecular evidence [J]. Syst Bot, 39(3): 965-973.
YANG YS, GAN L, QIN B, et al., 2017. Development of technique for chromosome squashing of taraxacum kok-saghyz rodin [J]. Chin J Trop Agric, 37(2): 31-34. [杨玉双, 甘霖, 覃碧, 等, 2017. 橡胶草染色体制片技术研究 [J]. 热带农业科学, 37(2): 31-34.]
YU XL, LI M, ZHOU JJ, et al., 2015. Petrocodon hunanensis (Gesneriaceae), a new species identifed by both morpholo-gical and molecular evidence from limestone area in Hunan, China [J]. Phytotaxa, 195: 65-72.
ZHANG RB, DENG T, FU LF, et al., 2019. Petrocodon tongziensis (Gesneriaceae), a new species from limestone areas in Guizhou, China based on morphological and molecular evidence [J]. Nord J Bot, 37(2): e01774. doi: 10.1111/njb.01774.
ZHANG RL, FU LF, LI S, et al., 2018. Petrocodon asterocalyx, a new species of Gesneriaceae from Guangxi, China [J]. Phytotaxa, 363(3): 259-268.
ZHANG HM, ZHANG SN, KONG YE, et al., 2009. Staining and slide-preparing technique of chromosome and karyotype analysis of broccol [J]. J Nanjing Agric Univ, 32(4): 33-36. [张红梅, 张蜀宁, 孔艳娥, 等, 2009. 青花菜染色体制片技术及核型分析 [J]. 南京农业大学学报, 32(4): 33-36.]
ZHENG JS, ZHANG SN, SUN CZ, et al., 2011. Chromosome preparation of somatic cell in root-tip of non-heading Chinese cabbage (Brassica campestris ssp. chinensis) and observation of mitosis process of its diploid and tetraploid [J]. J Plant Resour Environ, 20(4): 58-63. [郑金双, 张蜀宁, 孙成振, 等, 2011. 不结球白菜根尖体细胞染色体制片及其二倍体和四倍体有丝分裂过程观察 [J]. 植物资源与环境学报, 20(4): 58-63.]
( 责任编辑 蒋巧媛 )
