
鲟营养需求与饲料研究进展
2020-12-10 03:55张配瑜刘海燕杨振才
水生生物学报 2020年6期
张配瑜 刘海燕 杨振才
(河北师范大学生命科学学院, 石家庄 050024)
鲟(Sturgeons)是软骨硬鳞鱼, 属于脊索门, 硬骨鱼纲, 辐鳍亚纲, 鲟形目, 鲟科。共有27个物种, 其中4种已灭绝[1]。 现存23种分为4个属, 其中鳇属(Huso)2种, 铲鲟属(Scaphirhynchus)2种, 拟铲鲟属(Pseudoscaphirhynchus)3种, 鲟属(Acipenser)16种[2]。由于鲟的鱼肉鲜美和其鱼卵可以被加工成鱼子酱,因此具有较高的商业价值[3]。在自然生境中, 鲟由于生长缓慢和性成熟晚, 加上过度捕捞、自然栖息地的破坏、水质恶化、大坝建设等导致85%的鲟物种面临灭绝[4—6]。缓解鲟自然种群下降的最优策略是通过鲟的水产养殖以满足市场对鱼子酱和鱼肉需求[7]。中国是鲟和鱼子酱的主要生产国和出口国, 鲟养殖产业在鲟资源保护和渔业发展中发挥着重要作用。通过全球抽样调查, 推测2017年中国鲟产量为7.96×107kg, 约占世界总产量的78%, 之后分别是俄罗斯(6.80×106kg), 亚美尼亚(6.00×106kg),伊朗(2.51×106kg)和其他产量少于1.00×106kg的52个国家[8]。2016年世界鲟产量前三的品种分别是西伯利亚鲟(Acipenser baerii)、杂交鲟(达氏鳇×施氏鲟Huso dauricus♀×A. schrencki♂、西伯利亚鲟×施氏鲟A. baerii♀×A. schrenckii♂)和施氏鲟(A.schrenckii), 分别占总产量的39.5%、35.6%和10.2%[8]。我国开始鲟养殖以来, 进行大规模商业化养殖的品种也是西伯利亚鲟、达氏鳇×施氏鲟杂交鲟、西伯利亚鲟×施氏鲟杂交鲟和施氏鲟, 占国内鲟养殖总产量的90%[9]。主要养殖区域集中在山东、云南、湖北、贵州和湖南等省份, 上述省份2017 年的鲟养殖产量分别为1.14×107、0.96×107、0.81×107、0.79×107和0.65×107kg, 分别占全国鲟养殖总产量的13.8%、11.6%、9.7%、9.5%和7.9%[10]。
在全球鲟养殖方面, 早期一般用鲑鳟鱼饲料来饲喂鲟, 但长期的投喂会导致鲟生长较差和相关营养缺乏症, 之后关于鲟的营养需求研究逐渐增加,但主要研究对象是美国白鲟和西伯利亚鲟[11, 12], 对于其他鲟品种营养需求与利用的系统研究相对较少, 但这些信息对鲟的养殖又至关重要。近年来对主养的鲟品种营养需求研究越来越多, 本文就鲟对基本营养素和微量营养素的需求以及饲料的研究进行综述, 以供后续研究参考。
1 鲟营养需求研究进展
1.1 蛋白质营养研究
蛋白质需求关于鲟蛋白质需求的研究较多。Moore等[13]以酪蛋白、谷朊粉、鸡蛋白为蛋白源(比例为62﹕30﹕8), 配制蛋白含量为20.0%—52.7%配合饲料开展为期8周的养殖实验, 以增重率为评价指标, 发现高首鲟[Acipenser transmontanus,(145.20±0.20) g, 本处及后文都指初始体重]的蛋白需求量为40.5%。Mohseni等[14]以鱼粉和小麦粉为主要蛋白源, 以蛋白质效率为评价指标, 指出波斯鲟(Acipenser persicus, 136.8 g)的蛋白需求量为40%,合适的蛋能比18—20 mg/kJ。Guo等[15]以鱼粉和豆粕为蛋白源, 配制7种蛋白水平(250、300、350、400、450、500、550 g/kg饲料)的等能饲料, 以特定生长率为评价指标, 采用二项式回归分析得出杂交鲟[西伯利亚鲟×俄罗斯鲟Acipenser baerii♀×A.gueldenstaedtii♂, (25.10±0.12) g]的蛋白需求为370 g/kg饲料。刘阳洋[16]以鱼粉为主要蛋白源, 饲喂匙吻鲟[Polyodon spathula, (158.00±0.29) g] 8周, 以体增重为评价指标, 得出最适蛋白水平为40.67%。李同庆等[17]研究了4种蛋白水平(28%、32、36和41%)的饲料对施氏鲟的影响, 36%蛋白组生长速度最快。关于鲟的蛋白需求见表 1。从表 1可以看出,大部分鲟幼鱼的蛋白需求在40%左右, 杂交鲟、施氏鲟的蛋白需求约36%—37%。
合适的蛋能比饲料中的蛋能比在鱼类营养中扮演着重要角色, 影响鱼体的生长、饲料效率和体成分。合适的蛋能比既有利于鱼体的快速生长, 也可降低饲料成本和含氮废物的排放[18]。关于鲟合适的蛋能比研究不多, Guo等[15]得出25 g杂交鲟(西伯利亚鲟×俄罗斯鲟)适宜蛋白和能量为40%和17.9 kJ/g, 蛋能比为21.9 mg蛋白/kJ。Mohseni等[14]以鱼粉和小麦粉为主要蛋白源, 以蛋白质效率为评价指标, 指出波斯鲟(136.8 g)合适的蛋能比为18—20 mg/kJ。Mohseni等[19]以鱼粉为主要蛋白源,鱼油和玉米油(1﹕1)为主要脂肪源, 设计3组蛋白水平(40%、45%和50%)和4组能量水平(16、17、18和19 kJ/g)的12种饲料, 饲喂波斯鲟[(103.3±3.5) g]14周, 发现最大特定生长率(SGR)出现在40%蛋白水平和19 kJ/g的饲料组, 但与40%蛋白水平和18 kJ/g的饲料组无显著差异, 饲料效率在各组间无显著差异, 基于成本考虑和SGR, 波斯鲟最适蛋能比为22.0 mg/kJ。由表 1可知, 大多数鲟品种适宜蛋能比为18—22 mg/kJ。
必需氨基酸的需求蛋白质在经过鱼体消化酶分解后, 变成氨基酸进入体内, 鱼体对蛋白质的需求实质上是对蛋白质中平衡氨基酸的需求。有关鲟必需氨基酸需求的研究很少。研究表明, 鲟必需氨基酸主要有: 赖氨酸、亮氨酸、精氨酸、组氨酸、异亮氨酸、蛋氨酸、苯丙氨酸、苏氨酸、色氨酸和缬氨酸[24], 且含硫氨基酸(蛋氨酸和胱氨酸)是杂交鲟(达氏鳇♀×施氏鲟♂ Huso dauricus ♀×Acipenser schrincki ♂)肌肉的第二限制性氨基酸[25]。目前, 仅一篇文章用传统剂量反应来评估鲟的氨基酸需求, Wang等[26]以鱼粉、次粉、豆粕和玉米蛋白粉为蛋白源, 配制7种不同水平精氨酸(1.74%、2.04%、2.34%、2.64%、2.94%、3.24%和3.54%)的饲料, 饲喂杂交鲟[施氏鲟×西伯利亚鲟A.schrenckii ♀×A. baerii♂, (3.63±0.08) g] 8周, 以特定生长率(SGR)为评价指标, 得出饲料中最适精氨酸需求为2.47%, 占6.18%饲料蛋白。全鱼或肌肉组织必需氨基酸模式与必需氨基酸的需求呈现出高度关联, 因此首先通过传统剂量反应评估其中一种必需氨基酸的需求, 再根据此种必需氨基酸占全鱼或肌肉总必需氨基酸的比例, 即可推算出其他必需氨基酸的需求量[27]。Wang等[28]用以鱼粉和大豆浓缩蛋白为蛋白源饲喂杂交鲟[施氏鲟×西伯利亚鲟,(7.68±0.08) g] 8周, 其肌肉中必需氨基酸的含量分别为苏氨酸32.47、缬氨酸36.68、蛋氨酸20.57、异亮氨酸23.70、亮氨酸49.47、苯丙氨酸17.65、赖氨酸50.00、组氨酸15.45和精氨酸(44.08 g/kg氨基酸)。因此各必需氨基酸占总必需氨基酸的比例分别为苏氨酸(11.19%)、缬氨酸(12.64%)、蛋氨酸(6.98%)、异亮氨酸(8.17%)、亮氨酸(17.05%)、苯丙氨酸(6.08%)、赖氨酸(17.24%)、组氨酸(5.33%)和精氨酸(15.20%), Wang等[28]得出该杂交鲟精氨酸需求量是2.47% (g/100g饲料), 因此推算其他必需氨基酸的需求量分别为苏氨酸(1.82%)、缬氨酸(2.05%)、蛋氨酸(1.13%)、异亮氨酸(1.33%)、亮氨酸(2.77%)、苯丙氨酸(0.99%)、赖氨酸(2.80%)和组氨酸(0.87%)。Ng和Hung[24]评估了高首鲟必需氨基酸的需求量(表 2), 通过对比, 发现高首鲟和杂交鲟的必需氨基酸需求较接近, 因此, 杂交鲟和高首鲟的必需氨基酸需求可供其他品种鲟参考, 但需要进一步的证实。
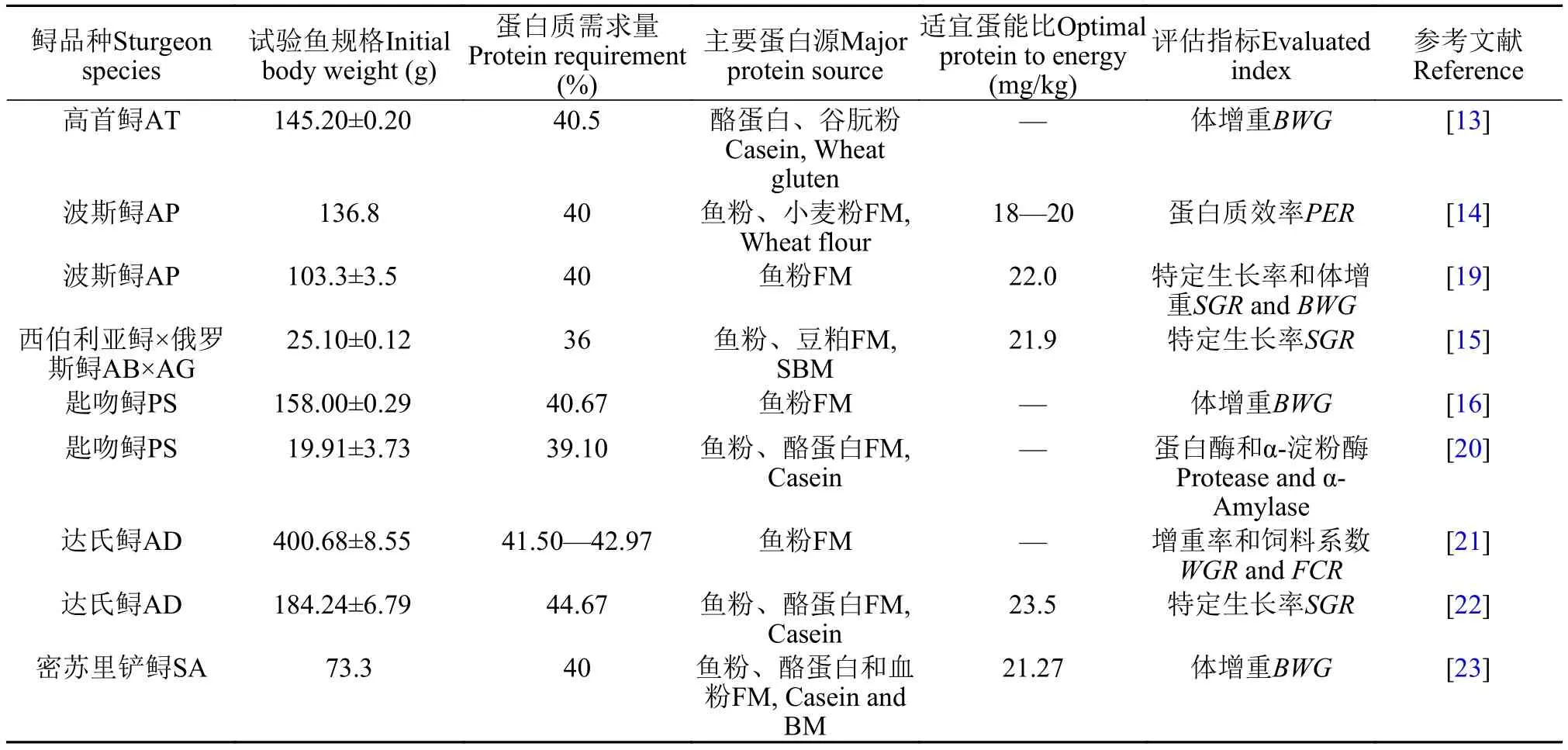
表 1 鲟对蛋白质需求量Tab. 1 The dietary protein requirement of sturgeons (%)
替代蛋白源的研究进展近些年, 由于鱼粉价格飙升, 寻找合适、廉价的替代蛋白源成为水产养殖的当务之急。表观消化率是评价水产动物对饲料原料的营养品质最基本的手段之一[29]。关于鲟对替代蛋白源消化率的报道较少。Liu等[30]评估了8.4 g西伯利亚鲟对鱼粉、肉骨粉、家禽副产品粉、水解羽毛粉、发酵羽毛粉、溶剂萃取棉粕和豆粕的蛋白消化率, 分别为94.5%、84.5%、90.4%、90.9%、87.7%、87.6%和91.8%, 研究表明幼鲟可以有效地利用动植物蛋白源, 但在制作配方时需考虑必需氨基酸的需求。Safari等[29]研究了达氏鳇(17.70±0.234) kg对5类不同蛋白原料(3种鱼粉、3种陆生动物副产品、3种植物浓缩蛋白、9种高植物蛋白源和3种低植物蛋白源)进行消化率的评估,发现鳀鱼粉(93.2%)的蛋白表观消化率高于沙丁鱼鱼粉(90.2%)和墨西哥沙丁鱼鱼粉(89.5%), 但无显著差异; 对于动物副产品, 达氏鳇对家禽副产品粉(91.7%)的蛋白消化优于羽毛粉(75.0%)和血粉(73.1%); 对于植物蛋白浓缩物(大豆浓缩蛋白、玉米蛋白粉和小麦蛋白粉)的蛋白消化率都超过了80%, 且对大豆浓缩蛋白消化率最高; 另外, 达氏鳇对高植物蛋白源的蛋白消化率(62.9%)高于低植物蛋白源(41.5%)。Stuart和Huang[31]研究高首鲟对不同蛋白源的利用情况, 依据能量留存率, 鲟蛋白源质量顺序为: 酪蛋白>脱脂虾粉>鳀鱼粉>大豆浓缩蛋白>鸡蛋蛋白>明胶, 由于酪蛋白组的死亡率为11.7%, 根据生长结果脱脂虾粉可以完全替代且优于鳀鱼粉。由表 3可知, 西伯利亚鲟可很好地消化动植物蛋白源, 而达氏鳇对蛋白源的消化效果顺序为鱼粉>植物蛋白浓缩物>动物副产品>高蛋白植物源>低蛋白植物源, 杂交鲟对豆粕的消化率高于家禽副产品粉。
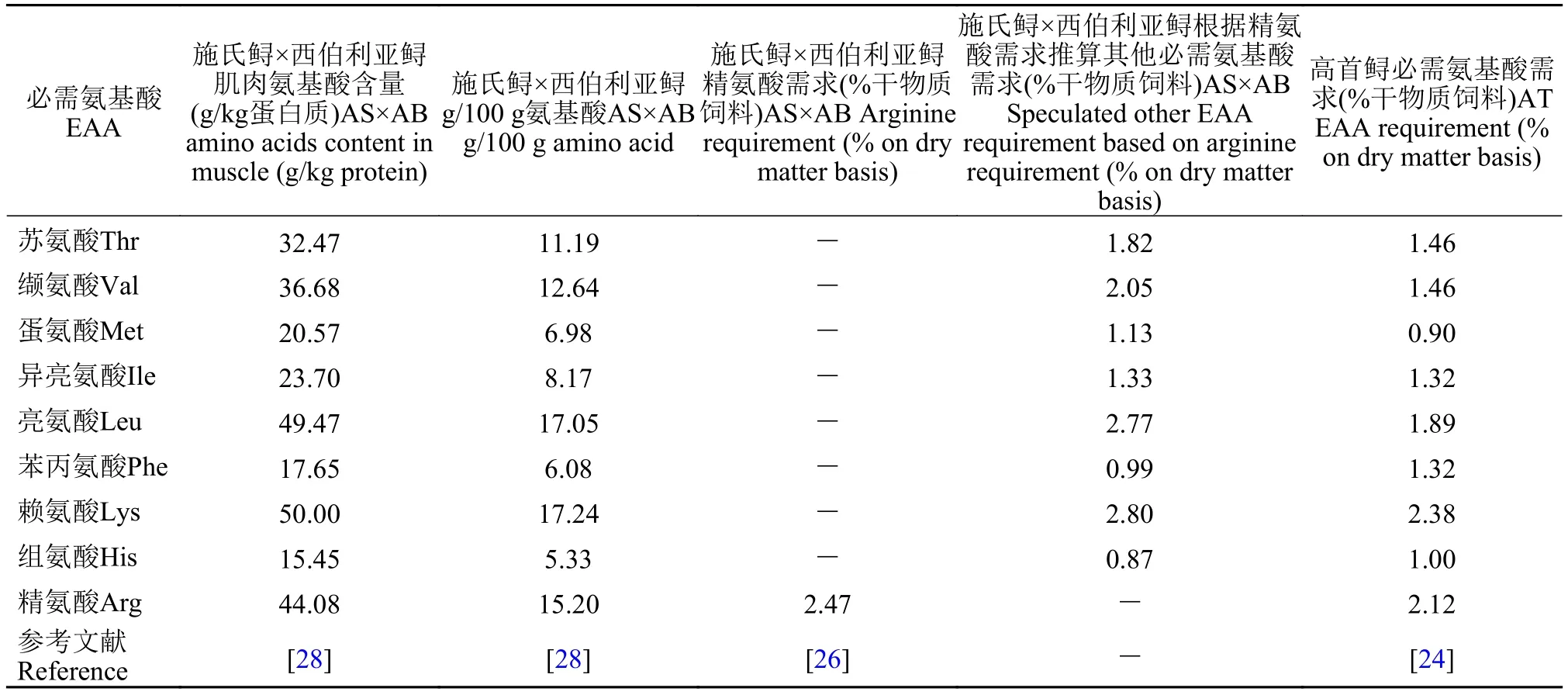
表 2 鲟对必需氨基酸的需求Tab. 2 The dietary essential amino acids requirement in sturgeons
Sicuro等[33]以杂交鲟(意大利鲟×西伯利亚鲟Acipenser naccarii×Acipenser baerii)为研究对象, 用玉米蛋白粉替代饲料中85%的鱼粉(玉米蛋白粉含量为55%), 进行为期60d的养殖实验, 结果显示对鲟生长无负面影响, 且饲料成本可减少20%。Xu等[34]以大豆分离蛋白替代施氏鲟饲料中0—100%的鱼粉, 并在100%替代水平添加晶体氨基酸(蛋氨酸、赖氨酸、苏氨酸), 以增重率为评价指标, 大豆分离蛋白适宜的替代比例为57.64%, 且晶体氨基酸的添加对生长无促进作用。Jahanbakhshi等[35]发现芝麻饼粕和玉米蛋白粉(1﹕1)替代达氏鳇饲料中37%的鱼粉对血液指标无显著影响。Yun等[36]用混合植物蛋白源(豆粕﹕小麦蛋白粉=6﹕4)在两个蛋白水平(36%和40%)下完全替代西伯利亚鲟饲料中的鱼粉,并补充晶体氨基酸和磷酸氢钙, 进行为期8周的养殖实验, 发现植物蛋白源替代鱼粉对生长无负面影响, 且能有效降低磷的排放。由表 4可见, 鲟可以较好地利用一些植物蛋白源, 分析其原因可能是替代源有较高的营养价值, 比如大米浓缩蛋白含有75.5%的蛋白质且有较高含量的n-6高不饱和脂肪酸[37]。玉米蛋白粉蛋白含量较高(67%—71%), 不含有抗营养因子, 较低含量的纤维素, 且含有生物活性肽[38, 39]。豆粕和大豆浓缩蛋白也含有较多的蛋白质, 广泛应用于水产饲料中。其次, 鲟能较好地消化吸收这些植物蛋白源, 由表 3可以看出, 鲟对大豆浓缩蛋白、麦麸和玉米蛋白粉的蛋白质消化率都高达80%以上; 第三, 鲟能够耐受较高的碳水化合物, 在Yun 等[36]的研究中发现西伯利亚鲟可耐受362.5 g/kg饲料的碳水化合物。另外, 杂交鲟(意大利鲟×西伯利亚鲟)可以耐受39.4%的碳水化合物[33],这是因为鲟肠道内含有较高的淀粉酶活性[40], 至于鲟能耐受碳水化合物的最大程度, 还有待进一步的研究。

表 3 鲟对不同蛋白源的表观消化率Tab. 3 Apparent digestibility in different protein sources consumed by sturgeons (%)

表 4 鲟饲料中其他蛋白源替代鱼粉量Tab. 4 The replacement amount of dietary fish meal by other protein resources for sturgeons (%)
在西伯利亚鲟(6.77 g)的研究中, 用酪蛋白酸钠替代0%、20%、40%、60%、80%和100%的鱼粉,进行为期50 d的养殖实验, 发现20%—60%水平下的鲟特定生长率最高[41]。Zhu等[42]在为期8周的西伯利亚鲟养殖实验中[(28.9±0.2) g], 发现混合动物蛋白源(肉骨粉﹕家禽副产品粉﹕水解羽毛粉 =4﹕4﹕2)可替代50%的鱼粉, 且外源添加晶体氨基酸(苏氨酸和赖氨酸)对鱼体生长有良好的促进作用。同样在西伯利亚鲟的研究中, Xue等[43]发现在36%和40%两个蛋白水平下, 混合动物蛋白源(家禽副产品粉﹕肉骨粉﹕水解羽毛粉﹕血粉=40﹕35﹕20﹕5)可以完全替代鱼粉, 在添加晶体氨基酸的情况下, 对鱼体生长无负面影响。Gong等[44]以磷虾粉替代鱼粉,以增重率和特定生长率为评价指标, 发现替代60%鱼粉俄罗斯鲟(481 g)的生长表现优于鱼粉组。Józefiak等[45]发现用全脂黄粉虫粉和黑水虻虫粉可以替代15%的鱼粉, 而对西伯利亚鲟无显著性影响。Caimi等[46]也发现黑水虻幼虫粉替代26.4%的鱼粉对西伯利亚鲟[(24.2±7.59) g]的肝脏和后肠微观结构无负面影响。由表 4可见, 鲟可以很好地利用部分动物蛋白源。
综上所述, 目前关于鲟替代蛋白源的研究主要集中在西伯利亚鲟上, 对于其他品种的鲟研究较少,西伯利亚鲟可以较好地利用植物蛋白源和动物蛋白源, 且能够较好地利用晶体氨基酸, 关于其他鲟品种饲料学的蛋白原料评价需要更多的研究。
1.2 脂肪营养需求研究进展
脂肪需求关于鲟脂肪需求的研究较少,Hung等[51]以110 g高首鲟为研究对象, 通过饲喂不同水平的鱼油(25.8%、30.4%、35.7%和40.2%)的饲料, 以SGR和饲料效率为评价指标, 发现高脂饲料(25.8%—35.7%)对鲟的生长无负面影响, 低蛋白高脂饲料(37.8%蛋白、40.2%脂肪)对生长有抑制作用, 可能是低蛋白高脂饲料降低了鲟的摄食量或饲料效率。Mohseni等[14]采用正交实验法设计2种蛋白水平(40%和45%)和4种脂肪水平(鱼油:10%、15%、20%和25%)共8种饲料, 饲喂波斯鲟(136.8 g) 150d, 以体增重为评价指标, 40%蛋白和25%脂肪处理组的增重率最高, 优于45%蛋白和25%脂肪处理组, 且在40%蛋白水平下, 增重率会随着饲料脂肪增加而增加, 表明波斯鲟可利用高脂饲料。Keramat等[52]用高脂(30%)和低脂(15%)饲料在2个投喂水平下(饱食和60%饱食)饲喂达氏鳇[(61.3±4.4) g] 90d, 发现高脂饲料组的体增重显著高于低脂饲料, 表明达氏鳇可很好地利用饲料中的脂肪。Zhu等[53]以葵花油、鱼油、亚麻油为混合脂肪源,配制成3种不同脂肪水平(5%、15%和25%)的俄罗斯鲟[(9.61±0.03) g]饲料, 进行为期8周的养殖实验,发现鲟在25%脂肪水平下生长最佳。Guo等[54]以豆油和鱼油(1﹕1)为脂肪源, 在蛋白水平为37%下配制不同脂肪水平(4%—19%)的杂交鲟饲料[西伯利亚鲟×俄罗斯鲟, (66.7±0.11) g], 以SGR为评估指标, 采用折线回归模型拟合鲟饲料最适脂肪水平为11.1%。肖懿哲等[55]以酪蛋白、鱼粉、糊精和纤维素等为主要原料配制而成的等氮(蛋白质含量41%)和等能(11.2 MJ/kg饲料)半精制饲料, 以等量混合的海水鱼油和豆油作为添加的脂肪源, 研究了油脂的添加水平对施氏鲟的影响, 提出8%是适宜的添加量(不包括饲料其他原料的脂肪)。关于鲟脂肪需求的研究进展见表综上所述, 波斯鲟、俄罗斯鲟、高首鲟和达氏鳇都能较好地利用高脂饲料(脂肪>25%), 但脂肪的精准需求还需要进一步的研究, 因为上述实验鲟生长最佳出现在最高脂肪水平处理组, 可能鲟还可利用更多的脂肪; 而杂交鲟 (西伯利亚鲟×俄罗斯鲟)的脂肪需求只有11.1%, 因此不同鲟品种对脂肪需求不同, 可能与体内脂肪酶活性高低有关。
脂肪酸的需求一般认为海洋鱼类不具备将短链不饱和脂肪酸转化为长链不饱和脂肪酸的能力, 而淡水鱼具备这种能力[56]。鲟是洄游性鱼类,据报道, 高首鲟具有合成长链不饱和脂肪酸的能力[57]。因此, 鲟饲料中需要补充不饱和脂肪酸以满足其正常生长和生理需求。Wang等[58]用葵花油和亚麻油为混合油, 用含有不同水平的α-亚麻酸饲料饲喂俄罗斯鲟[(11.68±0.13) g] 8周, 根据特定生长率和饲料转化效率, 鲟α-亚麻酸需求量为6.85—10.69 g/kg饲料, 过量的亚麻酸会抑制鱼体生长。
鱼类对必需脂肪酸除了考虑量的需求, 还需要考虑必需脂肪酸的比例。Li等[59]分别用葵花油和紫苏油替代椰子油, 配制成0.5%、1%、2%的亚油酸(LA)和亚麻酸(LN), 及用葵花油和紫苏油同时替代椰子油, 配制成含有0.5%LA+0.5%LN、1%LA+1%LN的俄罗斯鲟[(9.59±0.19) g]饲料, 以增重率为评估指标, 鲟的LA和LN需求为1%和1%。Zhu等[53]以鱼油、亚麻油和葵花油配制成3个脂肪水平(5%、15%和25%), 每个脂肪水平下3种n-3/n-6多不饱和脂肪酸(PUFA)比例(1﹕3、1﹕1和3﹕1)的9种饲料, 饲喂俄罗斯鲟[(9.61±0.03) g] 8周, 以增重率为评估指标, 发现脂肪水平为25%、n-3/n-6 PUFA为1﹕3时生长最佳。
鱼油替代的研究进展随着水产养殖业的不断发展, 鱼油的供需矛盾日益突出, 寻找合适和廉价的鱼油替代物成为学者和相关企业关注的焦点。鲟鱼油替代源的研究较多, Xu等[57]在高首鲟(35 g)饲料中添加15%混合油(玉米油、鱼肝油、猪油, 1﹕1﹕1)、玉米油、鱼肝油、猪油、亚麻油、豆油、红花油和加拿大菜油, 发现各组的体增重、饲料效率和体成分无显著影响。在短期养殖过程中(30—90d), 亚麻油可完全替代鱼油对生长无负面影响, 但在长期养殖过程中(360d), 则对鲟生长产生负面影响[60]。同样地, Hosseini等[61]发现加拿大菜油和豆油可完全替代欧洲鳇[(206±7.3) g]饲料中的鱼油而对生长无负面影响, 但会降低肌肉中二十二碳六烯酸(DHA)和二十碳五烯酸(EPA)的含量。Xu等[62]报道高首鲟可去饱和和延长饲料中的高不饱和脂肪酸(18﹕2n-6、18﹕3n-3)。Sener等[63]以豆油和葵花油完全替代俄罗斯鲟 [(27.23±0.98) g]饲料中的鱼油, 以SGR为评估指标, 葵花油可完全替代鱼油对生长无负面影响。Sener等[64]发现用玉米油和葵花油混合油替代50%鱼油, 以体增重为评价指标,对俄罗斯鲟[(143.89±2.84) g]无负面影响。Giovanni等[65]在以螺旋藻粉完全替代鱼粉的情况下, 再用玉米油和豆油完全分别完全替代鱼油, 发现对高首鲟(17.5 g)的生长表现无负面影响, 但会降低肌肉中的DHA和EPA。关于鲟替代脂肪源的研究进展见表 6。综上所述, 在短期养殖过程中(< 90d), 豆油、玉米油、亚麻油、葵花油、加拿大菜油和螺旋藻油等植物脂肪源可以完全替代鱼油而对生长无负面影响, 甚至生长表现优于鱼油, 如在达氏鳇的研究中, 加拿大菜油在低脂和高脂水平下完全替代鱼油生长快于鱼油组[66]。但能否真正完全替代鱼油,还需要进行长期实验验证(>360d)。另外, 俄罗斯鲟的研究, 混合脂肪源的替代生长效果要优于单一脂肪源替代效果, 更多的研究应集中在混合脂肪源的研究中[67]。这是因为以上这些植物油中富含n-3多不饱和脂肪酸(亚油酸和亚麻酸), 能满足鲟必需脂肪酸的需求, 但植物油替代鱼油会导致鲟肌肉组织中的高不饱和脂肪酸(HUFA)明显下降, 这是急需研究和解决的问题。

表 5 鲟脂肪需求及脂肪酸的需求量Tab. 5 Requirements of dietary lipid and fatty acid in sturgeons
1.3 能量收支
鱼类的一切生命活动都离不开能量, 饲料中的能量需求主要来自于饲料中蛋白质、脂肪和碳水化合物。影响鱼类能量代谢的因素有水温、养殖密度、不同生长阶段、饲料蛋白水平和投喂水平等[15, 72, 73]。国内外有关鲟能量学的研究只有零星报道, Cui等[74]用活水蚓饲喂不同体重(2.4—22.5 g)的高首鲟来研究摄食水平(0、2%、4%、6%、8%初始体重和饱食)和鱼体规格对能量收支的影响。无论鲟体重多少, 特定生长率都会随着摄食水平线性增加, 饲料转化效率在饱食时最大。可消化能占总摄入能量的94.8%—96.8%, 饱食投喂时, 64.9%的代谢能用于代谢消耗, 35.1%的代谢能用于生长, 且不受鱼体规格的影响。Guo等[15]研究7种蛋白水平(250、300、350、350、400、450、500和550 g/kg)对初重为25.1 g的杂交鲟(西伯利亚鲟×俄罗斯鲟)能量收支的影响, 生长实验表明, 当饲料蛋白水平在250—400 g/kg时, 特定生长率会随之上升, 之后就显著下降, 摄食量会随着饲料蛋白水平的升高而减少, 在鲟摄食350 g/kg蛋白水平的饲料时, 能量收支模型为: C摄入能量(100%)=F粪便能量(35.08%)+U尿能量(4.53)+R代谢能(34.1%)+G生长净能(26.23%)。邢浩春等[75]研究了在低蛋白含量(28%)不同浓度维生素B6对施氏鲟幼鲟(34.7 g)能量收支的影响, 结果表明45—60 mg/kg维生素B6饲料转化效率最高, 特定生长率最高, 45和60 mg/kg维生素B6能量收支方程分别为: 100C=21.3F+11.5U+46.3R+21.0G和100C=19.7F+10.3U+52.7R+17.3G,该研究与郭文英[76]鲟品种不同, 郭文英的研究对象是杂交鲟, 该研究使用的是施氏鲟, 在郭文英[76]的研究中蛋白水平为25%时, 生长能分配率22.6%, 高于该研究的最高值21.0%, 这可能由于鲟品种不同而引起的。也可能是由于该研究中饲料配方中鱼粉的用量(19%)低于郭文英[76]鱼粉用量的最低值(28.2%)导致的。郭文英[76]研究α-淀粉水平(9%—25%)对西伯利亚鲟能量收支的影响, 以鱼粉(15%)和膨化大豆(16%)为主要蛋白源, 生长能分配率随着淀粉水平的增加而增加, 为31.94%—40.31%,生长速度在21%和25%的添加水平下最高。
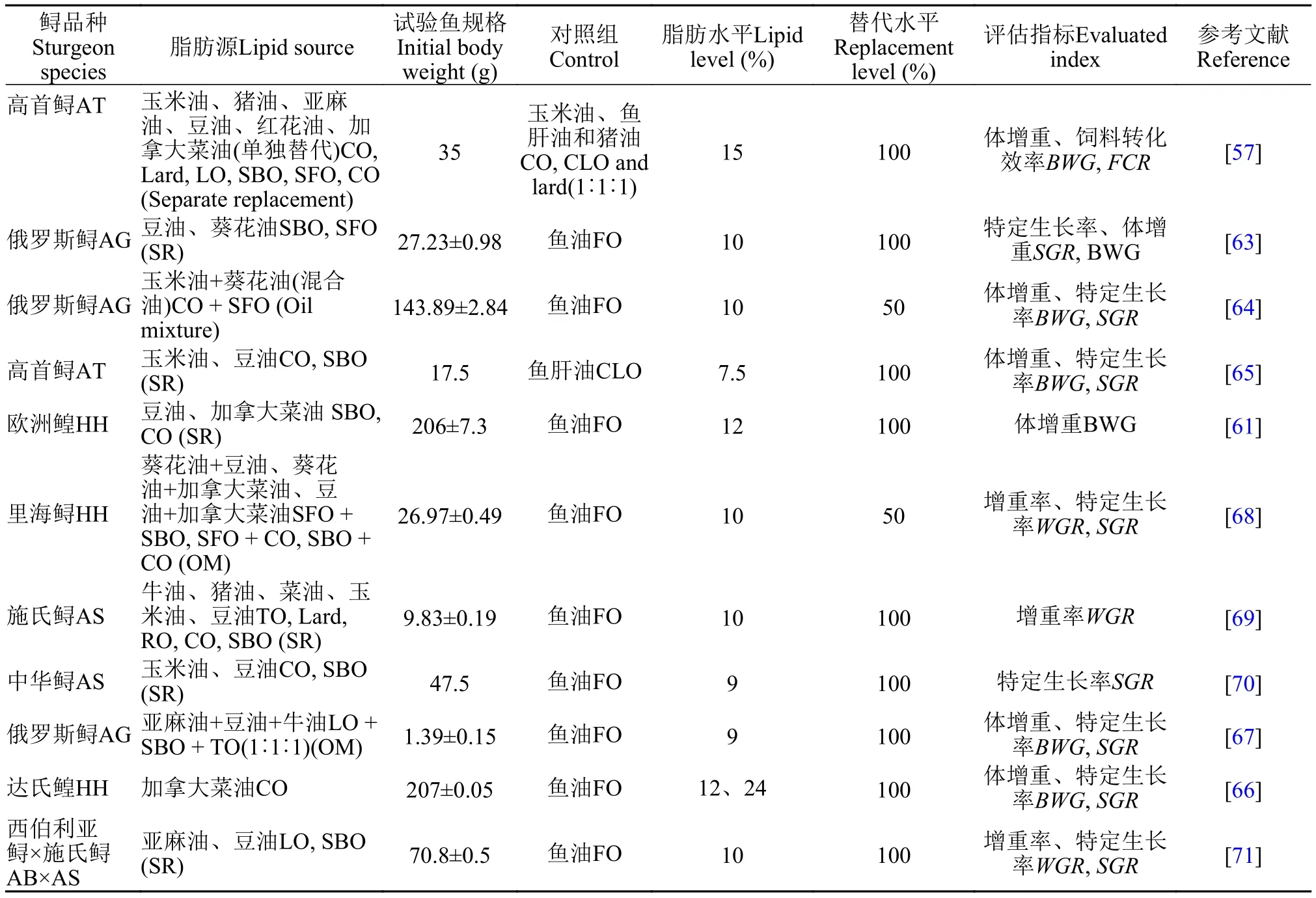
表 6 鲟饲料中其他脂肪源替代鱼油量Tab. 6 The replacement amount of dietary fish oil by other lipid sources in sturgeons
1.4 碳水化合物的营养需求
目前关于鲟适宜碳水化合物的需求研究较少。Fynn等[77]用含不同水平D-葡萄糖(0、7%、14%、21%、28%和35%)饲喂高首鲟8周, 以增重率为评估指标, 得出饲料葡萄糖的适宜添加水平为14%—35%。为了探究高首鲟合适的糖源, Hung等[78]配制8种等氮等脂但糖源不同(麦芽糖、葡萄糖、生玉米淀粉、糊精、蔗糖、乳糖、果糖和纤维素)的配合饲料饲喂鲟8周, 体增重和饲料效率按上述糖源顺序逐渐下降, 得出高首鲟能较好地利用麦芽糖和葡萄糖, 而对蔗糖、乳糖和果糖的利用较差。分析原因可能是鲟消化道里的蔗糖酶和乳糖酶活性较低, 从而对这两种糖源的消化率较差[79];其次, 也可能是高首鲟对蔗糖、乳糖和果糖的耐受力较差, Hung等[80]表明饲喂这3种糖源的高首鲟肝细胞的细胞质回缩且胞间糖原减少, 后肠肠腔水分较多, 与哺乳动物腹泻类似。同样对于高首鲟,Lin等[81]发现对淀粉的利用效果优于葡萄糖, 高首鲟对淀粉和葡萄糖利用效果的差异可能来自于2个报道中饲料配方、饲料蛋白水平和投喂频率的不同。而在施氏鲟的研究中, 其最适的糖源为糊精和α淀粉, 对果糖的利用最差[82], Yazdani等[83]也表明西伯利亚鲟可较好利用糊精(10%—20%)从而减少饲料中鱼油和植物油的用量。另外, 投喂频率对鲟利用碳水化合物也会产生影响。Lin等[81]采用连续投喂和1d两次投喂两种方式饲喂糖源为淀粉和葡萄糖的2种高首鲟饲料, 发现投喂方式对鲟利用碳水化合物的影响大于碳水化合物源的影响, 且连续投喂的特定生长率和饲料效率远高于1 d投喂两次。这与鲟的摄食习性有关, 鲟是下位口, 且为少量多次摄食, 摄食速度较慢。因此, 不同鲟品种利用碳水化合物源的能力不同, 根据目前的文献记载,鲟对果糖的利用较差, 建议鲟饲料中不要添加果糖。尽管鲟可较好地利用多糖如糊精和α淀粉, 但对生淀粉利用能力较差[84]。
饲料脂肪水平也会影响鲟利用碳水化合物。Mohseni等[85]采用正交实验法设计4种饲料蛋白水平(35%、40%、45%和50%)×4种碳脂比(碳水化合物﹕脂肪, C﹕L, 0.8、1.1、1.4和1.7)共16种饲料, 发现最适宜碳脂比(1.4)可有效节约蛋白质, 饲喂35%蛋白质和碳脂比为1.4的饲料鲟的特定生长率、饲料效率和蛋白质效率要高于40%蛋白和碳脂比为0.8以及50%蛋白水平及碳脂比为0.8和1.7。综上所述, 不同鲟品种对不同糖源的利用效果不同,高首鲟能较好地利用葡萄糖、麦芽糖和淀粉, 施氏鲟和西伯利亚鲟的最适糖源是糊精和α淀粉。
1.5 其他营养素的需求
维生素是维持鱼类生长和发育所必需的一类低分子有机化合物, 这类物质不能在体内合成或者合成较少, 必须由食物提供, 维生素对维持鱼类正常生理代谢有着极其重要的作用。关于鲟对维生素需求研究相对较少。鲟作为一群古老鱼类, 据报道, 具有自身合成维生素C(Vc)的能力[86]。这是因为鲟肾脏中含有将葡萄糖转化为L-维生素C的L-古洛糖酸内酯氧化酶[87]。这一观点在西伯利亚鲟中得到了验证, Moreau等[88]报道西伯利亚幼鲟[(25.5±0.5) g]在摄入无维生素C的饲料3个月后并没有出现VC缺乏症, 且生长表现与VC添加组(300 mg/kg饲料)无显著性差异。在杂交鲟(西伯利亚鲟×小体鲟)、达氏鳇也得到了同样的结果[89, 90]。而Desimira等[91]在闪光鲟(155 g)的研究中, 发现与高剂量VC添加组(800 mg/kg)和无VC组相比较, 低剂量的Vc(200—400 mg/kg)可显著地促进鲟的生长。Xie等[92]发现添加500 mg/kg的VC可增强西伯利亚鲟的免疫力。因此, 建议在鲟幼鱼阶段的饲料中添加适量Vc, 增强免疫力。Wen等[93]以体增重为评价指标, 发现施氏鲟对维生素A的需求量为923 IU/kg饲料。Amlashi等[94]以特定生长率为评估指标, 达氏鳇对饲料维生素E的需求量为25—400 mg/kg。鲟对维生素的需求汇总见表 7, 多数B族维生素需求的研究未见报道, 如维生素B1、B2、B12、烟酸、泛酸、生物素等。
最近几年对矿物盐的报道越来越多。矿物盐是构成鱼体组织的重要组分, 同时也是维持渗透压和酸碱平衡等正常生理代谢不可缺少的营养素。磷是水产动物不可或缺的营养元素, 磷缺乏会导致鱼类骨骼和鳞片矿化不良, 水产动物摄入过量的磷会导致磷的排放增加, 因此很有必要研究水产动物磷的需求[102]。Xu等[103]以磷酸二氢钠为磷源配制不同磷水平(0.12%—1.38%)的西伯利亚鲟饲料, 以增重率和全鱼磷含量为评价指标, 得出饲料适宜磷水平为0.5%—0.87%。Jin等[104]以玉米淀粉为载体给杂交鲟(达氏鳇×施氏鲟)灌喂不同剂量的磷酸二氢钙(0、100、200和400 mg/kg体重), 发现杂交鲟能较好地吸收磷源, 在灌喂4h后血磷就达到峰值,以磷的排泄率为评价指标, 磷的适宜添加水平为100 mg/kg体重(饲料中水平为1%)。
铜是水产动物必需的微量元素, 是许多酶的辅助因子, 如细胞色素氧化酶、Cu-Zn超氧化物歧化酶等(Cu-Zn SOD)[105]。Mohseni等[106]以CuSO4为铜源, 配制不同铜水平(1.1—195 mg/kg饲料)的达氏鳇饲料[(8.49±0.32) g], 以体增重为评价指标, 适宜的铜添加量为10.3—13.1 mg/kg饲料。在俄罗斯鲟(11.85 g)的研究中, 同样以CuSO4为铜源, 配制0.3—16.1 mg/kg的饲料, 以增重率、全鱼铜含量、肝脏Cu-Zn SOD和血浆铜蓝蛋白为评价指标, 饲料中适宜铜添加量为7—8 mg/kg饲料[107]。Wang等[108]比较俄罗斯鲟对不同水平的铜源的利用情况, 发现蛋氨酸铜和纳米氧化铜的生物利用度是CuSO4的1.5—2倍, 鲟对蛋氨酸铜、纳米氧化铜和CuSO4的需求量分别为5、5和8 mg/kg。Moazenzadeh等[109]以CuSO4为铜源, 配制不同铜梯度(1.8—28.3 mg/kg)的西伯利亚鲟[(27.57±0.24) g]饲料, 以特定生长率、全鱼蛋白质含量和超氧化物歧化酶活性为评价指标, 鲟饲料中铜的需求量分别是9.51、9.58和16.10 mg/kg。锌也是非常重要的必需微量元素, 它是200多种酶的辅助因子, 包括碱性磷酸酶、DNA聚合物和Cu-Zn SOD等, 参与各种生命活动[110]。Moazenzadeh等[110, 111]以硫酸锌为锌源, 配制不同锌水平(14.7—46.4 mg/kg)的西伯利亚鲟[(26.52±0.94) g]饲料, 以终末体重和肝脏锌含量为评价指标, 鲟饲料适宜的锌添加量为29.15 mg/kg和34.60 mg/kg。Arshad等[112]以L-蛋氨酸硒为硒源, 添加不同水平的硒(1.26—20.26 mg/kg)饲喂达氏鳇[(3.5±2.0) g] 8周,以体增重和特定生长率为评估指标, 得出饲料中适宜硒添加水平为11.56—20.26 mg/kg。鱼类对于不同来源的硒生物利用度不同, 如鲤对纳米硒的利用率高于亚硒酸钠和蛋氨酸硒, 同样, 虹鳟对酵母硒的生物利用度高于亚硒酸钠[113]。关于鲟对矿物盐的需求见表 8。其他的矿物盐还未见报道, 如钙、镁、铁、锰、钴和碘等。
1.6 饲料添加剂的研究进展
功能性饲料添加剂是一种具有动物保健作用的饲料添加剂, 具有改善动物生长性能、增加免疫力和提高抗应激能力等功效。目前应用于水产中的饲料添加剂主要包括益生菌、益生元、合生元、免疫增强剂、有机酸、核苷酸及中草药提取物等[116]。
Mohseni等[117]在达氏鳇饲料中添加不同梯度(0—1200 mg/kg)的L-肉碱, 发现添加剂量为300 mg/kg可显著促进鱼体的生长。Jalali等[118]在达氏鳇饲料中添加0、2.0、4.0和6.0 g/kg的海藻酸(Ergosan),以特定生长率和增重率为评价指标, 海藻酸的适宜添加剂量为4%—6%。Esterabadi等[119]研究益生元(Immunogen)对达氏鳇生长的影响, 发现饲料中添加0.5%—1%的益生元可显著促进鱼体生长。同样在达氏鳇的研究中, Hossein等[120]添加2%的灭活酿酒酵母可显著提高特定生长率、增重率及增加肠道中乳酸菌的数量。Askarian等[121]研究了来自达氏鳇消化道中的弯曲乳杆菌和波斯鲟胃肠道中的肠系膜明串珠菌两种乳酸菌对波斯鲟和达氏鳇生长、消化酶活性和肠道微生物的影响, 发现达氏鳇摄入含有9×109cfu/g饲料的弯曲乳杆菌可获得最高的特定生长率、存活率和消化酶(淀粉酶、蛋白酶和脂肪酶)活性, 而在波斯鲟中, 添加2×109cfu/g饲料的肠系膜明串珠菌也可取得同样的效果。Xu等[122]发现在杂交鲟(施氏鲟×达氏鳇)饲料中添加1%丙氨酰-谷氨酰胺和0.9%—1.5%谷氨酰胺可显著促进鲟的生长, 降低饲料系数和提高肠道消化酶活。同样在杂交鲟(施氏鲟×西伯利亚鲟)的研究中, 添加1%的丙氨酰谷氨酰胺可促进鱼体生长[123]。表 9按归纳总结近年来饲料添加剂在鲟上的研究, 为配制鲟优质高效饲料提供参考依据。
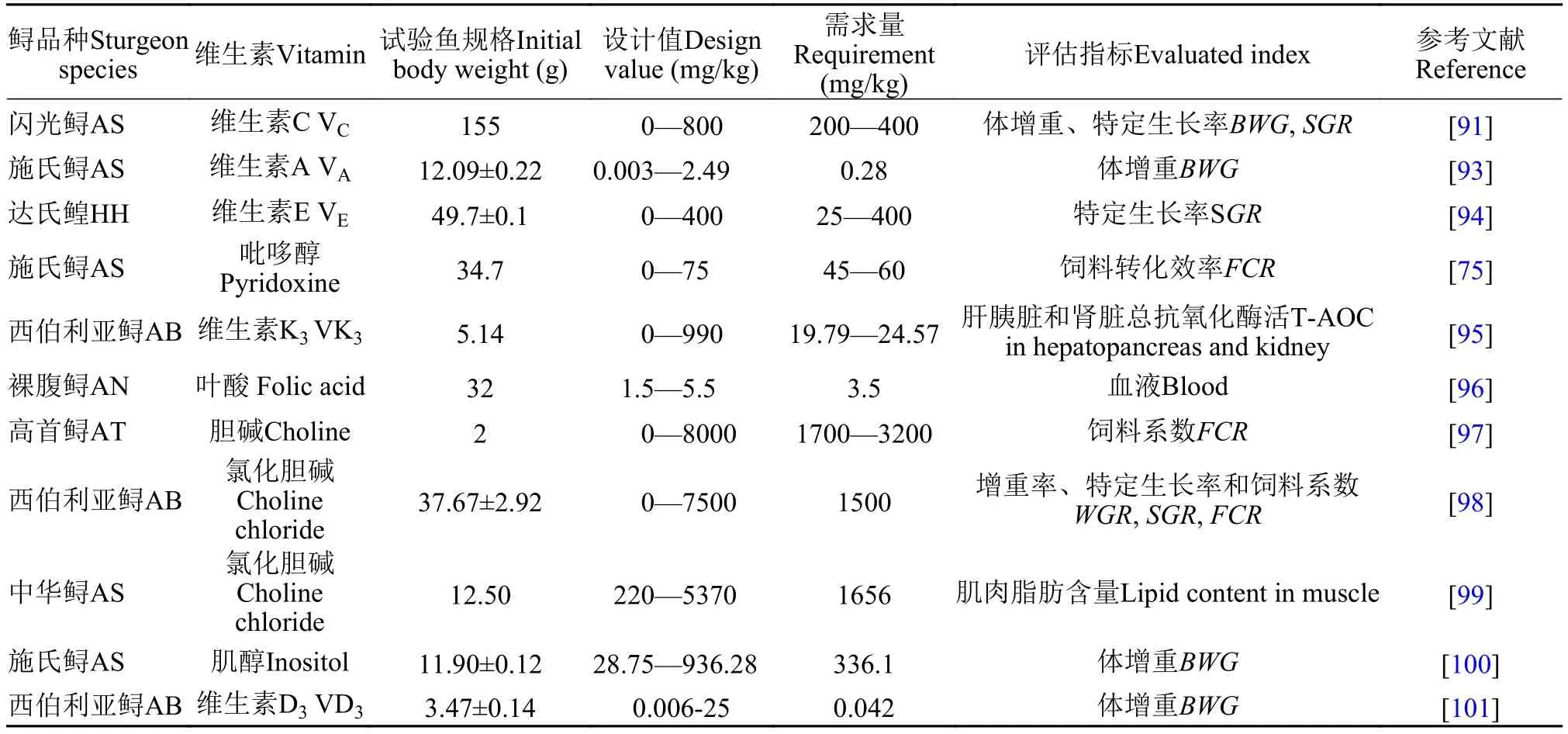
表 7 鲟对维生素的需求量Tab. 7 The requirement of dietary vitamin in sturgeons
2 总结与展望
近几年, 虽然关于鲟的营养学研究越来越多,但每种鲟的营养需求还需要进一步完善和扩充, 其中, 蛋白质需求在鲟中的报道并不集中在某一品种上, 在波斯鲟、匙吻鲟和达氏鳇等中均有报道, 需求量总体在40%左右。在蛋白源的替代研究中, 研究对象主要是西伯利亚鲟, 植物蛋白源和动物蛋白源均可被利用。不同鲟品种对脂肪需求不同, 需求量为11.1%—35.7%, 且鲟能较好地利用植物油; 碳水化合物的需求量为10%—35%; 关于矿物盐、维生素、添加剂的需求在鲟中均有报道。本文总结了鲟营养与饲料学相关的几乎所有文献, 为鲟精准营养需求提供了大量数据, 为鲟配合饲料的进一步开发提供科学依据。
但仍有一些问题需要解决: (1)鲟氨基酸需求的研究相对缺乏, 需要进一步的研究; (2)对鲟的研究均停留在幼鱼阶段, 很有必要研究其他生长阶段的营养需求; (3)关于维生素和矿物盐的需求还不全面, 需进一步的研究; (4)关于营养素对鲟肌肉品质和风味的影响较少, 只集中在全鱼和肌肉的营养成分分析, 需进一步研究营养素对肌肉物理和化学特性的影响; (5)需要比较不同品种的鲟对不同营养素需求的差异, 应用组学进行相关分子机制的研究。
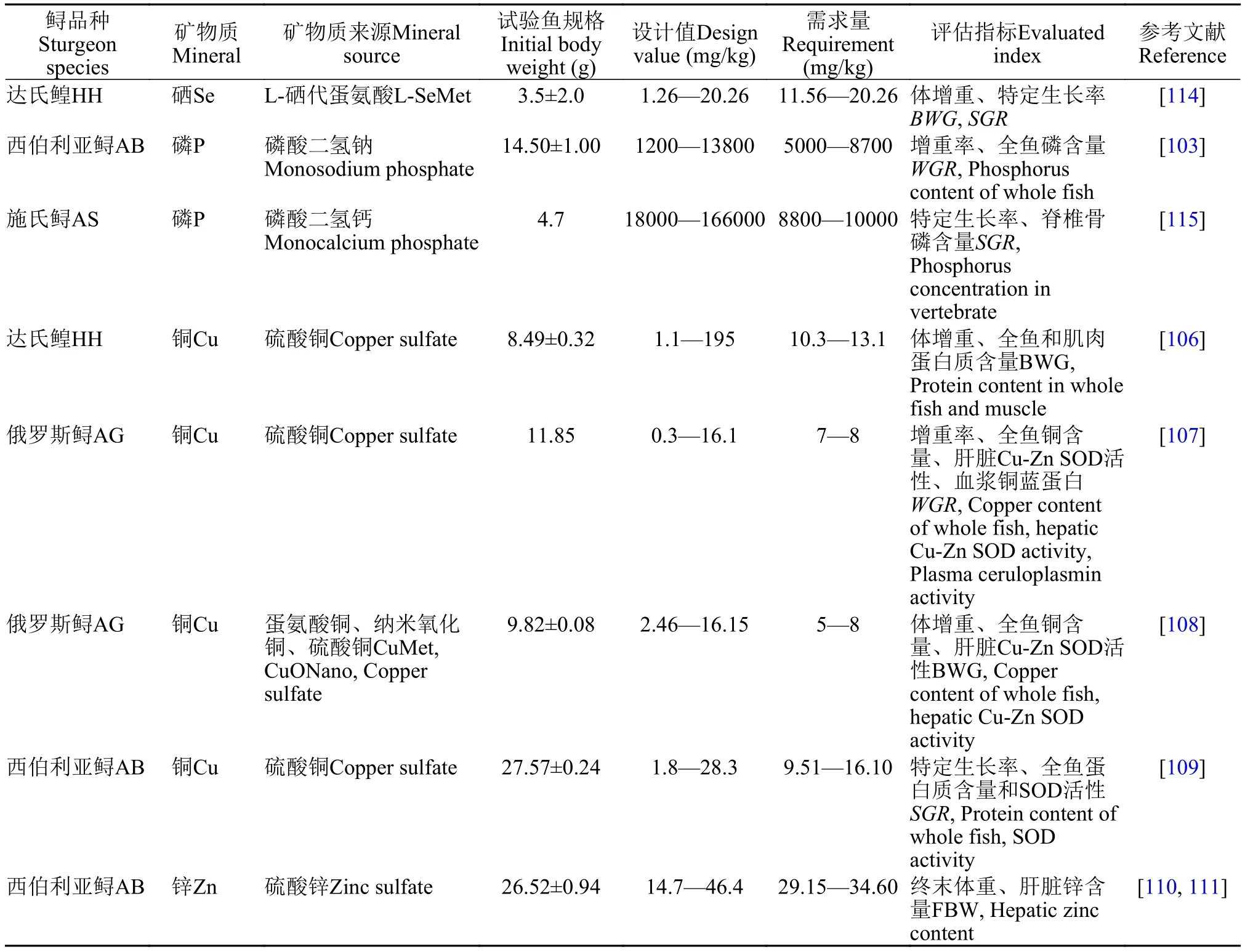
表 8 鲟对矿物盐的需求量Tab. 8 The requirement of dietary mineral in sturgeons

参Reference 献文]]]]]]]]]]]]]]]]]]考[117[118[119[121[121[122[122[123[124[125[126 R, SR[120[127[128[129[130[131[132 R, FCR zymes R, Digestive R, PE idan R, Digestive Serum zymes R t e en Fructooligosaccharide R, SG e en R活tiox SG R R R R, FC R, FC酶 an R R, FC R,WG Phagocytic activity,R, FC R, Digestiv, WG R, Digestiv R糖果氧聚SG数WGSG化抗评aluated index系 率R, WG SG R R, WG WG R, WG R R SR酶R, WG SG料SG SG长数SG SG率存mes性活mes活物率标血SG率饲SG清率系率性SG率S); 低生指 、重化、SG活率重 率料重重 率重和XO估 增率性酶数 性 定饲长氧脏增 长长ability、活化enzy酶特增增过增效生生系、化enzy生率活、Ev肝酶、、 、率酶消料质消、和体Complement activity, CA定率 定率 定率 率长化白和长化和 率4、 性饲特特特T生长长消长消数长蛋数 重 活定生、、生、系生生系生、增定率特定定定定特率 长率率料 料 、C3、C补 特特长特长饲 饲特生率长体生、生、定生、 活定补 性定率定 特 率存清 活ylan-oligosaccharides (A血Co特mplement C3, C4, Hepatic and serum特特重 重增增噬吞inox s Arab—1%量糖1.5—1.2%加4—%聚 sturgeon—3%适Optimal level 添—2%g 300 mg/kg木6 g/kg适 additive in量2%109 cfu/g 109 cfu/g 1%1%0 mg/kg 0.35 1%0.2%10%基加 宜0.5%9×2×0.9—0.9%25 1.5%1%糖添伯8宜g拉的剂); 阿加 围09 cfu/g 09 cfu/g n in Tab. 1—添 范00 mg/k 2%2%00 mg/k%料 值1 2 6 g/kg 4%2%1%1.5%1%1.5%0、10 0.5%2%3%0.3%2%10饲计0—0—0—0—0—0—0—0—0—0—0—0—0—0—9 鲟设Design value 109—9×1 2×109—9×1 2×esenteroides (LM表Optimal level of dietary e same as show Tab. 9 试itial body weight c m格规 g 2 g 5 g g g g osto鱼0 mg0—0.56 g 1.03 mg 2.06 mg 0.18 g 0.18 g 0.18 g 0.5 0.14 g 0.38 g验10 41.7±1.8 8.71±0.0.44±LC50 11.10±LM40 tamine22.38±.30±22 0.42±0.0.38±.38±25.9±0.9 10.2±0.8.57±.16±20.1±0.8 40.8±6 g SM32 22 Leucon 12 30 In wder5.5 g菌珠.16±串系8 Other abbreviations are th明er’s yeast-膜yl-L度表聚添Additive e complex tic Alanyl-G C); 肠 1—(平lu剂L-Alan tide mixture合均加写ine tus (L同Inactive brew酰简nogen e L-carnitin胺ine胺糖e degree of cleo氨他菌Ergosan聚Nu母mu酰木erag珠 氨Multienzym物-谷S Beta-glucan 糖元酵glutam菌), 其-L串Glutam Im基 av酶合polymerization 30 FO杆Garlic po酒Yeast prebio明-谷胺酰in胺 糖e S合混糖 聚生粉A T元碱酸酿膜乳酰酰酰氨伯XO混酸果粉 益藻acillus curva L-肉藻生活曲系氨 氨 氨L-谷拉 种苷阿30)A聚蒜 母旋Beta-葡菌Catalase (C谷HH灭ctob AP肠HH海La杆酶谷ecies HH弯亚Glutam鲟鳇种鳇Sturgeon sp AB利鳇乳氢丙品氏氏伯 氏 鲟曲化鲟HH HH益达达西 达 亚HH多HH核AS低AS大AP HH酵HH螺: 弯氧鳇鳇鳇鳇 鳇鲟鲟斯氏×H鲟×H×H×B×H氏×H鲟氏×A鲟 利 鳇鳇鲟鲟鲟鳇鳇注); 过氏氏氏氏氏氏×H伯 氏氏光光斯氏氏达达达达达波施A S施A S施A S施A S西 达达闪闪波达达O S(F
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
中国生殖健康(2020年5期)2021-01-18
小读者(2019年20期)2020-01-04
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
红领巾·成长(2019年3期)2019-04-16
中国生殖健康(2018年5期)2018-11-06
商周刊(2017年6期)2017-08-22
现代营销·信息版(2017年2期)2017-03-17
