
Acetyl-11-keto-β-boswellic acid inhibits proliferation and induces apoptosis of gastric cancer cells through the phosphatase and tensin homolog /Akt/ cyclooxygenase-2 signaling pathway
2020-12-10 04:08MengXueSunXiaoPuHePeiYunHuangQiQiWeiHaoSunGaoShuangLiuJieHua
World Journal of Gastroenterology 2020年38期
Meng-Xue Sun, Xiao-Pu He, Pei-Yun Huang, Qi Qi, Wei-Hao Sun, Gao-Shuang Liu, Jie Hua
Abstract
Key Words: Acetyl-11-keto-β-boswellic acid; Gastric cancer; Cell proliferation; Apoptosis; Cyclooxygenase-2; Tumor xenograft
INTRODUCTION
Gastric cancer is the fourth most common cancer and second leading cause of cancerrelated mortality in the world[1]. It is a multifactorial disease characterized by being highly drug resistant, and apoptotic deficiency could be the obstacle that hampers the efficacy of most anti-cancer agents. Currently, the main treatments include surgery, radiotherapy, and chemotherapy. However, the prognosis of gastric cancer is still poor; thus, it is necessary to find new drugs to treat gastric cancer.
Acetyl-11-keto-β-boswellic acid (AKBA) is a pentacyclic terpenoid extracted from the gum ayurvedic therapeutic plantBoswellia serrata. The molecular structure of AKBA is shown in Figure 1. AKBA is anti-inflammatory agent that exhibits potent cytotoxic activities against various types of tumors, such as hepatocellular carcinoma[2], glioblastoma[3], prostate cancer[4], lung carcinogenesis[5], colon cancer[6,7], and leukemia[8,9]. However, the efficacy and mechanism of action of AKBA in treating gastric cancer have not been studied. This study is the first attempt to explore the efficacy and mechanism of action of AKBA in gastric cancer.

Figure 1 Molecular structure of acetyl-11-keto-β-boswellic acid.
Cyclooxygenase (COX) catalyzes the formation of prostaglandin and other eicosanoids from arachidonic acid, which has been considered as a potential target for the prevention and treatment of cancer. It tends to be highly expressed in inflammation and tumor tissues but is undetectable in the majority of normal tissues[10-12]. COX-2, the mitogen-inducible isoform, is constitutively expressed in gastric cancer. It has been shown that overexpression of COX-2 is directly or indirectly involved in the carcinogenesis and development of gastric cancer, and is closely related to its prognosis[13]. However, the molecular mechanisms underlying the aberrant expression of COX-2 in gastric cancer remain unclear. Regulation of COX-2 expression depends on different cellular pathways, involving both transcriptional and post-transcriptional mechanisms[14]. Among all upstream regulatory factors, activation of the PI3K/Akt pathway is important for COX-2 up-regulation[15-18]. PTEN (phosphate and tension homology deleted on chromosome ten) is a tumor suppressor with phosphatase activity that can regulate cell survival, proliferation, and energy metabolism[19,20]. Many tumor tissues present with deficiency of PTEN[21]. PTEN acts as a lipid phosphatase to dephosphorylate phosphatidylinositol (3-5)-trisphosphate, antagonizing the PI3K/Akt pathway[22]. Recent studies have demonstrated that the PTEN/Akt pathway can regulate COX-2 expression in human cervical cancer, gallbladder cancer, and colorectal cancer[23-25].
In this study, human gastric cancer cell lines BGC823 and SGC7901, in which COX-2 was found to be overexpressed[10,13,26,27], were used to investigate the anti-tumor effects of AKBA on human gastric cancerin vitroandin vivo. Additionally, the expression of proliferating cell nuclear antigen (PCNA), Bcl-2, Bax, COX-2, PTEN, and p-Akt was detected to elucidate the possible mechanism underlying the anti-tumor effects of AKBA against gastric cancer.
MATERIALS AND METHODS
Reagents
AKBA (C32H48O5, CAS: 67416-61-9, purity: > 98%), dimethylsulfoxide (DMSO), and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenylformazan (MTT) were purchased from Sigma-Aldrich (St. Louis, MO, United States). Roswell Park Memorial Institute (RPMI)-1640 medium, fetal bovine serum, and penicillin/streptomycin were purchased from Gibco BRL (Grand Island, NY, United States). Hoechst 33342 and apoptosis detection kit were purchased from Beyotime Institute of Biotechnology (Nantong, China). PTEN inhibitor bpv (HOpic) and Akt inhibitor MK2206 were purchased from Selleck Chemicals (Houston, TX, United States). Other reagents were of analytic grade and obtained from Nanjing Chemical Reagent Co. (Nanjing, China), unless otherwise described. AKBA was dissolved in DMSO as a 2 mmol/L stock solution and stored at -20 °C.
Cell culture and treatments
Human poorly differentiated BGC823 and moderately differentiated SGC7901 gastric cancer cell lines were obtained from Shanghai Institute of Cell Biology (Shanghai, China). All cells were routinely cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. All the cells were kept at 37 °C in a humidified atmosphere of 5% CO2.
Methyl thiazolyl tetrazolium (MTT) assay
BGC823 and SGC7901 cells (4000-5000/well) were seeded into 96-well plates and incubated overnight in a 5% CO2atmosphere at 37 °C. The following day, the gastric cancer cells were treated with different concentrations of AKBA (0, 10, 20, 30, 40, and 50 µM) in serum-free conditions for 24, 48, or 72 h at 37 °C. Subsequently, 200 µL of MTT solution (0.5 mg/ML) was added to each well and incubated for 4 h at 37 °C. DMSO (200 µL) was then added to each well. The proliferation-inhibitory effects of each combination were assessed using a microplate reader (MJ Research Inc.) at 570 nm.
Flow cytometry
To detect the apoptosis rate of gastric cancer cells, we used the Annexin V fluorescein isothiocyanate (FITC) Apoptosis Detection Kit to measure apoptosisviaflow cytometry. BGC823 and SGC7901 cells were plated in six-well plates at 2 × 105cells per well. All the cells were treated with different concentrations of AKBA (0, 20, 30, or 40 µM) for 48 h. Detached cells were collected, combined with the trypsinized attached cells, and centrifuged at 1400 r/min, 4 °C, for 10 min. Cells were re-suspended in 1 mL of binding buffer and allowed to react with 10 µL of FITC-labeled Annexin V and 10 µL of PI for 15 min at room temperature in the dark. Afterward, the cells were analyzed on a flow cytometer (Becton Dickinson, San Jose, CA, United States). Apoptosis was assessed by Annexin V-FITC and PI staining.
Western blot analysis
BGC823 and SGC7901 cells were grown to sub-confluence in 100-mm dishes and cultured in serum-free medium for 24 h. The cells were treated with different concentrations of AKBA (0, 20, 30, or 40 µM) in serum-free conditions for 48 h. In another experiment, the cells were pre-treated with the PTEN inhibitor bpv (HOpic, 17.4 ng/ML) or Akt inhibitor MK2206 (38.4 ng/ML) for 30 min and then with AKBA (40 µM) for 48 h. The extraction of proteins from cells and Western blotting were performed as described previously[28,29]. Antibodies used include rabbit anti-PCNA, anti-Bcl-2, anti-Bax, anti-PTEN, anti-COX-2, and anti-reduced glyceraldehydephosphate dehydrogenase (GAPDH) (Abcam Inc., Cambridge, MA, United States), as well as anti-phospho (p)-Akt and anti-Akt (Sigma–Aldrich). Horseradish peroxidaseconjugated goat anti-rabbit IgG purchased from Abcam was used as secondary antibody. The relative expression of PCNA, Bcl-2, Bax, PTEN, and COX-2 was normalized to that of GAPDH. P-Akt was normalized to the total Akt level.
Wound healing assay
A wound healing assay was performed as described previously[30]. BGC823 and SGC7901 cells were seeded in six-well plates (7 × 105–8 × 105cells/well) to produce a con uent monolayer. The cell monolayer was scratched with a pipette tip and washed with phosphate-buffered saline. Cells were then incubated with different concentrations (20 µM) of AKBA. Scratch wounds were photographed under a phase contrast microscope (Leica, Nussloch, Germany) at 0, 48, and 72 h post treatment, respectively.
Anti-cancer activity in mice
The effect of AKBA was further evaluated in nude mice bearing xenografts of BGC823 and SGC-7901 cells. The use of all experimental animals was in compliance with the protocols set by Nanjing Medical University Institutional Animal Care and Use Committee. Female athymic BALB/c (nu+/nu+) mice (n= 20), aged 5 wk, were purchased from the Department of Laboratory Animal Center of Nanjing Medical University (Nanjing, China). The mice were kept in a specific pathogen-free conditional microenvironment (temperature 24 °C, humidity 60 ± 10%, and 12 h/12 h light/dark cycle). Then, 0.1 mL of cell suspension containing 5 × 106– 6 × 106BGC823 or SGC7901 cells was subcutaneously injected into the axillary space of the mice. When tumor volume reached approximately 100 mm3, mice were randomly divided into different groups (n= 5): AKBA (100 mg/kg) or control (equal volume of normal saline). Mice were observed daily for clinical symptoms. Tumor volume and body weight were measured every 2 d. The tumor volume was calculated as (shortest diameter)2× (longest diameter) × 0.5. At the end of treatment, nude mice were intraperitoneally injected with 3% pentobarbital sodium and were euthanized by excessive anesthesia at a dose of 90 mL/kg. The tumor tissue was weighed and excised for Western blot analysis.
Statistical analysis
All data from at least three independent experiments are represented as the mean ± SD and statistically analyzed by two-tailed Student’st-test or one-way analysis of variance with Dunnett’s multiple comparison tests. Statistical analyses were performed using GraphPad Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, United States).P< 0.05 was considered statistically significant.
RESULTS
AKBA inhibits proliferation and induces apoptosis of gastric cancer cells
MTT assay was performed to evaluate the survival of gastric cancer cells exposed to different therapeutics (0–50 µM for 24, 48, and 72 h). AKBA significantly inhibited the proliferation of BGC823 and SGC7901 cells in a dose- and time-dependent manner (Figure 2A and B). After 72 h of treatment with 50 µM AKBA, the proliferation of BGC823 and SGC7901 cells was almost completely blocked.
Annexin V-FITC/PI assay was also conducted to investigate the apoptosis induced by AKBA on the basis of flow cytometry. In the dual-parameter fluorescent dot plots, early apoptotic cells stained with Annexin V (Annexin V+/PI−, Q3), and late apoptotic cells stained with Annexin V and PI (Annexin V+/PI+, Q2) were counted (Figure 2C). AKBA induced the apoptosis of gastric cancer cells in a dose-dependent manner (Figure 2D).
To study the effect of AKBA on cell proliferation and apoptosis, the expression of PCNA (involved in proliferation), Bcl-2, and Bax (involved in apoptosis) in BGC-823 and SGC-7901 cells was evaluated by Western blot analysis. After 48 h of AKBA treatment, the expression of Bax was up-regulated and that of Bcl-2 and PCNA was downregulated in a dose-dependent manner in BGC-823 and SGC-7901 cells (Figure 2E-G). All of these data indicated that AKBA inhibits the proliferation and induces the apoptosis of gastric cancer cells.
Effects of AKBA on expression of PTEN, COX-2, and p-AKT in BGC-823 and SGC-7901 cells
PTEN, COX-2, and p-Akt expression in gastric cancer cells was detected by Western blot analysis. AKBA dose-dependently inhibited COX-2 expression and p-Akt expression, and increased PTEN expression in BGC-823 and SGC-7901 cells (Figure 3).
Effects of PTEN/Akt inhibitor on AKBA-induced downregulation of COX-2 expression in gastric cancer cells
To elucidate the signaling pathway involved in the down-regulation of COX-2 expression induced by AKBA, further studies were designed to determine the effect of PTEN and Akt inhibitors on COX-2 expression in gastric cancer cells. The PTEN inhibitor bpV (HOpic) blocked the high expression of PTEN induced by AKBA and reversed the low expression of p-Akt and COX-2 inhibited by AKBA (Figure 4). The Akt inhibitor MK2206 combined with AKBA downregulated the expression of p-Akt and COX-2, and the combined effect was better than that of AKBA alone. These results collectively indicated that AKBA elicited inhibition of proliferation and induction of apoptosis of gastric cancer cells may be mediated by down-regulation of COX-2 expression through the PTEN/Akt signaling pathway.
AKBA inhibits cell migration and wound closure of gastric cancer cells
We further examined the effect of AKBA on cell migration after monolayer wounding. When BGC823 and SGC7901 cell monolayers were wounded and treated with AKBA (20 µM) for 48 or 72 h, the AKBA-treated groups closed the gaps at a slower pace at 48 and 72 h , compared with the control group (Figure 5).
AKBA inhibits gastric xenograft tumor growth in vivo
The effect of AKBA on BGC823 and SGC7901 cell xenografts was examined (Figure 6). Twenty nude mice implanted with BGC823 or SGC7901 cells were treated daily with AKBA for 2 wk. No deaths occurred within that period. The average tumor volume of nude mice treated with AKBA was markedly lower than that in the control nude mice (Figure 6A and B). There was no significant difference in body weight between the control and AKBA-treated nude mice (Figure 6C). COX-2 protein was overexpressed in gastric cancer tissues of control nude mice (Figure 6D and E). The results indicated that AKBA reduced the expression of COX-2 in the tumor tissues of nude mice, suggesting that AKBA inhibits tumor growth in nude mice by down-regulating the expression of COX-2.
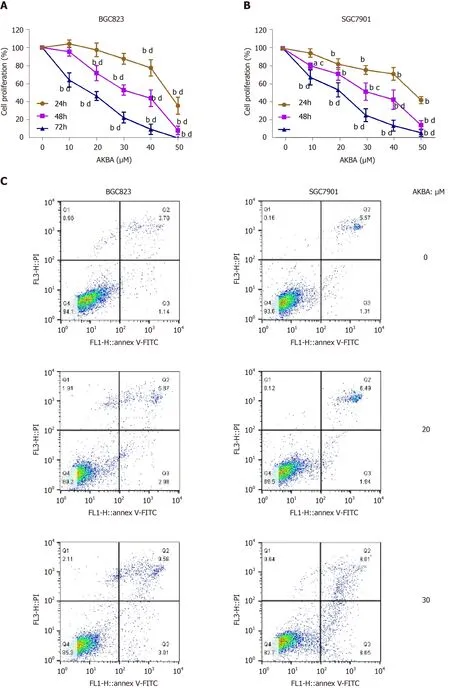
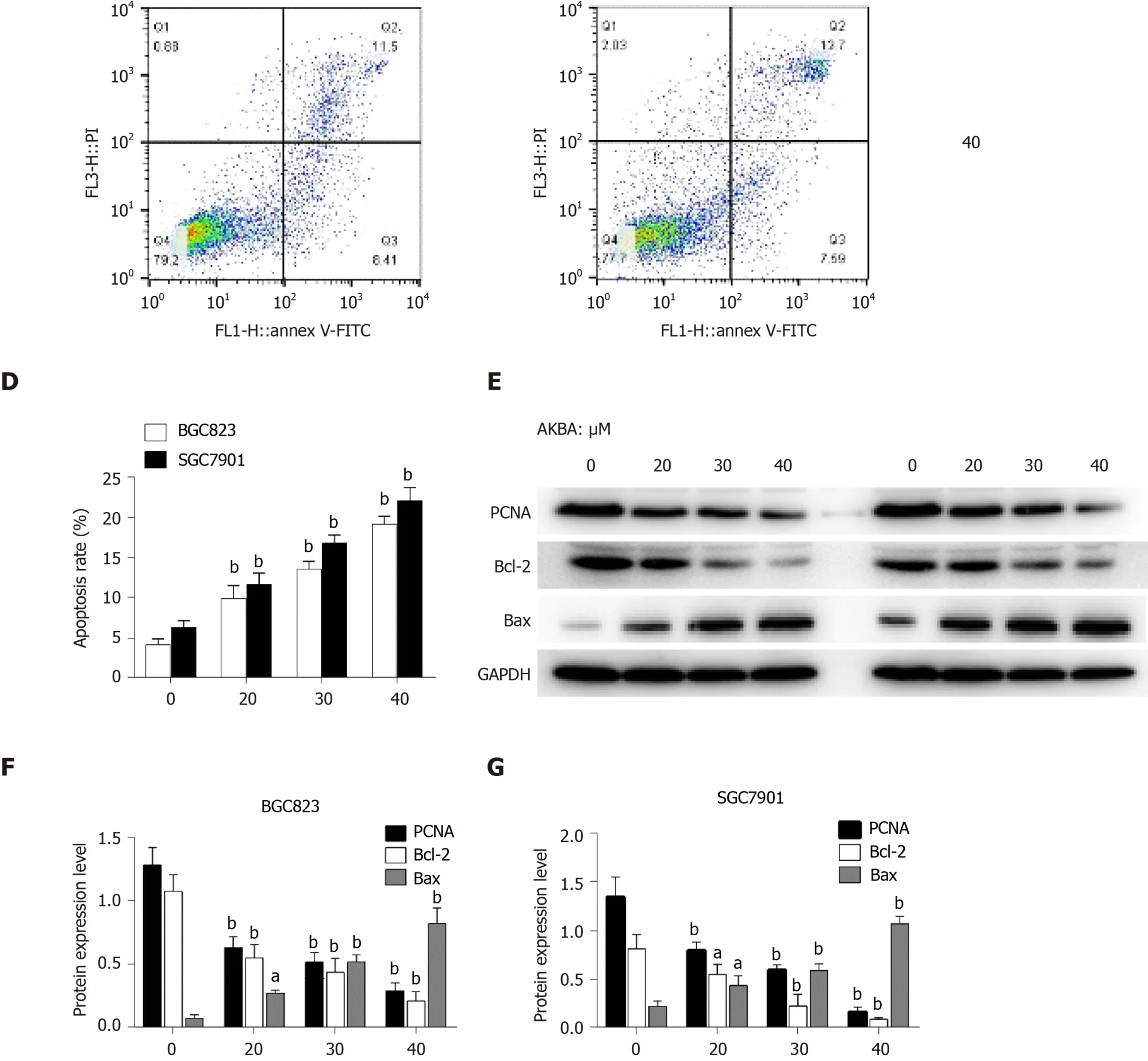
Figure 2 Acetyl-11-keto-β-boswellic acid inhibits the proliferation and induces the apoptosis of gastric cancer cells. A and B: Dose- and time-dependent effects of acetyl-11-keto-β-boswellic acid (AKBA) on BGC823 and SGC7901 cell proliferation; C: Flow cytometry-based annexin V-FITC/PI labeling of apoptotic cells; D: Histogram showing apoptosis rates; E: Expression levels of proliferating cell nuclear antigen (PCNA), Bcl-2, and Bax in BGC823 and SGC7901 cells measured via Western blot analysis after AKBA treatment; F and G: Histograms showing the relative expression levels of PCNA, Bcl-2, and Bax compared with GAPDH in BGC823 and SGC7901 cells. Each data point represents the mean ± SD from three independent experiments. aP < 0.05, bP < 0.01 vs control (0 µM). cP < 0.05, dP < 0.01 vs control (24 h). AKBA: Acetyl-11-keto-β-boswellic acid; PCNA: Proliferating cell nuclear antigen; GAPDH: Glyceraldehyde-phosphate dehydrogenase.
DISCUSSION
Gastric cancer is one of the most common malignancies with a high mortality rate. COX-2 is expressed in gastric cancer and is a factor involved in cancer cell proliferation and apoptosis, cancer invasiveness, and metastasis[31]. Studies have shown that COX-2 inhibition by selective COX-2 inhibitors or siRNA suppresses cell growth and leads to apoptosis in human gastric adenocarcinoma cells[29,32]. In this study, we demonstrated for the first time that AKBA can downregulate the expression of COX-2viathe PTEN/Akt signaling pathway in gastric cancer cells, and consequently induces the apoptosis and inhibits the proliferation of gastric cancer cells.
Previous studies have confirmed that AKBA can inhibit the proliferation and enhance the apoptosis of pancreatic cancer cellsin vitroby reducing COX-2 expression level[33]. The results of those studies are consistent with our finding that AKBA can significantly inhibit COX-2 expression in gastric cancer cells. However, the molecular mechanism by which AKBA down-regulates the expression of COX-2 in gastric cancer cells remains unknown.

Figure 3 Effects of acetyl-11-keto-β-boswellic acid on the expression of phosphatase and tensin homolog, cyclooxygenase-2, and p-Akt in gastric cancer cells. A: Expression levels of phosphatase and tensin homolog (PTEN), cyclooxygenase (COX)-2, and p-Akt in gastric cancer cells measured via Western blot analysis; B: Histograms showing the relative expression levels of PTEN and COX-2 compared with GAPDH, and p-Akt compared with Akt in gastric cancer cells. Each data point represents the mean ± SD from three independent experiments. aP < 0.05, bP < 0.01 vs control (0 µM). AKBA: Acetyl-11-keto-βboswellic acid; PTEN: Phosphatase and tensin homolog; COX-2: Cyclooxygenase-2; GAPDH: Glyceraldehyde-phosphate dehydrogenase.
Numerous studies have confirmed that the PI3K/Akt signaling pathway can induce apoptosis in a variety of cancer cells[34-37]. COX-2 expression can be induced by activating the PI3K/Akt signaling pathway in gastric cancer cells, thereby regulating the proliferation of gastric cancer cells[28]. PTEN deletion increases the activation of the PI3K/Akt pathway[22,38]. When the PTEN gene is mutated or lost, the Akt signaling pathway is activated, resulting in the proliferation of cells to form tumors[39]. miR-21 promotes cell proliferation by activating PTEN/Akt signaling in cancer cells[40]. CD2 selectively induces the expression of COX-2 through PTEN-mediated PI3K/Akt activation[41]. The selective COX-2 inhibitor celecoxib inhibits tumor growthviathe PTEN/PI3K/Akt pathway in an H22murine hepatocarcinoma model[17]. Our results are consistent with those previous findings. Our results showed that AKBA can effectively inhibit the proliferation and migration of gastric cancer cells and induce their apoptosis. Additionally, AKBA significantly inhibited the expression of PCNA, Bcl-2, and p-Akt, and increased Bax and PTEN expression in gastric cancer cells. To clarify the molecular mechanisms of the anti-cancer effect of AKBA on gastric cancer, we examined ethe xpression of proteins related to the PTEN/Akt/COX-2 pathwayin vitro(Figure 7). The PTEN inhibitor bpv (Hopic) blocked AKBA-induced PTEN expression, and reversed AKBA-induced downregulation of p-Akt and COX-2 expression. The Akt inhibitor MK2206 combined with AKBA downregulated the expression of p-Akt and COX-2, and the combined effect was better than that of AKBA alone.
In vitrodata show that AKBA inhibits the occurrence and development of gastric cancer by inducing apoptosis of gastric cancer cells. Our animal experiments verified the findings ofin vitroexperiments. Previous research confirmed that AKBA has an anti-tumor effect in nude mice[2-4,33,42,43]. Our results indicate that the tumor size of AKBA-treated nude mice is significantly smaller compared to the control group. AKBA significantly inhibits tumor growth in xenograft nude mice without causing significant weight loss, mortality, or other adverse effects.

Figure 4 Effects of phosphatase and tensin homolog /Akt inhibitor on acetyl-11-keto-β-boswellic acid-induced down-regulation of cyclooxygenase-2 expression in gastric cancer cells. Gastric cells were pre-treated with the phosphatase and tensin homolog (PTEN) inhibitor bpv (HOpic, 17.4 ng/mL, A and B) or Akt inhibitor MK2206 (38.4 ng/mL, C and D) for 30 min and then treated with acetyl-11-keto-β-boswellic acid (40 µM) for 48 h. A: Expression of PTEN, cyclooxygenase (COX)-2, and total and phosphorylated forms of Akt in gastric cancer cells detected by Western blot analysis. B: Histograms showing the relative expression of PTEN and COX-2 compared with GAPDH, and phospho-Akt (p-Akt) compared with total Akt, respectively. Each data point represents the mean ± SD from three independent experiments. aP < 0.05, bP < 0.01 vs Control; cP < 0.05, dP < 0.01 vs AKBA. AKBA: Acetyl-11-keto-β-boswellic acid; PTEN: Phosphatase and tensin homolog; COX-2: Cyclooxygenase-2; GAPDH: Glyceraldehyde-phosphate dehydrogenase.
CONCLUSION
In summary, these results suggest that AKBA inhibition of proliferation and induction of apoptosis of gastric cancer cells may be mediated by down-regulation of COX-2 expression through the PTEN/Akt signaling pathway.

Figure 5 Acetyl-11-keto-β-boswellic acid inhibits the monolayer wound healing of gastric cancer cells. A and B: Phase micrographs of BGC823 and SGC7901 cells at various times after monolayer wounding (original magnification, × 100); C and D: Quantification of cell migration using monolayer woundhealing assay. The gastric cancer cells were treated without (control) or with 20 µM acetyl-11-keto-β-boswellic acid. Each data point represents the mean ± SD from three independent experiments. aP < 0.05, bP < 0.01 vs control (0 µM). AKBA: Acetyl-11-keto-β-boswellic acid.

Figure 6 Acetyl-11-keto-β-boswellic acid inhibits gastric cancer tumor growth in vivo. Twenty nude mice were subcutaneously injected with BGC823 and SGC7901 cells. When tumor size was approximately 0.1 cm3, the nude mice were randomly divided into different groups (n = 5) to receive different treatments. A: Acetyl-11-keto-β-boswellic acid (AKBA) inhibits gastric cancer xenograft tumor growth in vivo; B: Tumor volume was measured, and the tumor growth curves were plotted; C: Body weight change in the nude mice treated with saline or AKBA; D: Western blot analysis of cyclooxygenase (COX)-2 protein expression in gastric cancer tissues of control and AKBA treated nude mice (100 mg/kg/d); E: Histogram representing the relative expression of COX-2 compared with that of GAPDH. aP < 0.05, bP < 0.01 vs control (saline). AKBA: Acetyl-11-keto-β-boswellic acid.
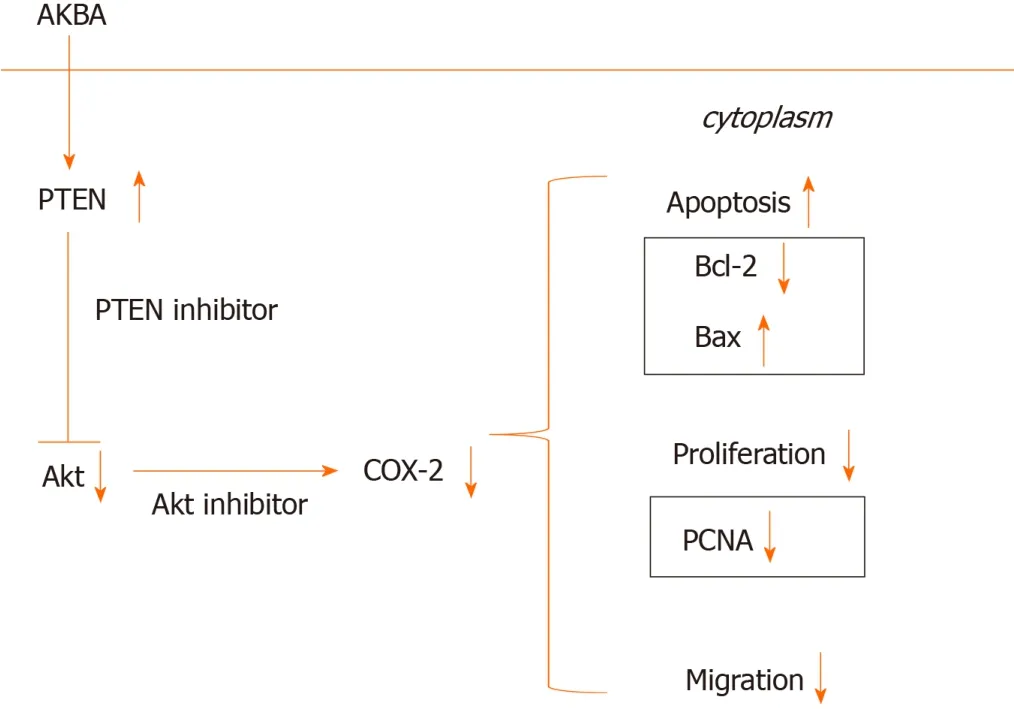
Figure 7 Hypothetical model of acetyl-11-keto-β-boswellic acid’s inhibition of proliferation of gastric cancer cells and induction of apoptosis. AKBA: Acetyl-11-keto-β-boswellic acid; PTEN: Phosphatase and tensin homolog; COX-2: Cyclooxygenase-2; PCNA: Proliferating cell nuclear antigen.
ARTICLE HIGHLIGHTS

Research perspectives
COX-2 is involved in the carcinogenesis and development of gastric cancer. AKBA is a promising drug for gastric cancer therapy.
 World Journal of Gastroenterology2020年38期
World Journal of Gastroenterology2020年38期
- World Journal of Gastroenterology的其它文章
- Role of betaine in liver disease-worth revisiting or has the die been cast?
- Management of an endoscopy center during the outbreak of COVID-19: Experience from West China Hospital
- Gastrointestinal complications after kidney transplantation
- Is vitamin D receptor a druggable target for non-alcoholic steatohepatitis?
- Endogenous motion of liver correlates to the severity of portal hypertension
- Longitudinal decrease in platelet counts as a surrogate marker of liver fibrosis