
ZAP蛋白在抗病毒天然免疫应答过程中的功能研究
2020-12-09 08:36:38陈纳张宝戈洪艳许李锋谢正露陈吉龙马燕梅
畜牧与兽医 2020年12期
陈纳,张宝戈,洪艳,许李锋,谢正露,陈吉龙,3,马燕梅*
(1. 福建农林大学动物科学学院/蜂学学院,福建 福州 350002; 2. 福建农林大学金山学院,福建 福州 350002;3. 中国科学院微生物研究所/中国科学院病原微生物与免疫学重点实验室,北京 100101)
机体免疫包括天然免疫和适应性免疫。天然免疫是机体在长期种系发育与进化过程中逐渐形成的一种先天性免疫防御功能,为宿主抵御病原微生物入侵的第一道防线,是当前免疫学前沿研究热点之一。宿主细胞与病毒长期共存过程中进化出一些具有抗病毒作用的因子称宿主限制因子,其可在病毒增殖的特定环节与病毒相互作用从而阻断病毒的复制。近年来,新型宿主限制因子ZAP蛋白(zinc-finger antiviral protein,锌指抗病毒蛋白)逐渐成为抗病毒天然免疫领域的研究热点,人们发现该蛋白在天然免疫应答中发挥重要作用,可通过多种途径直接或间接抑制病毒的复制,同时病毒也已进化出多种机制拮抗ZAP蛋白的抗病毒活性。本文对宿主ZAP蛋白与不同病毒互作机制进行综述,有助于从不同角度拓展人们对宿主抗病毒天然免疫与病毒拮抗宿主天然免疫两者的认识。
1 ZAP蛋白的发现及其结构特征
ZAP蛋白又称为PARP13、ZC3HAV1,是一种RNA结合蛋白,可调节特定mRNA的合成、稳定性和翻译[1]。2002年中国科学院Gao等[2]在筛选抗Moloney鼠白血病病毒(moloney murine leukemia virus,M-MLV)增殖的宿主因子时首次发现了ZAP蛋白,并将其鉴定为宿主限制性因子。人类ZAP蛋白主要存在两种剪接变异体,分别为长链的ZAP(ZAPL)和短链ZAP(ZAPS),2019年研究又发现了超长链的ZAP(ZAPXL)和中链的ZAP(ZAPM)两种新的存在形式[3-4]。ZAPL和ZAPS的N端包含4个与RNA结合的保守锌指基序,每个基序都有半胱氨酸-组氨酸重复,呈cys-cys-cys-his(CCCH)构型[5]。ZAPL的C端含有多聚ADP核糖聚合酶(PARP)样结构域,该结构域在ZAPS中,因此ZAPL与ZAPS的结构仅在C末端存在差异[6]。ZAP蛋白主要位于细胞质中,可通过核转运信号蛋白1(CRM1)依赖的方式在细胞核和细胞质之间穿梭,进而发挥调节RNA稳定性等功能[6-7]。
2 ZAP蛋白在抗病毒天然免疫反应中的作用
自发现ZAP蛋白有抗M-MLV的增殖后,人们对其抗病毒功能的研究逐渐拓展到其他RNA病毒和一些DNA病毒。截止目前,已经证实ZAP蛋白的抗病毒谱包括猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)和M-MLV在内的RNA病毒及鼠γ疱疹病毒68和乙型肝炎病毒在内的DNA病毒,该蛋白能够结合靶病毒上的ZAP敏感元件(ZAP-responsive element,ZRE),并依赖不同机制抑制靶病毒的复制。研究发现,ZAP蛋白主要通过抑制病毒RNA合成、降解病毒mRNA或抑制病毒mRNA翻译,介导病毒靶蛋白泛素化降解等多种途径,在RNA水平和蛋白水平抑制病毒的复制从而起到抗病毒作用。
2.1 抑制病毒RNA合成
PRRSV是动脉炎病毒科的一种RNA病毒,Nsp9蛋白和N蛋白分别是其非结构蛋白和结构蛋白,Nsp9可与N蛋白相互作用以调节病毒RNA的生物合成。Zhao等[8]报道,Marc-145细胞中ZAP蛋白可以通过其锌指基序与Nsp9的150至160位氨基酸残基相互作用,阻碍病毒RNA的合成,最终抑制PRRSV的复制,并且仅在病毒复制早期阶段起抑制作用。
2.2 降解病毒mRNA
2.2.1 ZAP蛋白与Moloney鼠白血病病毒(M-MLV)
M-MLV是逆转录病毒科的一种RNA病毒。Gao等[2]在筛选M-MLV宿主限制因子时,首次发现了ZAP蛋白的抗病毒功能。研究结果表明,ZAP蛋白能够与病毒的3′长末端重复序列(3′-LTR)相互作用,并通过其锌指基序介导3′-LTR序列的降解,最终抑制Rat2细胞中M-MLV的复制。
2.2.2 ZAP蛋白与辛德毕斯病毒(sindbis virus,SINV)
SINV是披膜病毒科的一种RNA病毒。Kozaki等[9]研究报道,ZAP蛋白通过其锌指基序与病毒mRNA结合,诱导mRNA降解从而起到抗病毒作用。为了深入研究动物水平ZAP蛋白在SINV感染中的作用,Wang等[10]构建ZAP基因敲除小鼠模型,而后制备野生型小鼠和基因敲除小鼠的胚胎成纤维细胞(MEF),感染SINV后,ZAP基因敲除型小鼠比野生型小鼠的MEF细胞中存在更高的病毒载量,另外发现感染病毒后,10日龄的ZAP基因敲除小鼠比野生型小鼠有更高的死亡率。这些结果表明,ZAP蛋白有抗SINV的作用。
2.2.3 ZAP蛋白与埃博拉病毒(ebola virus,EBOV)和马尔堡病毒(marburg virus,MARV)
EBOV和MARV同属丝状病毒科EBOV,L蛋白属于它们的结构蛋白。Muller等[11]研究表明,ZAP蛋白在EBOV和MARV感染宿主Rat2细胞过程中起到重要的抗病毒作用,ZAP蛋白是通过其锌指基序与EBOV和MARV的L基因的mRNA相互作用,最终抑制它们在Rat2细胞中的复制。
2.2.4 ZAP蛋白与鼠异嗜性鼠白血病病毒相关病毒(xenotropic murine leukemia virus-related virus,XMRV)
XMRV是逆转录病毒科的一种RNA病毒,Wang等[12]报道,ZAP蛋白可以抑制XMRV在293T细胞中的复制,其机制是ZAP蛋白能够与病毒mRNA的3'UTR相互作用并介导其降解,最终抑制病毒在293T细胞中的复制。
2.2.5 ZAP蛋白与戊型肝炎病毒(hepatitis E virus,HEV)
HEV是戊型肝炎病毒科的一种RNA病毒。龙飞燕等[13]研究发现,ZAP蛋白在HIV感染A549细胞后显著上调,过表达ZAP能够有效抑制HIV的复制,相反,干扰ZAP后显著促进HIV的复制,且研究表明,ZAP蛋白是通过特异性结合HIV的5′UTR,并介导其降解从而起到抗病毒作用。为了分析在BALB/c小鼠体内ZAP蛋白是否也能够抑制HIV的复制,该团队通过尾静脉注射不同浓度的ZAP质粒后,再注射HIV进行定量分析。结果表明,小鼠体内过表达ZAP可以抑制HIV的复制,且抑制作用呈ZAP质粒浓度依赖性。
2.2.6 ZAP蛋白与柯萨奇病毒B3(coxsackie virus B3,CVB3)
CVB3是小RNA病毒科的一种RNA病毒。Li等[14]研究表明,感染CVB3的HeLa细胞中,ZAP的mRNA和蛋白质水平在24 h增加,并在48 h达到峰值,而后在72 h开始下降,并且HeLa细胞中ZAP蛋白可以通过靶向CVB3的3′和5′UTR的降解来抑制CVB3的复制。为深入研究ZAP蛋白在动物水平是否也具有抗CVB3的作用,该团队进一步构建了BALB/c小鼠ZAP过表达模型,与对照组相比,ZAP过表达小鼠心脏病毒载量更低,心肌炎损伤程度较轻。以上结果说明,ZAP蛋白在小鼠体内可以抑制CVB3的复制。
2.2.7 ZAP蛋白与鼠γ疱疹病毒68(murine gammaherpes virus 68,MHV68)
MHV68是疱疹病毒科的一种DNA病毒,M2蛋白属于其结构蛋白。Xuan等[15]首次将ZAP的抗病毒谱拓展至DNA病毒,研究报道称,ZAP蛋白能够通过与病毒M2基因mRNA的3′非翻译区(3′UTR)相互作用并诱导其降解,从而抑制MHV68在293T细胞中的复制。
2.2.8 ZAP蛋白与乙型肝炎病毒(hepatitis B virus,HBV)
HBV是嗜肝DNA病毒科的一种DNA病毒,Mao等[16]研究表明,HepG2细胞中ZAP蛋白可以通过其锌指基序与HBV RNA末端冗余序列相互作用,并介导其降解从而起到抗病毒作用。Chen等[17]构建ZAP过表达小鼠模型发现,感染病毒后ZAP过表达小鼠肝脏相比于对照组存在更低的病毒mRNA。以上结果说明,ZAP蛋白能够抑制小鼠体内HBV的复制。
2.3 降解病毒mRNA和抑制病毒mRNA的翻译协同作用
2.3.1 ZAP蛋白与人类免疫缺陷病毒(human immunodeficiency virus,HIV)
HIV是逆转录病毒科的一种RNA病毒,截止目前,在ZAP蛋白针对HIV起到重要抗病毒作用及其机制方面的研究较深入。试验证明,ZAP蛋白主要通过诱导病毒mRNA降解或抑制病毒mRNA的翻译两个途径起到抗HIV的作用。
第一,诱导病毒mRNA的降解。Zhu等[18-20]报道,ZAP蛋白是通过与HIV mRNA的5′UTR结合并诱导其降解从而起到抗病毒作用,具体作用机制包括以下两个方面。一方面,ZAP蛋白与病毒mRNA的特异序列直接结合后,募集聚腺苷酸特异性核糖核酸酶(PARN)以去除3′poly(A)尾巴,而后进一步通过招募RNA外切酶加工复合体exosome从3′端介导病毒mRNA的降解;另一方面ZAP蛋白也可依赖其辅助因子p72(RNA解螺旋酶)募集脱帽复合体Dcp1/2以去除病毒mRNA的5'帽子结构,而后进一步通过5′外切酶Xrn1从5′端降解暴露的病毒mRNA。
第二,抑制病毒mRNA的翻译。Zhu等[21]研究表明,ZAP蛋白能够通过干扰翻译起始因子eIF4A和eIF4G之间的相互作用来抑制HIV mRNA的翻译。
2.3.2 ZAP蛋白与日本乙型脑炎病毒(Japanese encephalitis virus,JEV)
JEV是黄病毒科的一种RNA病毒,报道ZAP的过表达显著抑制JEV在A549细胞中的复制,ZAP蛋白能够通过其锌指基序与病毒mRNA的3′UTR序列结合,并介导病毒mRNA的降解或干扰病毒mRNA的翻译,从而抑制JEV在A549细胞中的复制[22]。
2.4 介导病毒靶蛋白泛素化降解和降低PB2 mRNA水平或抑制其翻译协同作用
甲型流感病毒(influenza A virus,IAV)是正黏病毒科的一种RNA病毒,PA、PB2、NA属于其结构蛋白。最初,报道ZAPL能够通过其PARP结构域结合PA和PB2蛋白,介导它们泛素化进而被蛋白酶降解,最终抑制IAV在293T细胞中的复制[23]。而后的研究发现,尽管缺乏PARP结构域,HeLa细胞中ZAPS仍然可以通过降低PB2基因mRNA水平并干扰其翻译,进而抑制IAV的复制,并且这种抑制作用局限于病毒复制的早期阶段[24]。
2.5 ZAP蛋白与其他RNA病毒
塞姆利基森林病毒(semliki forest virus,SFV)、罗斯河病毒(ross river virus,RRV)、委内瑞拉马脑炎病毒(venezuelan equine encephalitis virus,VEEV)同属于披膜病毒科的RNA病毒。Bick等[25]报道,ZAP蛋白在细胞水平能够抑制上述病毒的复制。
禽白血病病毒J亚群(avian leukosis virus subgroup J,ALV-J)是逆转录病毒科的一种RNA病毒。Zhu等[26]研究表明,ZAP的mRNA和蛋白质水平分别在ALV-J感染DF-1细胞后24 h和48 h极度显著升高,然后在72 h开始降低,通过在DF-1细胞中过表达,ZAP能够显著抑制ALV-J的复制。
3 ZAPS通过激活RIG-I介导的I型干扰素反应调节天然免疫应答
病毒入侵时,宿主首先通过天然免疫应答来抵抗病毒感染,由模式识别受体触发,包括RIG-I样受体和Toll样受体等[27-28]。在RIG-I样受体信号通路中,RIG-I识别病毒RNA结合并激活线粒体抗病毒信号蛋白(MAVS),该蛋白通过激活胞质IκB激酶(IKK)和TANK结合激酶1(TBK1)进而分别刺激核转录因子(NF-κB)和干扰素调节因子3(IRF3)。NF-κB和IRF3协同诱导干扰素β(IFN-β)和其他抗病毒分子的产生[29-32]。
Hayakawa等[33]研究发现,ZAPS能够通过其锌指结构与RIG-I相互作用,进而激活NF-κB与IRF3信号通路,最终增强RIG-I介导的I型干扰素的产生。试验结果表明,293T细胞中ZAPS能够通过增强IFN-β的表达最终抑制流感病毒H3N2的复制,另外,ZAPS能够通过增强A549细胞中IFN-β的表达最终起到抗新城疫病毒的作用。
4 病毒感染诱导ZAP基因表达调控的机制研究
ZAP蛋白作为一种新型宿主限制因子,病毒感染诱导其高表达,发挥着重要的抗病毒作用。Wang等[34]报道,ZAP基因的转录可直接由IRF3控制,也可以是病毒感染过程中干扰素自分泌或旁分泌信号转导的结果,但不依赖于NF-κB活性。研究表明,在感染的早期,IRF3被激活并转移到细胞核中,而后与ZAP基因启动子结合,从而上调ZAP基因的转录水平,此过程独立于干扰素信号传导。随后,受感染的细胞分泌IFN-β,继而激活感染细胞和或未感染邻近细胞中的JAK-STAT(janus family of tyrosine kinase-signal transducer and activator of transcription)信号传导,导致STATs的活化,后者转移到细胞核中与ZAP基因启动子结合并维持ZAP基因的表达。
5 ZAP蛋白的表达调控及其抗病毒等机制总结
ZAP蛋白的合成需要经过转录、翻译,其中转录水平受到IFNβ、IRF3的调控。一方面,ZAP蛋白可作为宿主限制因子,主要通过抑制病毒RNA合成、降解病毒mRNA或抑制病毒mRNA翻译和介导病毒靶蛋白泛素化降解等多种途径在RNA水平和蛋白水平抑制病毒的复制,但其对不同病毒的抗病毒机制不尽相同,具体机制见表1。
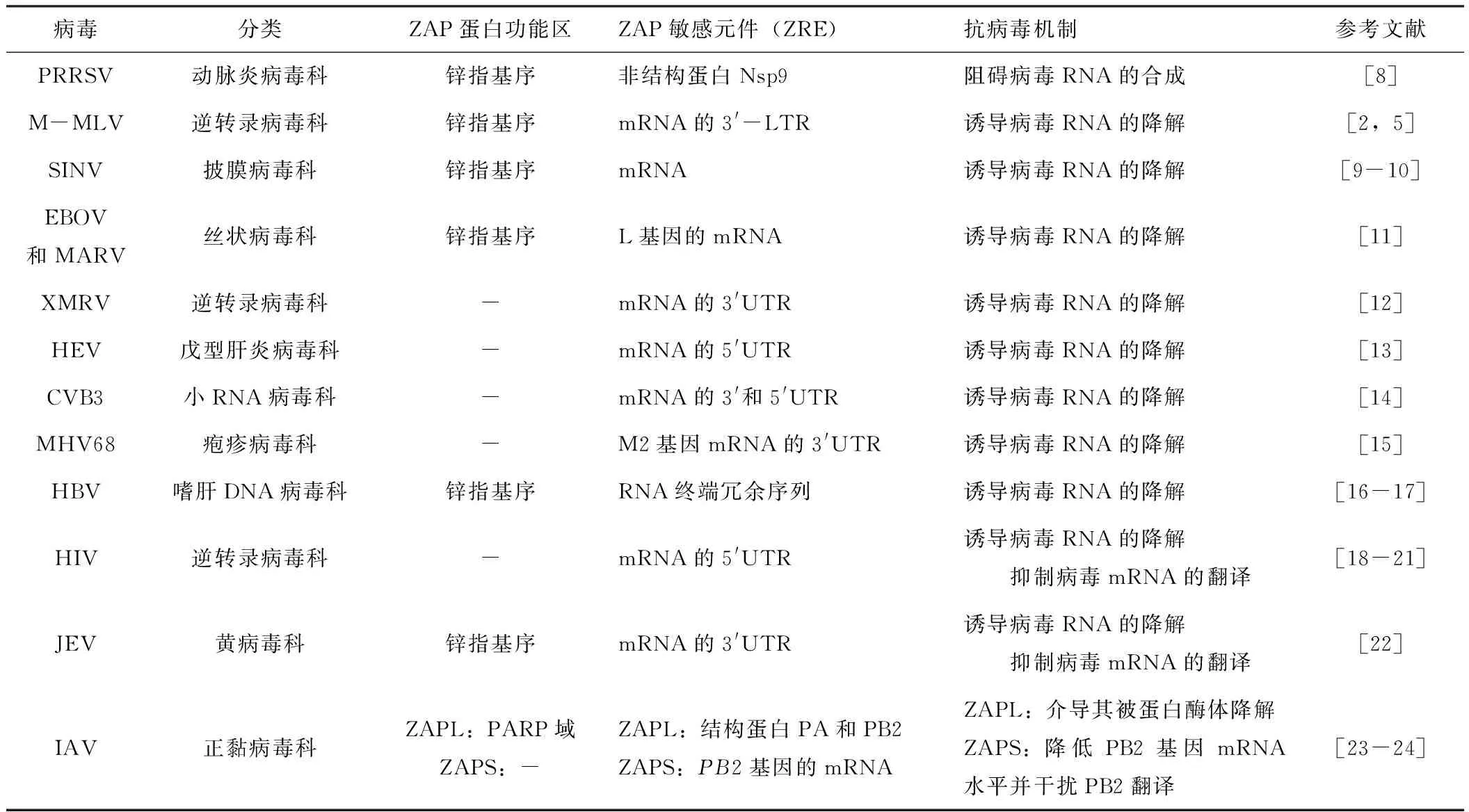
表1 ZAP蛋白在抗病毒天然免疫中的作用
另一方面,ZAP蛋白可作为免疫调节因子,通过增强RIG-I介导的I型干扰素反应诱导抗病毒因子的产生,从而发挥抗病毒作用,具体机制见图1。
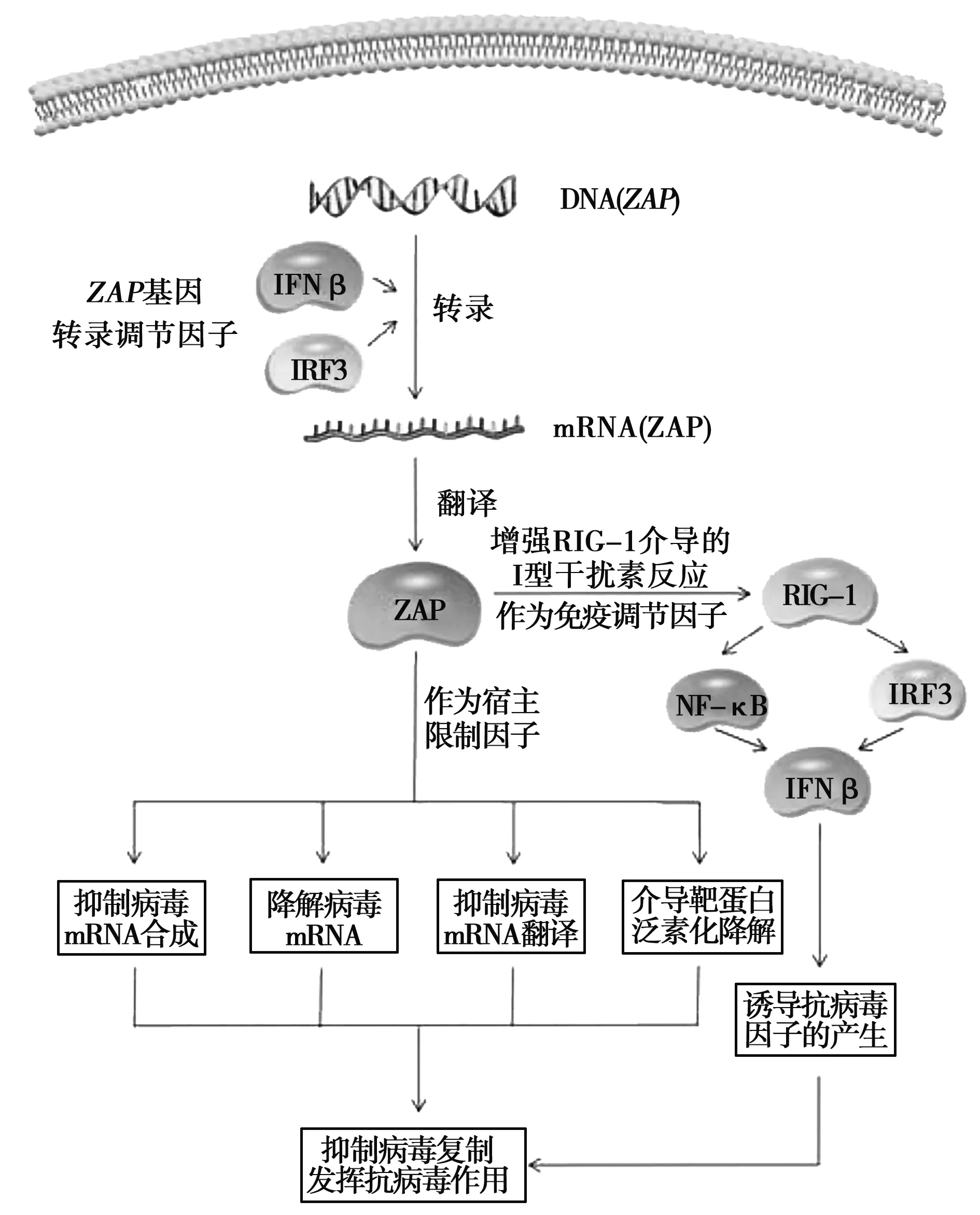
图1 ZAP蛋白的表达调控及其抗病毒等机制
6 不同病毒拮抗ZAP蛋白介导的宿主天然免疫应答
天然免疫系统的细胞(如巨噬细胞)可表达模式识别受体(pattern recognition receptor,PRR),以识别病原体相关的分子模式(pathogen-associated molecular patterns,PAMP),启动细胞内信号转导通路,诱导分泌细胞因子或趋化因子,发挥抗感染作用。病毒感染能够激活天然免疫应答并诱导包括ZAP在内抗病毒因子的表达,其可在病毒感染的不同阶段抑制病毒的复制[35]。同时,在与宿主免疫系统相互作用的过程中,不同病毒也已进化出一系列拮抗ZAP蛋白介导的天然免疫应答抗病毒途径,来满足自身复制的需要,主要包括抑制ZAP基因mRNA合成、降解ZAP基因mRNA、降解ZAP蛋白和抑制ZAP蛋白与靶病毒的mRNA或目标蛋白结合等多种机制。
6.1 抑制ZAP基因mRNA合成
MicroRNA(miRNA)是一类长度约为22 个核苷酸的非编码单链RNA分子,它能够通过与目标mRNA特异结合,从而抑制转录后基因的表达。Kincaid等[36]报道,托克特诺病毒(torque teno virus,TTV)编码的TTV-tth8miRN能够通过靶向N-Myc相互作用因子(NMI)抑制干扰素信号通路,从而促进病毒逃避宿主免疫应答。
牛病毒性腹泻病毒(bovine viral diarrhea virus,BVDV)是黄病毒科的一种RNA病毒。刘升[37]报道,BVDV感染宿主细胞MDBK后18至48 h内,ZAP基因mRNA水平显著下降,而过表达ZAP的MDBK细胞显著抑制病毒的复制,表明BVDV能够通过某种机制抑制ZAP的转录,从而拮抗ZAP蛋白的抗病毒活性。该研究同时发现,BVDV感染后能够提高miR-2459的表达,miR-2459可直接靶向ZAP基因3′UTR区的miRNA识别位点并导致其转录水平的下降,从而促进BVDV在细胞中的复制。
6.2 降解ZAP基因mRNA
I型单纯疱疹病毒(herpes simplex virus 1,HSV-1)是疱疹病毒科的一种DNA病毒。UL41属于其非结构蛋白,是一种具有RNase活性的核酸内切酶。Murphy等[38]报道,UL41可以通过多种机制抑制IFN介导的抗病毒反应。Su等[39]研究表明,ZAP蛋白并不能抑制HSV-1在293T细胞中的复制,但过表达ZAP能够显著抑制UL41突变失活的HSV-1在宿主细胞中的复制。该研究发现,HSV-1是通过UL41的RNase促进ZAP基因mRNA的降解,从而拮抗ZAP蛋白的抗病毒活性。
6.3 降解ZAP蛋白
6.3.1 肠道病毒A71型(enterovirus A71,EV-A71)与ZAP蛋白
EV-A71是微RNA病毒科的一种RNA病毒,结构蛋白VP1和非结构蛋白3C蛋白酶(3Cpro)在病毒与宿主互作过程中起到重要作用。多项研究表明,EV-A71已进化出多种逃逸宿主细胞I型干扰素反应的策略,从而起到拮抗宿主天然免疫的作用[40-42]。Xie等[43]报道,在EV-A71感染过程中,HeLa细胞中ZAPL基因的mRNA表达显著上调,而ZAP蛋白水平却显著下降,尤其在感染后30 h最为明显。此外,过表达ZAPL可降低VP1及3C蛋白的表达即抑制病毒的复制,且EV-A71的3C蛋白能够通过其蛋白酶活性介导ZAP蛋白的切割,从而拮抗宿主天然免疫。
6.3.2 PRRSV与ZAP蛋白
nsp4、nsp7β、nsp9、nsp12均属于PRRSV的非结构蛋白,其中nsp4属于典型的具有切割活性的3C样丝氨酸蛋白酶,Chen等[44]报道,nsp4能够通过抑制IFN-β的转录从而起到拮抗宿主天然免疫的作用。杨玉婷[45]研究表明,ZAPL与ZAPS的过表达均能显著抑制MARC-145细胞中PRRSV的复制,且nsp4能够呈剂量依赖性识别ZAP第411位谷氨酸并切割ZAP蛋白,从而拮抗ZAP蛋白介导的抗病毒作用,此过程不依赖细胞凋亡和细胞自噬途径。
6.4 抑制ZAP蛋白与靶病毒的mRNA或目标蛋白结合
NS1IAV的非结构蛋白之一。许多研究表明,NS1蛋白在流感病毒拮抗宿主天然免疫作用中扮演着重要角色,例如它可以通过抑制三联基序蛋白25(TRIM25)的E3连接酶活性,限制流感病毒感染诱导的I型干扰素的产生[46]。ZAPL与ZAPS在IAV感染过程中均起到显著的抗病毒作用,Liu等[23]报道,IAV的PB1蛋白能够促使ZAPL、PA和PB2的解离,保护PA和PB2蛋白免受蛋白酶体的降解,从而起到拮抗ZAPL抗病毒活性的作用。Tang等[24]研究发现,IAV的NS1蛋白能够抑制ZAPS与病毒mRNA的结合,拮抗ZAPS的抗病毒作用。
7 总结与展望
作为新型抗病毒因子,ZAP蛋白可通过抑制RNA合成、降解mRNA或抑制mRNA翻译、介导靶蛋白泛素化降解等多种途径直接或间接抑制RNA或DNA病毒的复制,从而在宿主抗病毒天然免疫应答中发挥重要作用,此外,它还能通过激活RIG-I介导的I型干扰素反应调节天然免疫。同时,在与宿主免疫系统斗争的过程中,病毒也已进化出多种拮抗ZAP蛋白介导的抗病毒天然免疫应答机制,包括抑制ZAP mRNA合成,降解ZAP mRNA,降解ZAP蛋白和抑制ZAP蛋白,与靶病毒的mRNA或目标蛋白结合等。
ZAP蛋白能够通过其不同抗病毒功能区结合靶病毒上的ZAP敏感元件,并依赖不同机制抑制靶病毒的复制,但ZAP并非对所有病毒都具有抗病毒作用。Bick等[25]研究表明,水泡性口炎病毒、脊髓灰质炎病毒、黄热病病毒在ZAP过表达的细胞中仍可正常复制,这些现象背后的分子机制是什么,ZAP蛋白在发挥抗病毒作用过程中,又是哪些因素决定着其抗病毒功能区和靶病毒上ZAP敏感元件的选择,这些问题还有待进一步深入研究。
猜你喜欢
肝博士(2022年3期)2022-06-30 02:48:52
肝博士(2021年1期)2021-03-29 02:32:14
科学(2020年3期)2020-11-26 08:18:22
学苑创造·A版(2020年9期)2020-10-13 09:41:02
肝博士(2020年4期)2020-09-24 09:21:26
当代水产(2020年3期)2020-06-15 12:03:02
疯狂英语·初中天地(2020年3期)2020-05-21 03:36:52
小学生学习指导(低年级)(2017年10期)2017-10-10 01:00:05
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:55
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
