
短翅足距小蠹生物学特性初步研究
2020-12-08 08:03:34邱建生吴跃开李小荣孙坜坜陈又清
林业科学研究 2020年5期
翟 鸣,邱建生,吴跃开,钱 涛,李小荣,孙坜坜,陈又清
(1. 安顺市森林植物检疫站,贵州 安顺 561000;2. 贵州省林业科学研究院,贵州 贵阳 550005;3. 中国林科院资源昆虫研究所,云南 昆明 650000)
短翅足距小蠹(Xylosandrus brevis(Eichhoff))属小蠹科(Scolytidae)齿小蠹亚科(Ipinae)材小蠹族(Xyleborini)足距小蠹属(Xylosandrus)[1],原属材小蠹属(Xyleborus),异名为(Xyleborus brevis( Eichhoff) )[2], (Xyleborus cucullatusBlandford)[3],Buchanan 将其转移到足距小蠹属[4],这一分类结果已得到国内外学术界普遍认同[5-7]。主要分布于日本、朝鲜、泰国,以及中国的台湾[8]、西藏[1-2]、四川[9]。寄主植物有小檗(Berberis sp.)、山茶花(Camellia japonicaLinn.)、茶梅(Camellia sasanquaThunb.)、天竺桂(Cinnamomum japonicumSieb.)、柿树(Diospyros kakiThunb.)、钝 齿 水 青 冈 (Fagus crenataBlume) 、 银 桦(Grevillea robustaA. Cunn.) 、 金 缕 梅(Hamamelis mollisOliver)、绣球花(Hydrangeamacrophylla(Thunb.) Seringe)、山胡椒(Lindera glauca(Sieb. et Zucc.) Bl)、日本润楠(Machilus japonicaSieb. et Zucc.) 、 红 楠 (Machilus thunbergiiSieb. et Zucc.) 、 软 弱 杜 茎 山 (Maesa teneraMez) 、 泡 花 树 (Meliosma cuneifoliaFranch) 、 油 乌 药 (Parabenzoin praecoxSieb. et Zucc.)、栎(Quercussp.)、菝葜(Smilax chinaL.)、玉铃花(Styrax obassiaSieb. et Zucc.)、荚蒾(Viburnumsp.)、锦带花(Weigela hortensis(Bunge) A.DC.)[2-3,9]。短翅足距小蠹属于贵州新记录种,2017 年, 首次在安顺地区发现其危害广玉兰(Magnolia grandifloraLinn.),造成大量枝条枯死,严重破坏城市园林景观。国内对其系统研究甚少,仅任茂琼等对成虫形态特征进行了较为深入的报道[9]。在全面调查广玉兰受害情况的基础上,对短翅足距小蠹的生物学特性进行了室内外观察研究,现将研究结果报告如下,以期为害虫治理工作提供参考。
1 材料与方法
1.1 室内饲养及观察
1.1.1 生活史观察 2018 年3 月,将野外采集的广玉兰受害枝条带回室内,选取从蛀孔外能观察到成虫的枝条,剪掉叶片,截成小段,每段枝条上保留1 个 蛀 孔 。 准 备10 个1 000 mL 广 口 瓶 , 按1~10 进行编号。1 个广口瓶内,放入1 段枝条,用纱布罩住瓶口,然后用橡皮筋扎紧,置于室内通风处,保证室内外温湿度大体一致。观察越冬成虫出孔情况,记录每天成虫出孔数量和雌雄比例。根据当天出孔的雌成虫数量,采集适量新鲜广玉兰枝条,剪掉叶片,截成20 cm 左右小段放入瓶内,观察越冬成虫钻蛀情况。枝条脱水或发霉变色,及时更换新鲜枝条。
待新鲜枝条被钻蛀后,将枝条分成两部分处理。一部分直接移入新的广口瓶,用纱布橡皮筋封口,发现有新成虫羽化,再补充新鲜广玉兰枝条继续饲养,观察小蠹世代数;一部分先纵向解剖,留下有虫的一半坑道,再用透明保鲜薄膜缠好后,移入新的广口瓶,用纱布橡皮筋封口,观察小蠹生长发育情况。每3 天向瓶内喷适量清水,保持枝条新鲜。
1.1.2 危害特点观察 采集受害枝条,用游标卡尺测量枝条直径、蛀孔直径。纵向剖开枝条,测量坑道剖面长度和宽度,观察记录坑道内小蠹虫态、数量和形态特征。随机测量数头各虫态相关数据,体长和体宽平均值用x±SE 表示。
1.1.3 寄主范围测试 取新鲜红叶石楠(Photinia×fraseriDress)、 桂 花 (Osmanthussp.)、 香 樟(Cinnamomum camphora(L.) Presl.) 枝 条 各2 段 和10 头越冬雌成虫一起放入广口瓶,用纱布橡皮筋封口,测试寄主范围。
1.2 危害情况调查
对安顺城区广玉兰受害情况开展调查,以枝梢受害率确定单株发生情况,以受害株率确定总体发生情况,参照国家林业局林业有害生物发生及成灾标准,林业主要有害生物调查总则[10-11],按枝梢受害率(或受害株率)确定危害程度,10%~20% 为轻度,21%~50% 为中度,50% 以上为重度。同时,设置标准株,定株观察广玉兰受害枝条变化状况。同时对周边多种植物进行调查,观察是否受短翅足距小蠹危害。
1.3 生殖类型研究
观察统计越冬期坑道内小蠹雌雄成虫性比结构,分析其可能存在的生殖类型。
2 结果与分析
2.1 形态特征
雌成虫 圆柱形,体长2.38~2.81 mm,平均值2.55±0.12 mm;体宽0.96~1.21 mm,平均值1.04±0.07 mm(n=21)。触角着生于复眼前缘下方,柄节棒状,鞭节5 节。前胸背板黄褐色,背面观,呈盾形,背中部基缘的前面密生细柔竖立的黄色茸毛;侧面观前2/3 为瘤区,呈弓曲上升,后1/3 为刻点区,呈倾斜下降直达鞘翅基部;瘤区颗瘤呈同心圆排列,颗瘤中后部密生直立或半倒伏茸毛;刻点区刻点细小。鞘翅黄褐色,后半部在后翅映衬下呈黑褐色;背面观前2/3 平直,后1/3 急剧下降形成一截面[9],截面茸毛明显;侧面观平直部分短于截面部分,截面中部略突出,呈浅圆弧形(图1)。
雄成虫 圆柱形,体长1.75~2.05 mm,平均值1.97±0.09 mm;体宽0.71~1.00 mm,平均值0.89±0.08 mm(n=18)。前胸背板黄褐色,背面观,呈盾形,前缘向前延伸,呈中部凹陷,两侧上卷的“M” 形铲状结构。前翅黄褐色,无后翅,不能飞行。鞘翅背面观前 2/3 平直,后 1/3 急剧下降形成一截面[9],截面茸毛明显;侧面观平直部分短于截面部分,截面中部略突出,呈浅圆弧形。成虫存在明显二型性,雌雄个体差异明显(图1)。
卵 椭圆形,长0.72~0.84 mm,平均值0.77±0.05 mm;宽0.36~0.40 mm,平均值0.38±0.02 mm(n=19)。白色半透明,表面光滑(图2a)。
幼虫 全体乳白色。老熟幼虫长与成虫接近,常呈C 形弯曲,无足。上颚较发达,浅褐色(图2b)。
蛹 离蛹,初期乳白色,后渐变浅褐色,与成虫近长(图2c)。
2.2 生物学特性

图1 短翅足距小蠹成虫背、侧面观(左雄右雌)Fig. 1 Back and side view of the adult of Xylosandrus brevis(left male and right female)

图2 短翅足距小蠹卵、幼虫和蛹Fig. 2 Egg、Larva and Pupa of Xylosandrus brevis
2.2.1 生活史 短翅足距小蠹在安顺地区1 a 发生1~2 代,有世代重叠现象,以成虫在受害枝条坑道内越冬。翌年5 月,越冬成虫开始外出活动,5 月上旬产卵,卵期5~6 d。5 月中旬幼虫出现,幼虫期约15 d。5 月下旬化蛹,蛹期约5 d。6 月上旬成虫羽化。早期羽化的第一代成虫出孔后,继续侵入健康枝条,繁殖第二代,并与第二代成虫一起在坑道内越冬;晚期羽化的成虫不再出孔活动和繁殖,与其母代雌成虫一起在坑道内越冬。雌成虫寿命可达1 a 以上。由于越冬成虫出孔活动持续时间长,因此,第一代成虫常与越冬代成虫同时出现(表1)。
2.2.2 生活习性 成虫 5 月上旬,越冬成虫中的雌成虫率先从侵入孔爬出,并很快死亡,之后,子代成虫陆续出孔,整个出孔期,历时3 个多月,一直持续到8 月下旬,出孔盛期集中在5 月中旬至6 月上旬(图3)。成虫出孔时间,多在每日下午1 点至5 点之间,此时段出孔的成虫活性最强。其他时段出孔的成虫活性较弱,甚至静止不动。子代雄成虫出孔后四处爬行,数天后死亡。子代雌成虫出孔后数小时内即可蛀孔危害,常侵入附近直径6~11 mm 左右的健康枝条,侵入孔圆形,直径约0.8~1.5 mm(图4c、图4D、表2)。雌成虫向内钻蛀到达髓心后,便向枝条一端或两端修筑坑道,一边筑坑道,一边产卵,并不时将蛀屑和粪便推出坑道外,最后形成长9~57 mm,宽1.5~3 mm 的不规则坑道(图4e、表2)。雌成虫多数时间用身体堵住孔口,保护坑道和子代,表现出明显的护巢习性。在雌成虫修筑坑道的过程中,其身上携带的真菌孢子在坑道内壁萌发生长,菌丝和孢子为小蠹生长提供食物。

表1 安顺城区短翅足距小蠹生活史(2017—2018) Table 1 Life history of Xylosandrus brevis in Anshun(2017—2018)

图3 短翅足距小蠹越冬成虫出孔活动时期Fig. 3 Overwintering adult activity period of Xylosandrus brevis
每个坑道内小蠹数量1~25 头不等,雌雄性比约为10:1。子代雌雄成虫在坑道内完成交尾,由于雌雄性比差异大,导致少数雌成虫没有交尾机会。对22 个越冬期蛀道内小蠹雌雄成虫性比结构观察发现,有8 个蛀道内全部为雌成虫,占比达27%,在这类全雌成虫的蛀道内,雌成虫数量最多可达6 头,最少仅有1 头。这种较高比例的全雌成现象说明,短翅足距小蠹可能存在两性生殖和孤雌生殖两种生殖方式,以两性生殖为主。进行两性生殖的小蠹,其后代数量较多,后代中既有雌虫,也有雄虫;未交尾的雌成虫繁殖的后代数量较少,且全部为雌虫,有的甚至不繁殖后代(表2)。
卵 卵单粒或数粒粘附在坑道内壁,5~6 d 孵化。
幼虫 幼虫取食坑道内壁真菌,约15 d 后化蛹,老熟幼虫不做蛹室。
蛹 蛹腹部能活动,历期约5 d。
2.2.3 危害特点及分布 短翅足距小蠹能造成枝条大量枯死,枯死枝条上的叶片外形基本保持不变,仅自叶柄处下垂,并不从枝条上脱落。受外力作用,枯死枝条常从蛀孔处折断,野外调查极易发现。 定株观察表明, 短翅足距小蠹主要危害2~3 年生枝条,并不危害当年生枝条或更老的枝条。健康广玉兰一旦有枝条被害,其周围枝条会次第表现出受害症状,枯死枝条呈现较为均匀的簇状分布(附图4a、图4B)。
调查表明,短翅足距小蠹在安顺市所属的西秀区、平坝区、普定县、开发区4 个县区均有分布,占县区总数的50%。持续观察结果显示,广玉兰上短翅足距小蠹扩散速度快,危害大,2017 年调查平均受害株率为16.2%, 尚属轻度危害水平,2018 年调查平均受害株率已上升至39.3%,达中度危害水平(表3)。

图4 短翅足距小蠹危害状Fig. 4 Damage situation of Xylosandrus brevis
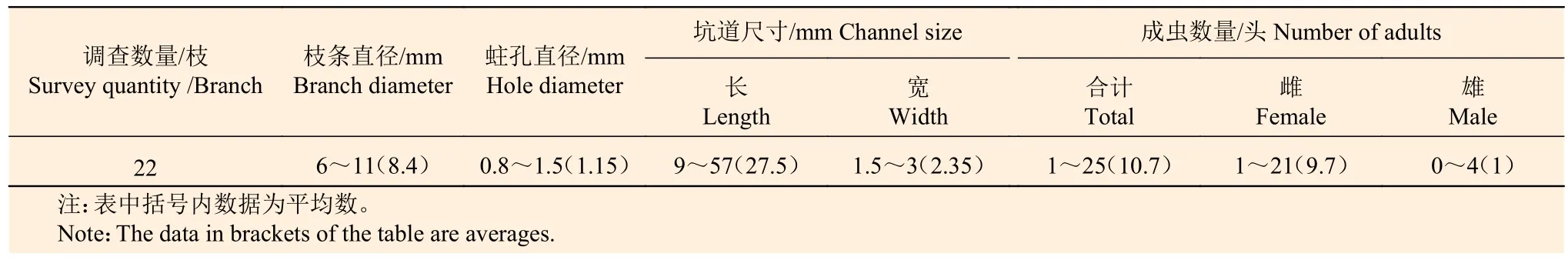
表2 广玉兰受害枝条调查(2018) Table 2 Investigation for damaged branches of Magnolia grandiflora(2018)
导致枝条枯死的原因主要是:其一,短翅足距小蠹蛀空枝条的髓心造成水分传输中断;其二,雌成虫修筑坑道过程中,其贮菌器内的真菌侵入枝条,菌丝在坑道内壁生长,导致木质部由内向外颜色加深,质地疏松。通过对广玉兰受害枝条变色组织初步分离培养,分别检测出拟茎点霉(Phomopsissp.)及镰刀菌(Fusariumsp.)等重要的植物病原真菌[12]。在小蠹和真菌的合力作用下,木质部水分传导系统遭受破坏,枝叶逐渐失绿变黄直至变成褐色,脱水萎蔫,最后枯萎死亡。
短翅足距小蠹的少数雌成虫有转移危害特点,即在一个健康枝条上钻蛀形成1.5~5 mm 浅坑后,转而危害其它健康枝条,并在其中繁育下一代。留下浅坑的健康枝条因受雌成虫携带真菌感染,蛀孔周围表皮颜色变深,呈近圆形黑褐色膏药状,用手挤压枝条能溢出水状物,枝条亦会缓慢枯死。
调查发现,短翅足距小蠹在安顺城区除危害广玉兰外,也危害二球悬铃木(Platanus acerifoliaWilld.)(图4F),常与同属的另一种小蠹——小滑足距小蠹(Xylosandrus compactus(Eichhoff))寄生于同一寄主上。

表3 安顺城区广玉兰受害情况调查(2017—2018) Table 3 Investigation on the victimization of Magnolia grandiflora in Anshun(2017—2018)
通过调查及寄主测试,未发现短翅足距小蠹危害红叶石楠、桂花、香樟等植物,是否还有更多的寄主,有待进一步研究。从区系分布看,短翅足距小蠹为东洋及古北两界的跨界分布种,根据已有记载的分布区域,我国大部分地区均是其潜在的发生区域,应引起高度重视[12]。
2.3 防治建议
2.3.1 监测 加强苗圃和种植区域监测,发现有枝条枯死,即刻采集镜检,检查是否有侵入孔,并通过侵入孔观察孔口的成虫,进一步解剖是否有坑道及短翅足距小蠹其他虫态,一旦发现,应及时开展防治工作。
2.3.2 人工防治 根据短翅足距小蠹生物学特性,2018 年冬季,园林部门对市区部分受害广玉兰枯死枝条进行清理,目前清理后的广玉兰未出现新受害枝条,防治效果比较明显。因此,在越冬成虫出孔之前,修剪枯死枝条,并集中烧毁,能有效降低虫口基数,提高防治效果。
2.3.3 化学防治 国内有使用杀螟硫磷和毒死蜱+氯氰菊酯防治同属其它小蠹的报道[13-14],均取得较好的防治效果,值得参考借鉴。
2.3.4 诱捕防治 可利用配有无水乙醇作为诱芯的粘虫板诱捕器进行诱杀,有一定的诱杀效果[15]。
3 讨论
足距小蠹属(Xylosandrus)很多的亚洲种类成为欧洲、 美洲的入侵物种, 如削尾足距小蠹(Xylosandrus mutilatusBlandford),小滑足距小蠹(Xylosandrus compactusEichhoff.)等[16],我国也把该属的非中国种列为禁止入境的种类。研究发现短翅足距小蠹(Xylosandrus brevis(Eichhoff))和小滑足距小蠹(Xylosandrus compactusEichhoff.)均对广玉兰(Magnolia grandifloraLinn.)和悬铃木(Platanus acerifoliaWilld.)两种植物表现出明显的偏好性,而这两种植物的原产地均为美洲,是我国引入的外来物种,因此,值得进一步开展足距小蠹对两种外来物种的选择机制研究,对深入研究足距小蠹属(Xylosandrus)的亚洲种类为什么成为欧洲、美洲的入侵种有积极的意义。
尽管孤雌生殖在鞘翅目昆虫中非常罕见,但在足距小蠹属中,孤雌生殖在有的种类中是存在的,如光滑足距小蠹(Xylosandrus germanus)能行产雄孤雌生殖[17-18]。该小蠹的雌雄性比约为9:1 或10:1[18]。与短翅足距小蠹很相似。对于短翅足距小蠹可能存在孤雌生殖的现象,是基于对22 个越冬期蛀道内小蠹雌雄成虫性比结构观察发现,有8 个蛀道内全部为雌成虫,而且占比达27%,不是偶然现象,因此,推测其存在孤雌生殖的可能性。是否如此,还需作进一步的研究。
用于室内饲养的枝条容易干枯脱水,难以保证与自然生长的枝条性状相一致,导致短翅足距小蠹生长发育环境存在差异,因此,本研究的结果可能与自然状态下短翅足距小蠹生物学特性尚有一定差距,还需做更深入的研究和比较。
4 结论
短翅足距小蠹在安顺城区1 a 发生1~2 代,有世代重叠现象。子代成虫在坑道内交尾,并在坑道内越冬。由于雌雄性比差异大,导致少数雌成虫可能没有交尾机会。成虫可能存在两性生殖和孤雌生殖两种生殖方式,以两性生殖为主。两性生殖繁殖的后代数量较多,既有雌虫,也有雄虫;孤雌生殖繁殖的后代数量较少,且全部为雌虫,有的甚至不繁殖后代。5~8 月份,为成虫出孔活动期,高峰期集中在5 月中旬至6 月上旬。雄成虫无后翅,不能飞行,出孔数天后死亡。雌成虫翅完整,不善飞行。母代雌成虫出孔后很快死亡;子代雌成虫出孔后,常选择2~3 a 生健康广玉兰枝条危害,通过小蠹危害和真菌寄生,破坏枝条髓心,阻断水份输送,造成枝叶枯死,被害枝条常呈现出较为均匀的簇状分布。
研究表明,短翅足距小蠹扩散速度快,对园林景观影响较大,应引起高度重视。目前,较为有效的防治措施是在冬春季节集中清除受害枝条,减少越冬虫口基数,控制其扩散蔓延,降低危害。
猜你喜欢
作文与考试·小学高年级版(2023年8期)2023-07-17 09:32:52
青海农林科技(2022年3期)2022-09-26 03:16:18
林业科学(2021年12期)2021-02-12 06:46:22
浙江林业科技(2019年5期)2019-01-10 16:43:51
环球时报(2019-01-10)2019-01-10 06:25:43
中国诗歌(2018年5期)2018-11-14 20:52:30
作文评点报·低幼版(2018年30期)2018-09-27 12:21:24
作文评点报·低幼版(2018年30期)2018-09-27 12:21:24
防护林科技(2013年1期)2013-08-08 11:20:18
东北林业大学学报(2011年8期)2011-03-31 16:51:12
