
昆虫的趋光性及其应用于害虫治理的研究进展
2020-12-07 02:52张锦芳徐文平陶黎明编译
世界农药 2020年11期
张锦芳,张 阳,徐文平,陶黎明 编译
(华东理工大学 药学院 药物化工研究所,上海市化学生物学重点实验室,上海 200237)
由于全球环境和气候的变化,导致虫害频发,严重影响了农业生产。喷洒化学农药是目前控制害虫的主要手段之一,但长期过度使用农药造成了许多问题,诸如对使用者的伤害、害虫抗药性、环境污染和生物多样性丧失。害虫防治已成为有机农业面临的最大挑战之一,因此,寻找能有效控制害虫的新颖的、可替代的、生态友好的防治手段(如电网捕虫器、诱捕器和生物农药),在世界范围内受到了广泛的关注。
许多昆虫特别是夜行性昆虫,在夜间会向人造光移动,这种现象称为正趋光,而昆虫远离光源的现象就称为负趋光。昆虫趋光行为的程度取决于光源的属性(例如,波长和强度)和其他条件(如天气和昆虫的生理状况)。根据视觉线索和时间线索,昆虫可以精确地躲避捕食者,并找到宿主、猎物和配偶。不同的昆虫对光的敏感性不同,例如夜蛾的复眼对光的敏感性比蝴蝶的复眼高大约1 000倍。大多数昆虫能够感知波长在350~760 nm的光,其中也包括像紫外线(UV)和红外线(IR)这些人类肉眼无法看到的光。昆虫的复眼通常含有3种感光受体:紫外光敏感型、蓝光敏感型和绿光敏感型;因此,许多昆虫对这3个波段的光线表现出强烈的趋光行为。除了这3种感光受体外,一些膜翅目的叶蜂科昆虫还有一种红色感光受体。
为了寻找引起昆虫强烈的趋光行为的特定光波长,许多研究人员将昆虫的电生理研究与昆虫趋光行为的研究相结合。在光的刺激下,昆虫复眼视网膜细胞发出一种电信号传递到大脑,从而导致昆虫产生相应的行为反应。当昆虫暴露在光照下,复眼视网膜细胞的视网膜电图(ERG)数值可以用来表征昆虫对光的敏感性。然而,一些关于昆虫趋光性的研究报告显示昆虫ERG结果与趋光行为试验的结果并不相同。因此,ERG水平仅能代表昆虫眼睛的光敏性,而向光源或远离光源的行为可能很难用ERG数据来判断。
利用昆虫光趋向的特性,可以吸引或驱退农业、温室、森林和粮仓中的害虫,然而,光照可能会同时吸引益虫和害虫,因此使用诱虫灯进行害虫治理时,必须根据昆虫的趋光特性对诱虫灯的光源做出相应的调整。
已研究了许多昆虫对特定光源的趋光行为反应,其中一些研究结果已经应用于防治实践。例如,温室中已使用诱虫灯捕捉蚜虫和粉虱;通过趋光反应研发的捕蚊器,已被用于减少蚊媒介传播疾病的发生。从保护生物多样性和低能耗的方面考虑,目前常用发光二极管作为诱虫灯的光源。诱虫灯的使用在害虫综合管理(IPM)中具有重要意义。本文介绍了一些昆虫在特定波长下正向趋光行为方面的研究进展,总结了70多种昆虫的正趋光行为的敏感波长信息,旨在为研究昆虫行为与特定光之间的关系提供更多有价值的信息,用以阐明昆虫趋光行为的原因,以便开发更高效、环保和经济的诱虫灯。
1 昆虫趋光行为假说
昆虫对光的敏感性和行为反应,因其不同的生活史和生物学特性而不同。昆虫求偶、种群分布、觅食、防御捕食以及对于食物、宿主、休息地点、产卵地点和越冬地点的位置判断都受视觉的控制。昆虫的趋光行为与其适应光照、昼夜周期、光周期和光毒性有关。为什么许多昆虫会表现出特殊的趋光行为,目前尚不清楚其原因。几十年来,研究人员试图通过行为、生理、生化和分子试验来解释昆虫的趋光行为,并根据试验结果提出了一些假说。
光定位假说认为夜行昆虫将星光和月光作为飞行或运动的坐标,昆虫的眼睛总是与光源保持一定的角度,以保持夜间的直线飞行,夜间昆虫会将人造光与星光或月光混淆。当昆虫离光源太近时,昆虫便不能保持直线飞行,因此就会冲向光源或在周围螺旋飞行。虽然这一假设解释了昆虫正向趋光性行为,但它并不能解释一些昆虫的静止或反向趋光行为。
生物天线假说认为昆虫触角以类似于无线电天线的方式感知异性释放的信息素分子的振动。信息素分子的振波属于远红外光谱,昆虫在寻找配偶时使用触角感受到这些振波,因此,一些昆虫可能会被发射远红外波长的光源所吸引。然而这一假设不能解释昆虫在其他波长下(如紫外光和可见光)的趋光行为。
光干扰假说认为昆虫在夜间已适应暗光状态,当接近光源周围明亮的区域时,因无法迅速适应明亮的环境而冲向光源。一些研究表明,这种趋光行为的发生是因为已适应暗光的昆虫复眼在强光的刺激下无法正常调节,从而其正常行为受到干扰。另一项研究表明,这种现象的发生是因为昆虫试图避开它们不适应的环境(或明亮的光线),但因强光刺激而迷失方向。虽然这一假设可能解释了停留在光源周围昆虫的趋光性,但它并不能解释为什么远处的昆虫也会向光源移动。
光胁迫假说从光应激反应的角度对昆虫趋光行为进行了解释。夜间昆虫已经习惯了长时间处于黑暗(或弱光)环境中,它们对于强光的应激能力会减弱。对特定波长的反趋向行为可能表明昆虫试图减少光照带来的刺激。当昆虫暴露在紫外光下时,体内几种抗氧化酶的活性显著增加,乙酰胆碱酯酶水平降低,乙酰胆碱滴度增加,昆虫持续兴奋,可能会导致昆虫混淆飞行方向(例如向光冲),甚至导致死亡(路灯周围的昆虫尸体)。但这一假设也并不能解释昆虫会从远处飞向光源的原因。
虽然许多研究已经从不同的角度对昆虫趋光行为进行了解释,但更加完整解释仍然需要更多的科学研究。
2 影响昆虫趋光行为的3个因素
昆虫对人造光的敏感性受外部环境(例如,光强、光照时间、偏振性、天气和季节)和昆虫生理状态(例如,雌雄、交配状态、虫龄和暗适应)的影响。
影响昆虫趋光行为的环境因素主要是光属性和周围条件。即使昆虫暴露在相同的光波长下,由于光照强度和曝光时间长短的不同,昆虫的趋光行为水平也会有所不同。趋光昆虫对光具有偏好,微弱的光照便可引起昆虫较高的趋光响应。例如,蚊子可以探测到较远距离的光源,并朝向光源飞行,但当它们离光源太近时,感知到高强度的光,便飞离照明区域。此外,人造光的偏振性也会影响某些昆虫的趋光行为。许多陆生和水生昆虫更喜欢非偏振光,同种属性的光对不同属种的昆虫的吸引力也不相同。例如,一些摇蚊会更易被偏振光所吸引。
天气也会影响昆虫的趋光行为。利用LED诱虫灯捕获稻飞虱[如白背飞虱(Sogatella furcifera)和褐飞虱(Nilaparvata lugens)]会受温度、湿度、降水和风等天气条件的影响。
昆虫的趋光行为也受光源类型和外部形状的影响。诱虫灯捕捉到的昆虫种类的多样性和丰富程度可根据光源的类型而改变。一些研究表明,昆虫被不同类型光源所吸引的程度因昆虫种类而异。蜉蝣科昆虫易被LED灯吸引,与LED灯相比尺蛾科昆虫更喜欢金属卤化物灯和高压钠灯。即使诱虫灯中的光源相同,诱虫灯的效能也会因光源的外部形状不同而不同。研究表明,3种波长相同但外部形状(半球形、管状和锥形形状)的LED诱虫灯表现出明显不同的诱虫效果,其中锥形LED灯是这些诱虫灯中诱虫效果最好的。
昆虫的趋光行为受其生理状态的影响显著。新羽化的成虫趋光反应较弱,成虫的趋光反应强度随着虫龄的增长先增加后下降。当有翅豌豆蚜(Acyrthosiphon pisum)暴露于光源时,它们的趋光行为随发育阶段的变化而变化;无翅豌豆蚜对红光和绿光更敏感,1日龄的有翅豌豆蚜对蓝光更敏感。此外,在1至5日龄的梨小食心虫(Grapholita molesta)和黏虫中,3日龄虫对绿色(520 nm) LED灯光的正向趋向反应最强烈。
性别差异是影响昆虫趋光行为的重要因素。一项研究指出蝴蝶的性别差异显现出了对不同波长光线的敏感性差异。另一些研究报告说明,蛾类的两性差异导致其在光趋向反应中产生巨大差异。一项关于在森林中使用诱虫灯捕捉昆虫的研究发现,被捕的大型鳞翅目昆虫中雄性居多,并且这种两性之间丰度差异取决于物种的不同。昆虫两性间趋光反应的差异可能是由于复眼结构、小眼数量和体重差异造成的。
一些昆虫交配前的雄性与已交配的雄性相比,前者表现出强烈的趋光行为。然而,已交配雌性会比未交配的雌性表现出更强的趋光行为。例如,未交配雄性和已交配雌性黏虫的光趋向反应高于其他黏虫。对被诱虫灯捕获的棉贪夜蛾(Spodoptera littoralis)数量分析表明,虽然捕获的雄性数量比雌性多,但大多数被捕雌蛾已经交配。造成这种情况的原因可能是雄性飞蛾需要以灯光为导航寻找交配的雌性,而已交配的雌性飞蛾需要以灯光为导航寻找合适的产卵地点。
昆虫的趋光反应受其暗适应状态的影响。到达昆虫复眼光感受器的光通量受筛选色素颗粒运动的调节。昆虫在黑暗环境中眼中的色素在结晶锥之间的周边收缩,而在有光的环境中,眼内的色素是扩散的。筛选色素的运动和分布与昆虫对光谱敏感性密切相关。当梨小食心虫暴露在绿光下时,趋光反应根据适应黑暗所需时间的长短而不同。与适应黑暗需要较短(1 h)和较长时间(3 h和4 h)的蛾类相比,绿色LED光源对适应黑暗需要2 h的黏虫的吸引力最强。暗适应时间可能与从光适应眼睛转变为暗适应眼睛所需的时间有关,具有完全暗适应眼睛的昆虫的光谱灵敏度高于其他昆虫。虽然很少有科学证据清楚地证实这些因素对昆虫趋光行为的影响,但这些研究对今后昆虫趋光行为应用提供了确切可靠的依据。
3 昆虫在不同波长光照下的行为反应
人造光吸引昆虫的程度取决于光源的光谱组成。许多研究人员试图发现那些目标害虫最为敏感的人造光源波长。表1给出了一些害虫发生趋光行为的敏感波长。许多昆虫能够感知单色波长,人造光的波长是决定害虫行为光反应水平的关键因素。一项将短波长和长波长光源对蛾类的吸引力进行比较的研究报告显示,相对于长波长光源,短波长光源会使蛾类产生更加强烈的趋光反应,从而吸引到更多种类和数量的夜蛾。能让大多数害虫产生光趋向行为的波长为280~400 nm紫外光和400~700 nm可见光;很少有害虫会对长波长光(如红外光)发生趋光反应。表1中的大多数趋光害虫敏感波段集中在紫外光(350~390 nm)、蓝光(400~470 nm)和绿光(505~575 nm)波段。
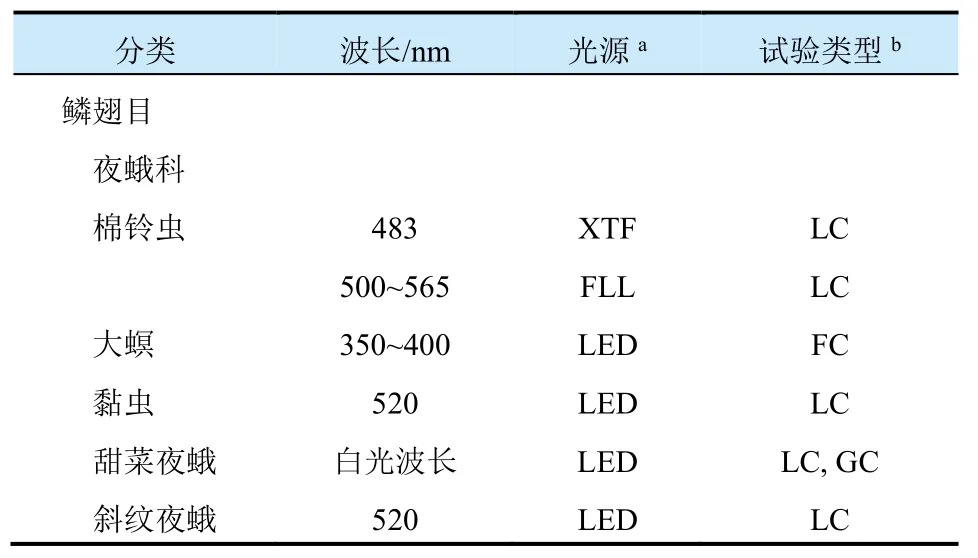
表1 多种害虫趋光反应的敏感波长
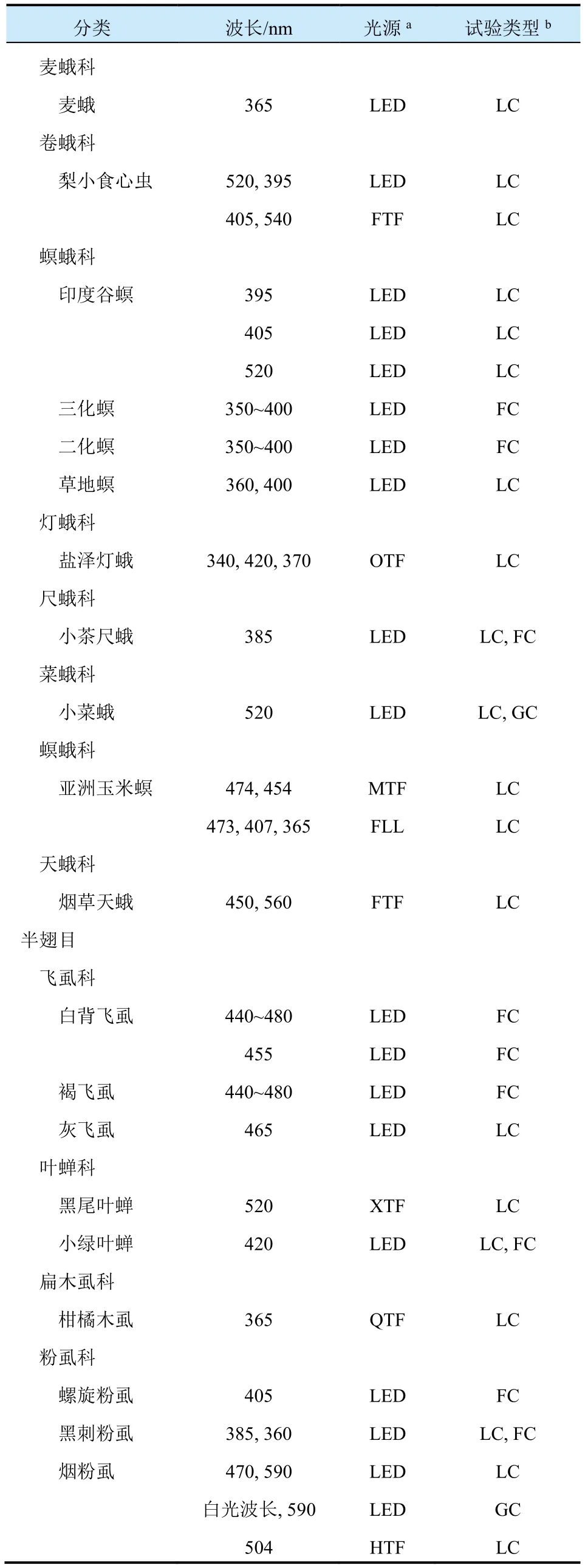
续表

续表

续表
不仅害虫会产生正向趋光行为,益虫同样也会发生趋光行为。一些益虫的正向趋光行为的敏感波长见表2。与害虫一样,益虫也对3个波段光较为敏感:紫外光(350~390 nm)、蓝光(400~470 nm)和绿光(505~575 nm)。

表2 多种益虫趋光行为的敏感波长
总的来说,夜间人造光对昆虫的吸引力取决于光源的类型和昆虫种类。对装有不同波长光源的诱虫灯捕捉到昆虫进行研究发现,紫外光(320~390 nm)对半翅目和直翅目的益虫和害虫吸引力最强;绿光(500~560nm)受到鳞翅目昆虫的青睐;蓝光(350~490nm)受到鞘翅目的青睐;黄光(555~610nm)受到双翅目的青睐。表1和表2中的数据表明,鳞翅目对紫外和绿色波长光有偏好,半翅目对蓝光和绿光有偏好,鞘翅目对紫外和紫光有偏好,双翅目对绿光的偏好略高,膜翅目对蓝光和绿光的偏好较高。昆虫的这些偏好与进化多样性密切相关,特别是与眼睛内外部结构和功能有关。
在确定目标昆虫对特定波长光敏感性的时候,应当考虑外部条件、昆虫行为特征和生理状态等多种因素。此外,应使用对目标害虫有特异吸引性的诱虫灯,达到治理害虫和保护非目标昆虫的目的。
4 昆虫对光的生理应激反应
昆虫在一定波长的光照下会产生趋光行为,对昆虫光生物反应的研究有助于解释趋光行为的机制。近几十年来,为了探索昆虫的趋光机制,许多人员对不同光照下昆虫的生理反应进行了研究,包括生化、内分泌和分子、蛋白质组学和适应度变化。特定波长的光可引起昆虫的光胁迫反应,甚至对昆虫器官(如视网膜)造成损害,光对昆虫的胁迫水平是具有物种特异性的或波长特异性的。例如,长期紫外线照射可导致昆虫细胞活性氧水平(ROS)的增加,而由ROS引起的应激可导致昆虫死亡。同时,特定波长的光可能会有利于生活在这些光环境下的某些昆虫的发育和延长昆虫寿命。
昆虫体内的抗氧化酶的活性因光波长和暴露时间的不同而不同,昆虫也会对光照做出相应的应激反应。许多昆虫易被紫外(320~380 nm)光源所吸引,所以许多研究集中研究紫外波段光的影响。在紫外线照射下,棉铃虫体内的抗氧化酶的活性,包括超氧化物歧化酶、过氧化氢酶、过氧化物酶和谷胱甘肽S-转移酶,以及总抗氧化能力和蛋白质羧基含量随暴露时间的增加而显著增加;而蛾类体内乙酰胆碱酯酶和解毒酶(如羧酸酯酶)的活性会降低。这些结果表明,紫外线照射会导致昆虫DNA、蛋白质和脂质的损伤。此外,光胁迫不仅会引起昆虫不自觉地向光冲,还会引起昆虫持续兴奋,进而死亡。
诱虫灯是否能捕获昆虫可能取决于光源诱导昆虫的氧化应激水平。有研究探讨了不同波长诱虫灯捕获的地下害虫数量与这些害虫的氧化应激水平之间的关系。该研究发现,如果诱虫灯装有的光源引起的氧化应激水平较弱,则会吸引更多的地下害虫,如鳃金龟科昆虫。一些研究报告显示,在绿光(520 nm)下黏虫的氧化应激反应较弱,而在紫外光照射下则较强。因此,黏虫对绿光具有更强的光趋向行为。然而,仍需要更多的科学依据来验证昆虫的趋光行为是否是由于光胁迫而被动产生的。
特定波长的光可以影响昆虫体内某些基因的表达。当棉铃虫(H.armigera)成虫暴露于 UV-A(320~400 nm)光下时,蛋白质组学分析表明,在检测的1 200多个蛋白点中,其中12个上调,21个下调。当雄性和雌性棉铃虫暴露于UV-A光线下30~90 min,体内的Cu/ZnSOD、CAT和GPX等几种抗氧化酶基因的表达水平显著上调;但较长的暴露时间不会导致mRNA表达水平的升高。一些研究表明,光线刺激昆虫的复眼,会使昆虫视蛋白酶基因表达,视蛋白酶基因可能在趋光反应中起重要作用。甜菜夜蛾(Spodoptera exigua)的3个视蛋白基因(Se-uv、Se-bl和Se-lw)在白天的表达水平高于夜间,具有Se-lw基因(与昆虫复眼中的长波长感光器有关)的昆虫在绿光下的趋光行为有所抑制,这表明Se-lw在甜菜夜蛾的趋光行为中起着重要作用。在紫外光、蓝光和绿光照射6 h后,棉铃虫体内紫外蛋白酶基因表达水平上调,并且雄性的基因表达水平高于雌性。此外,甜菜夜蛾和棉铃虫的饥饿会导致这些基因的表达下调,这表明昆虫的趋光行为可能受营养状况的影响。当黏虫暴露于不同的光线条件下,参与光转导、昼夜节律、嗅觉转导和味觉转导的基因表达模式发生了不同的变化。同时,紫外线照射引起了与应激反应相关基因表达水平的增加,如热休克蛋白(HSP)和细胞色素P450家族。同样,在紫外线照射下果蝇(Drosophila melanogaste)体内的11个热休克蛋白基因表达显著增加。毫无疑问,这些相关的分子研究将增加人们对昆虫趋光行为的理解。
光在昆虫的生活中必不可少,正常的光周期可以促进昆虫的正常发育。在昆虫生存环境中,光周期的光部分可以在夜间由人造光提供。夜间常见的人造光与昆虫生命中的光周期密切相关,并且某些波长的光照对昆虫的孵化、发育、滞育、死亡率、繁殖力和寿命等生物学特性有很大的影响。有研究表明,昆虫的生长可能受到夜间(或暗期)某些光照的影响,生物参数水平的变化可能取决于光的波长和照射期。在几种不同波长的LED光下饲养黏虫,与对照组(14 h∶10 h光/暗)相比,白光组(24 h∶0 h光/暗)的黏虫在不同发育阶段的所有测量生物特性均受到负面影响。与对照组相比,在白光组中,卵的孵化率、幼虫的发育周期和蛹的羽化率没有产生明显变化,但幼虫存活率、化蛹率和成虫存活率明显降低。在紫外光(365 nm)下饲养棉铃虫,棉铃虫卵的孵化率和孵化期,幼虫的存活和发育期、蛹的羽化率和寿命均受到负面影响,这与紫外线照射时间延长有关。虽然这些昆虫的F1代在卵期受到负面影响,但幼虫、蛹和成虫期没有受到显著影响。当在几个不同光波长下饲养家蝇(Musca domestica)和大头金蝇(Chrysomyia megacephala)时,当波长为435 nm时,二者卵的孵化率、幼虫存活率和化蛹率会下降。
此外,光的波长会影响生活在其中的成虫的繁殖能力和寿命。与对照组相比,夜间在绿光(520 nm)下饲养的雌性棉铃虫的繁殖力和寿命显著降低。橙光(600 nm)对迟眼蕈蚊(Bradysia coprophila)求偶、交配行为和繁殖不利,而455 nm蓝光与600 nm橙光有相反的影响。此外,冬尺蛾(Operophtera brumata)的交配可以被人造光抑制,特别是红光和绿光。
在一定波长光下生活的成虫的生殖参数和寿命可以相互影响(例如,如果雌性的繁殖力增加,雌性成虫的寿命就会缩短)。当黏虫每天暴露于紫外线(365 nm)下1 h,其雌虫的繁殖力和产卵率与对照相比显著提高,但寿命降低。与对照组相比,每天暴露在紫外线(365nm)下5 h的雌性黏虫的繁殖力和产卵率显著提高,但雄性和雌性黏虫的寿命都显著降低。
在夜间一定波长光线下生活的昆虫,其生物学特性的变化可能与幼龄虫激素(JH,参与发育、滞育、繁殖和寿命)的调节有关。随着暴露时间的延长,在紫外光下饲养的棉铃虫的JHS (如JHI、JHII和JHIII)水平逐渐降低,表明在紫外环境中饲养的昆虫的生物学特性可能会随着JHS的调节而改变。一些研究人员推测,生活在紫外线下的一些蛾类的生物参数的负变化可能是由于紫外线照射引起的难以忍受的光胁迫所致。然而,昆虫的生物学特性为什么会受到特定光照的影响,目前尚不清楚,需要更科学和更有说服力的证据来解释这一现象。
5 诱虫灯应用方面存在的问题
鉴于保护生物多样性的需要和IPM目标,在使用诱虫灯进行害虫治理的过程中面临一个重要问题:如何设计使用一种只吸引靶标害虫(或吸引很少的非靶标昆虫)的诱虫灯。诱虫灯的原理是基于昆虫对光的正向趋光行为,许多诱虫灯已被开发并应用于趋光性害虫的监测和防治。然而,在实践中使用的光源(包括高压汞灯、荧光灯和白炽灯)也会吸引有益的昆虫,如害虫天敌。因此,开发一种波长范围窄、靶标害虫敏感的光源具有重要意义。LED灯具有许多优点,例如,波长范围窄,功率低,发光效率高,温度低,体积小,成本低,寿命长;还可以以太阳能电池作为电源,因此目前LED灯是诱虫灯光源的最佳选择。现在已经开发了一些能发出特定波长光的LED诱虫灯用于监测和治理虫害。茶树的主要害虫茶叶蝉(Empoasca onukii)和茶毛虫(Ectropis obliqua)分别对420 nm和385 nm的光有较强的敏感性,茶园中还有10种益虫对380 nm的光源较为敏感。一个装有双波长(385 nm和420 nm) LED光源的诱虫灯,已被用来治理茶叶害虫。实践表明,此款LED诱虫灯可以更多地吸引2种害虫,与带有荧光灯的诱虫灯相比,双波长LED诱虫灯捕获天敌益虫的量明显减少。户外使用的装有LED (470 nm、15 000 mcd、20~30°、20 mA、3.2 V)的Silva诱虫灯,与装有白炽灯泡(4 W、6.3 V、CM-47)的商用CDC诱虫灯相比,前者吸引了更多的库蚊(Culexspp)。此外,在野外装有UV-LED的诱虫灯捕集的蚊子数量高于装有白炽灯泡的CDC诱虫灯、白光LED诱虫灯和可产生CO2(酵母产生)的诱虫灯捕获的蚊子数量。
关于昆虫对发出特定波长光的LED灯的趋光行为的研究正在进行中,使用发出特定波长光的LED诱虫灯可以监测和治理农业和森林害虫种群,可作为未来IPM战略中害虫治理的重要手段之一。
一般来说,翅膀较大的蛾类对光的敏感性更高。一项研究分析了装有378种不同光源的诱虫灯捕捉的蛾类的翅膀,结果表明,诱虫灯捕捉的飞蛾数取决于光源的类型和飞蛾翅膀的类型。
诱虫灯应在适当的时间使用,例如在目标害虫量较多或非常活跃的时候。大多数鳞翅目害虫成虫表现出趋光性,但其爆发期因种类的不同而不同。此外,不同种类的趋光昆虫被诱虫灯捕获的时间不同,这可能与每种昆虫在夜间的昼夜节律有关。研究报告指出,一些种类的昆虫在夜间特定时间段会更容易被诱虫灯所吸引。图1为一些害虫在夜间易被诱虫灯捕获的时间段。根据图1可知,许多害虫主要在18时至24时被诱虫灯捕捉,有一些在01时至05时期间会再次被捕获,如图2所示,能发出不同波长光的LED诱虫灯也会在不同时间捕捉到一些种类的害虫。这些结果表明,昆虫在夜间的活动时间可达24:00,在上述2段时间使用诱虫灯可能会更有效。从保护生物多样性的角度看,应在适当的时间使用发出特定波长光的诱虫灯,从而使这些诱虫灯能更好的吸引目标害虫。

图1 夜间诱虫灯捕获14种昆虫的时间

图2 夜间3种不同波长的诱虫灯捕获11种昆虫的时间
诱捕色卡和昆虫性信息素也是目前常用的害虫诱捕手段。例如,可利用黑色诱虫板诱捕迟眼蕈蚊(Bradysia coprophila),利用性引诱剂干扰梨小食心虫(Grapholita molesta)的交配。为了提高诱捕害虫的效率,诱虫灯可与这2种方法结合使用。诱虫灯与彩色卡片的结合使用已被用于监测和防治温室中的害虫。相对于单一的诱捕方法,装有黄色黏虫板的绿光LED诱虫灯能吸引更多的害虫(包括迟眼蕈蚊(Bradysia coprophila)、烟草白蝇(Bemisia tabaci,B.argentifolii)、西花蓟马((Frankliniella occidenta)和叶蝉(Empoascaspp))。同时这种结合诱捕法对有益昆虫如寄生蜂的吸引力更低。诱虫灯与昆虫性信息素的结合使用可以捕获更多的昆虫成虫。使用装有绿光(520 nm) LED灯和性信息素的诱虫灯,相对于单独使用这2种方法,将会吸引更多的雄性甘薯象鼻虫(Euscepes postfasciatus)。
同时,在诱虫灯使用过程中应考虑其结构和高度,在增加害虫的捕获量的同时保护有益昆虫。悬挂在茶园2 m高的装有吸力风扇的锥形LED(385和420 nm)诱虫灯,与对照组(配备电网的白色荧光灯诱虫灯)或配备LED灯和水盆(或电网)的其他捕虫器相比,装有吸力风扇的锥形LED诱虫灯会捕捉到更多的害虫。与单一方式捕虫器相比,结合型捕虫器捕获的有益昆虫数量反而更少。
在筛选诱虫灯光源波长的时候,要考虑该波长是否能高效吸引害虫,同时还要考虑对作物生长是否有负面影响。
6 结论和展望
早在很久以前人们对昆虫的趋光行为就有所认知,例如,在古代人类就会利用火把来吸引害虫。在过去的几十年里,科学家们研究了昆虫趋光行为的性质和机制,并在实践中应用了装有特定光源的诱虫灯。虽然昆虫趋光行为的确切原因目前尚不清楚,但诱虫灯的使用早已在害虫物理防治中发挥了不可替代的作用。此外,LED灯的使用将有助于诱虫灯的发展,在IPM中使用诱虫灯进行害虫物理防治正引起世界范围的广泛关注。今后,通过对昆虫与光照之间生物学关系更加深入的研究,将会逐渐明确昆虫趋光行为的原因,从而推动诱虫灯的使用朝更有效、更环保、更经济的方向发展。
猜你喜欢
杭州电子科技大学学报(自然科学版)(2022年3期)2022-06-08
家庭影院技术(2021年9期)2021-11-05
阅读(科学探秘)(2021年8期)2021-09-01
上海工艺美术(2021年4期)2021-04-24
家庭影院技术(2021年2期)2021-03-29
意林·少年版(2016年11期)2016-09-10
扬子江(2016年1期)2016-05-19
祝你幸福·知心(2016年3期)2016-03-29
学苑创造·A版(2009年5期)2009-06-29
故事大王(2009年5期)2009-06-02
