
深层土壤水分对黄土塬区旱作冬小麦耗水的贡献
2020-11-24 01:37程立平王亚萍刘文兆
干旱地区农业研究 2020年5期
程立平,林 文,王亚萍,刘文兆
(1. 平顶山学院,河南 平顶山 467000;2. 山西农业大学农学院,山西 太谷 030801;3. 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100)
黄土高原南部的黄土塬区是我国北方重要的旱作冬小麦产区,基于其独特的地形、水文、气候等环境条件,冬小麦所能利用的水源只有降水和土壤水,土壤供水可占冬小麦生长期耗水量的一半左右。土壤水库调蓄水分功能可有效缓解降水不足和季节分布不均所导致的旱情,对于该区冬小麦生长发育至关重要,特别是在拔节-抽穗-灌浆期的高需水阶段[1-2]。良好的底墒和土壤水库的调蓄功能对于提高旱作冬小麦水分利用效率、稳定冬小麦产量具有重要的意义。已有的研究多将0~2 m或0~3 m土层储水作为整体,研究其对冬小麦耗水及产量形成的贡献[2-4],而不同土层土壤储水对冬小麦耗水的贡献,尤其是深层土壤水的贡献尚不明确。有研究通过对比发现,黄土塬区高产麦田0~5 m深度范围内土壤水分含量低于长期休闲地,并结合根系分布推测冬小麦根系直接吸水层为3 m,3 m以下土壤水分减小是水分上移的结果[5];但是小麦高产所形成的土壤干层能使降水入渗深度显著变浅[6],深层土壤补给来水减少,亦有可能使高产麦田3 m以下范围内土壤湿度降低。可见黄土塬区冬小麦能否利用3 m以下土壤水分缺乏直接证据,鉴于此,本文拟回答以下2个问题:(1)黄土塬区冬小麦生育过程中是否利用3 m深度以下土壤储水?何时利用?(2)不同深度土层土壤水,尤其是3 m深度以下土壤水对冬小麦耗水的贡献是多少?
稳定同位素技术为解决以上两个问题提供了可能。植物根系从土壤中吸收水分和水分在植物体内运输到达叶片或幼嫩未栓化的枝条之前均不发生同位素分馏[7-8],因此植物木质部水同位素组成是其所利用水源同位素组成的综合体现。要定量研究植物对各水源的利用比例,可采用二源或三源线性混合模型[9],但该模型不能用于确定多于3种以上水源的利用比例,而IsoSource多源混合模型[10]可用于估算多种水源对植物用水的贡献比例范围。随着同位素示踪技术的发展与普及,已有学者开展了小麦水分溯源的试验研究。苑晶晶等[11]利用δ18O信息揭示了黄河下游引黄灌区不同灌溉条件下冬小麦对土壤水和地下水的利用比例;张丛志等[12]研究表明封丘地区冬小麦对地下水以及不同深度土壤水的利用比例随着生育期的变化而变化。但是上述两项研究均在有灌溉条件且地下水埋深较浅的地区进行,杜俊杉等[13]则选择了地下水埋深远大于地下水蒸发极限水位埋深的北京大兴地区,确定了0~2 m土层范围内不同深度土壤水对冬小麦耗水的贡献。安江龙等[14]应用3 m塑料管土柱法在田间进行冬小麦种植试验,并应用同位素溯源发现冬小麦自灌浆期起开始明显利用2~3 m土层土壤水。程立平等[15]初步研究了黄土塬区底墒充足条件下冬小麦拔节期和抽穗期的水分利用规律,但其结果仍需进一步验证。
旱作冬小麦对深层土壤水分的利用是土壤水库调蓄的重要体现,在黄土塬区土壤—植物—大气连续体水分运动与平衡中占据重要位置,是旱作农业研究的重要内容。本试验首先对3 m土层土壤水分人工标记氘水(D2O),并通过测定小麦茎秆水δD明确冬小麦是否利用3 m深度土壤水分;其次通过测定小麦茎秆水、降水和不同土层土壤水δ18O值,利用二源线性混合模型和IsoSource多源混合模型确定冬小麦对降水、不同土层土壤水的利用比例。本研究对于明确冬小麦水分利用规律,进一步推进旱作农田生态系统土壤水分循环研究具有重要理论意义和实际意义。
1 材料与方法
1.1 研究区概况
研究在中国科学院长武农业生态试验站(107°40′ E, 35°14′ N)进行。试验地点位于陕西省长武县所在的黄土塬上,塬面平坦,海拔1 220 m。研究区属暖温带半湿润易旱大陆性季风气候,多年平均降水量580 mm,降水年际变化较大,年内分布不均,7—9月降水量占全年降水总量的55%以上。研究区黄土堆积深厚,土壤为黑垆土,田间持水量和凋萎湿度分别为22%(质量含水量,下同)和9%,容重为1.3 g·cm-3,母质为中壤质马兰黄土,土质疏松,孔隙度在50%左右,通透性良好,利于水分入渗和植物根系生长。研究区地下水埋深40~80 m,不参与土壤—植物水分循环。
1.2 样品采集与测定
试验于2016年5—6月进行。选择长势良好的小麦地(品种为长旱58),于5月9日在地块中央位置约1 m2样方处,利用土钻于四角处钻至3 m深度,共注入氘水50 mL进行人工标记,然后回填。
1.2.1 土壤样品采集 分别于2016年5月9日(抽穗期)、5月19日(开花期)、5月29日(灌浆期)和6月10日(乳熟期),在人工标记样方内使用土钻采集土壤样品,采样深度4 m,间隔20 cm采样1次。因标记样方面积小,为减少扰动,每次仅取1个土壤剖面。土壤样品分为2份,1份密封保存带回实验室利用低温真空抽提系统Li-2000抽提土壤水样;1份装入铝盒利用烘干法测定土壤含水量。将抽提的土壤水样密封于水样瓶中,编号、记录并冷藏待测。
1.2.2 小麦茎秆水样采集 分别于2016年5月9日、5月13日、5月19日、5月24日、5月29日、6月4日和6月10日,围绕人工标记样方采集冬小麦样品。采集方法为随机选择2~3株长势良好冬小麦,连根拔起采集近根部的茎秆,剔除叶鞘部分,快速装入10 mL玻璃瓶内密封保存,每次采集3瓶作为重复,实验室内利用低温真空抽提系统Li-2000抽提小麦茎秆水样。将抽提的植物水样密封于水样瓶中,编号、记录并冷藏待测。
1.2.3 降水样品采集 利用人工雨量筒收集次降水水样,并记录降水量。
1.2.4 氢氧稳定同位素测定 小麦水样δD值测定:采用LGR IWA-45EP液态水同位素分析仪测定抽提小麦水样的δD值,分析精度为±0.5‰。
土壤、小麦和降水水样δ18O值测定:采用质谱仪Finnigan MAT253,TC/EA法测定水样内δ18O值,分析精度为±0.3‰。
所有水样测定结果以标准平均海水(SMOW)为标准的千分偏差表示。
δD(或δ18O)=[(Rsa-Rst)/Rst]×1000‰
式中,Rsa和Rst分别表示样品和标准物中稳定性氢同位素D/H或稳定性氧同位素18O/16O的比率,δ值越小表明重同位素越贫化,δ值越大表明重同位素越富集。
1.3 数据统计与分析
利用二源线性混合模型和IsoSource多源线性混合模型(V1.3.1)计算不同水源对冬小麦耗水的贡献比例,IsoSource模型参数中的来源增量(source increment)设置为2%,质量平衡公差(mass balance tolerance)为0.1%。
采用Excel 2007软件进行数据处理与制图,利用单因素方差分析比较不同时期土壤水同位素组成差异显著性。
2 结果与分析
2.1 土壤水分剖面动态变化
由图1可见,从抽穗期至乳熟期,小麦样地0~4 m土壤水分剖面随发育时间的推进而向左移动,土壤水分含量逐渐减少,其中1~2 m土层内土壤含水率减幅较大,而0~1 m土层范围内减幅较小,尤其是0.5 m以上土层范围内几乎无变化。推测原因有以下3点:一是0.5 m以上土层内土壤含水率与萎蔫系数持平,可以认为该层范围内土壤水已不能为冬小麦所利用,冬小麦只能利用0.5 m以下土壤水;二是土壤水分含量低于毛管断裂含水量(15%[16]),土壤蒸发微弱;三是研究时段内降水较少,5月1日—6月10日共记录到13个降水日,但是降水量大于5 mm的有效降水仅6 d(图2),最大降水量仅为13.6 mm,很难入渗至20 cm深度土层,而本试验则是从20 cm深度起测定土壤水分。

图1 冬小麦样地土壤水分剖面动态变化Fig.1 Dynamics of soil water profiles underwinter wheat plot

图2 研究时段内降水量及降水δ18O值Fig.2 Precipitation and its δ18O values during study period
2.2 冬小麦茎秆水样δD值变化
尽管液态水同位素仪LGR在测定植物水δD值时存在甲醇或乙醇类有机污染[17],但是其测试结果仍然可以为冬小麦是否利用了人工标记的土壤水分提供有效信息。5月9日、5月13日和5月19日冬小麦茎秆水δD平均值分别为-63.67‰、-58.66‰和-60.46‰,无明显变化;但是从5月24日起冬小麦茎秆水δD值明显变大(表1)。δD值变化表明5月19日之前,冬小麦并未利用标记在3 m深度处的氘水,但是从5月24日起冬小麦已经能够利用这部分水,从而导致茎秆水δD值变大。可见冬小麦从5月24日起开始利用3 m深度以下土壤水分,其所占比例则需进一步研究。
表1 不同采样日期冬小麦茎秆水样δD值/‰
Table 1 D values of winter wheat stem water samples in different dates

表1 不同采样日期冬小麦茎秆水样δD值/‰
重复样品 Duplicate sample05-0905-1305-1905-2405-2906-0406-101-66.83-55.35-61.31150.87-39.69-41.20-2-59.79-61.31-62.87-54.52--34.17-47.503-64.40-59.30-57.19-60.55-38.28-33.77588.56
注:-表示水样中氢同位素太富集未能测出。
Note:- means that hydrogen isotope in water sample was too rich to be determined.
2.3 不同来源水样δ18O值变化
黄土塬区地下水埋藏极深而不参与土壤-植被系统水分循环,因而冬小麦能够利用的水源只有降水和土壤水。受蒸发效应影响,浅层土壤水分重同位素富集[18]。由图3可见,土壤水δ18O值在20 cm处最大,20~280 cm深度范围内土壤水δ18O值随深度增加而逐渐减小,280 cm土层以下土壤水δ18O值随深度增加而逐渐增加;4个时期土壤水δ18O平均值分别为-12.17‰、-12.03‰、-12.49‰和-12.70‰,方差分析表明4个时期之间并无显著差异。蒸发和降水入渗混合作用是引起土壤水同位素值变化的主要原因[19],结合2.1部分可知研究时段内土壤蒸发微弱,降水量小难以入渗至20 cm深度处,因而土壤水同位素值无显著时间变化。
研究时段降水δ18O值(图2)与多年降水δ18O平均值-10.38‰[20]相比明显偏大,原因在于长武黄土塬区降水同位素组成具有冬春富集、夏秋贫化的季节变化特征[19-20]。降水只有转化为土壤水才能被植物利用,研究时段属于旱季,降水量小且很难入渗至20 cm深度,降水转化的土壤水集中在0~20 cm深度范围以内,而小麦根系密度在这个深度范围以内最大,能够直接利用降水转化而来的土壤水。本研究中土壤水同位素采样是从20 cm开始的,故将降水作为一个独立水源。已有研究表明干旱地区植物体在降水之后的7 d内仍保存有大量的降水[21],因此本文分别利用5月7日、5月14日、5月27日和6月9日降水δ18O值来计算抽穗期、开花期、灌浆期和乳熟期降水对冬小麦耗水的贡献率,这4日降水量分别为7.7、13.6、5.5 mm和6.4 mm(图2),降水δ18O值分别为-6.03‰、4.72‰、6.57‰和5.67‰(图3)。
植物根系吸收水分过程一般并不发生同位素分馏现象[7-8],因此小麦茎秆水的同位素组成是其所利用水源同位素组成的综合反应。由图3可见,4个生育期小麦茎秆水同位素组成均介于降水和土壤水同位素组成之间,其δ18O值分别为-9.16‰、-9.82‰、-10.37‰和-10.02‰。
2.4 降水和土壤水对冬小麦耗水贡献率的模型预测
研究时段内样地0.5 m土层以上土壤含水率低于或与萎蔫系数持平,因而冬小麦仅能吸收该深度以下土壤水分;由冬小麦茎秆水样δD值变化可知,冬小麦在5月9日和5月19日并未利用3 m深度土壤水分,在5月29日和6月10日则利用了3 m深度土壤水分。因此,本文根据土壤水同位素剖面分布特征,在抽穗期和开花期将50~300 cm土层分为4组,分别是50~100, 100~150, 150~200 cm和 200~300 cm;在灌浆期和乳熟期将50~400 cm土层分为5组,分别是50~100, 100~150, 150~200, 200~300 cm和300~400 cm;然后将冬小麦不同生育期茎秆水、降水以及各组土壤水δ18O值分别输入IsoSource模型,计算各水源对冬小麦耗水的贡献率。
IsoSource模型最终结果不是唯一解,而是依据计算过程中各水源相对贡献率的频率确定可行的概率分布组合[10],即冬小麦对各水源利用比例的分布范围,但是也可利用各潜在水源相对贡献率的频率加权平均值作为唯一解的体现。图4给出了不同生育期降水和各组土壤水分对冬小麦耗水的平均贡献率,其中误差线代表了各组水源相对贡献率的分布范围。
从图4看出,无论是降水还是不同土层土壤水对冬小麦耗水贡献率,IsoSource模型结果分布范围都较宽,从而给确定不同水源的贡献率带来一定的不确定性。为了减小这种不确定性,本文利用二源混合模型计算降水和土壤水对冬小麦耗水的贡献率,结果如表2所示。二源模型计算结果为唯一解,但是其土壤水贡献率为总贡献率,不能得出分层土壤水的贡献率,因此本文利用二源模型结果对IsoSource模型结果进行校对,以确定不同深度土层土壤水对冬小麦耗水的贡献率。
对比2种模型所得降水贡献率可知,4个生育期二源模型结果均分布于IsoSource模型结果的分布范围之内(图4,表2),表明两者数据相符,所得结果可信。但是与多源模型唯一解形式(平均贡献率)相比,二源模型计算所得降水贡献率偏高,土壤水贡献率偏低,4个生育期差值分别为9.6%、0.5%、4.8%和2.1%。据此对IsoSource模型计算所得的不同土层土壤水平均贡献率进行校对,具体方法为以平均贡献率的大小为权重,对差值进行分配,结果见表3。

图3 不同生育期冬小麦茎秆水、土壤水和降水中δ18O值Fig.3 δ18O values of winter wheat stem water, soil water, and rainwater at different growing stages

注:柱高度代表平均贡献率,误差线代表贡献率分布范围. Note:Column height represents the mean value of contribution rate and bar represent the range of contribution rate.图4 不同生育期降水和各层土壤水对冬小麦耗水的贡献率Fig.4 Contribution rates of rainwater and soil water in different soil layers to winterwheat water consumption at different growing stages
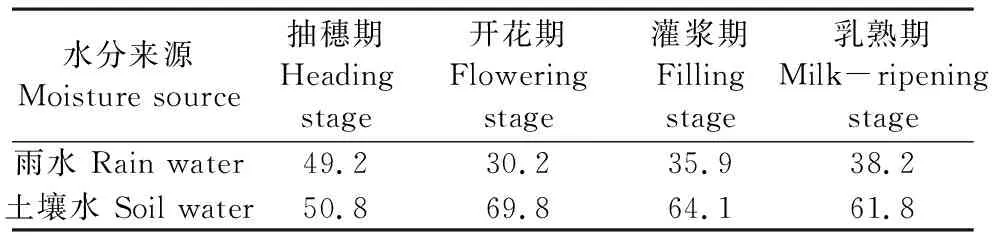
表2 各生育期雨水和土壤水对冬小麦耗水的贡献率/%

表3 各生育期不同土层土壤水对冬小麦耗水的贡献率/%
综上可得从抽穗期至乳熟期,降水对冬小麦耗水的贡献率在30.2%~49.2%之间,平均值为38.4%,贡献率大小取决于小麦采样日期距离上次降水日期的远近以及降水量的大小[21]。而土壤水对冬小麦耗水的贡献率总体上表现为随土层深度增加而减小,50~100 cm土层土壤水平均贡献率为20.2%;100~150 cm土层土壤水贡平均献率为15.4%;150~200 cm土层土壤水平均贡献率为11.1%;200~300 cm土层范围土壤水平均贡献率为9.3%。冬小麦自灌浆期起开始利用3 m深度以下土壤水,5月29日和6月10日利用量分别占其总耗水量的11.4%和11.5%。
3 讨 论
3.1 冬小麦根系吸水率剖面变化特征
冬小麦根系吸水率在土壤剖面上的分布状况是由土壤含水量的垂直分布和根密度分布状况所决定的[22]。相关试验表明研究区冬小麦返青期根系已生长至250 cm深度,至收获期小麦根系最深可达到320 cm[1],但是冬小麦根长密度具有随土壤深度增加而减小的变化特征,即深层剖面根系分布较少。研究区冬小麦拔节期前土壤水分含量较高且剖面分布均匀[23],在这种条件下根系吸水率的分布与根密度的分布规律相同,吸水率随着深度的增加而降低,吸水率大的地方土壤水分消耗较多[22],随着时间的延长,土壤含水量逐渐减少,根系吸水速率随之降低[24],高吸水区逐渐向下转移[22],同位素试验也证明了这种现象的存在[15]。根系吸水与土壤物理蒸发效果叠加,使浅层土壤水分迅速减少,图1表明黄土塬区抽穗期50 cm以上土层含水量已低至萎蔫系数。抽穗期和开花期1~2 m土层范围土壤水对小麦耗水贡献率分别为土壤水总贡献率的25.2%和32.8%,表明小麦高吸水区已转移至1~2 m土层范围;灌浆期和乳熟期1~2 m土层范围土壤水对小麦耗水贡献率有所降低,分别为24.4%和23.5%,其原因在于冬小麦已开始利用3 m深度以下土壤水,高吸水区依然在向下移动。图1可见,3 m深度以下土壤含水量丰富,但是冬小麦自灌浆期才开始利用该层土壤水分,占其耗水量的11.4%,原因在于冬小麦对某一层次土壤水分的利用不仅与土壤水分状况有关,还与该层中根量大小有关。Barraclough等[25]指出土壤中具备吸水能力的根量阈值为1 cm·cm-3,低于此值,即使土壤含水量较高, 也难以被充分利用。说明冬小麦根系可能在灌浆期前已扎至3 m深度以下,但是灌浆期以后3 m以下深度小麦根量才达到吸水阈值,从而开始有效利用3 m深度以下的土壤水分。
3.2 冬小麦土壤水分利用与土壤干层
黄土塬区高产农田下普遍存在着土壤干层现象[6,26],土壤湿度低于田间稳定湿度时(毛管断裂含水量,15%)即认为形成土壤干层[16],是该区土壤水分负平衡所导致的一种土壤水文现象。土壤干层位于降水入渗深度以下土体某一范围以内,具有一定的时间和空间稳定性,干燥化程度因植物种类和生长年限而定。研究表明农田下土壤干层分布深度一般为1~3 m[27],本研究中冬小麦生育中后期主要吸水层为1~2 m土层范围,其在2 m以上土层含水量均低于田间稳定湿度,可以推断若冬小麦主要吸水层下移至2~3 m土层,则其含水量极有可能低于稳定湿度,具备了形成土壤干层的条件。但是冬小麦主要吸水层下移深度取决于土壤底墒以及生育期降水量,若底墒充足或者降水量丰富,则浅层土壤含水量减少速度缓慢,高吸水区向下转移深度少[22, 24],否则转移深度大。土壤干燥化是冬小麦对土壤水分利用率高低的一个标志,干燥化土层厚度愈大,干燥化程度愈剧烈,说明冬小麦对土壤水分利用愈充分,土壤水库调蓄作用愈大。但是,若干层水分不能得到充分恢复,便会削弱土壤水库的供水调节能力,影响土壤水库调蓄功能的持续发挥[6]。频率分析表明,长武高产农田条件下,年降水入渗至2 m和3 m深度分别需要3 a和10 a才能重现1次[28],表明高产农田即使经过夏闲期降水补充,其深层土壤水分也未必能够恢复,从而形成土壤干层,降低土壤水库调节供水的能力。因此在生产实践中,可通过夏闲期蓄水保水等田间措施,减少土壤蒸发、增加雨水入渗量和入渗深度,促使深层土壤水分恢复,以此保证土壤水库调蓄功能的持续发挥。
3.3 农田水量平衡计算中深度的选择
黄土高原地区土壤水量平衡计算土层深度至关重要,深度选择不当会使植物与水分关系研究的结论失真[5, 23]。以往在利用水量平衡法计算农田蒸散量时多以2 m或者3 m为计算深度[6, 26],本研究表明从灌浆期起冬小麦对3 m深度以下土壤水分利用量占其总耗水量的11.4%,因此建议在农田水量平衡计算中采用4 m深度为宜,这也与长期定位监测农田深剖面水分数据计算所得结论相符[23]。
4 结 论
稳定同位素试验表明,在长武黄土塬区,从抽穗期至乳熟期降水对冬小麦耗水的平均贡献率为38.4%,土壤水的平均贡献率为61.6%。分土层来看,从抽穗期至乳熟期,50~100 cm土层土壤水对冬小麦耗水的平均贡献率为20.2%,100~150 cm土层为15.4%,150~200 cm土层为11.1%,200~300 cm土层为9.3%。随着生育期的推进,冬小麦主要吸水层逐渐向下延伸,吸水深度不断增加;冬小麦茎秆水δD值变化表明,冬小麦自灌浆期开始利用3 m以下土壤水分,300~400 cm土层土壤水对灌浆期至乳熟期冬小麦耗水的平均贡献率为11.4%。可见,黄土塬区深层土壤储水对于旱作冬小麦高产稳产具有重要意义。生产实践中,应通过夏闲期蓄水保水措施,减少土壤蒸发、增加雨水入渗量和入渗深度,促使深层土壤水分恢复,以此保证土壤水库调蓄功能的持续发挥。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年10期)2022-08-22
农业工程学报(2022年5期)2022-06-22
水土保持学报(2022年3期)2022-05-26
全球定位系统(2022年1期)2022-04-14
农业工程学报(2022年1期)2022-03-25
现代农村科技(2021年11期)2021-11-05
全球定位系统(2019年4期)2019-08-30
分析化学(2017年5期)2017-06-21
同位素(2014年3期)2014-06-13
