
夏玉米苗期对不同时长干旱-复水的生理响应机制
2020-11-24 01:36蔡焕杰
干旱地区农业研究 2020年5期
肖 钢,蔡焕杰,沐 青,赵 龙
(1.西北农林科技大学旱区农业水土工程教育部重点实验室,陕西 杨凌 712100;2.西北农林科技大学中国旱区节水农业研究院,陕西 杨凌 712100)
干旱是限制农业生产的主要非生物因素之一,提高作物抗旱性是实现农业增产的重要举措[1]。近年来研究发现,干旱并非只对植物产生负面影响,一定程度的干旱胁迫能诱导对植物的生长发育以及后期产量的补偿效应,以弥补干旱胁迫期间作物遭受到的伤害和损失[2]。玉米是三大栽培作物之一,在干旱半干旱地区,有限的水资源不足以达到夏玉米正常灌溉需求[3-4]。而夏玉米苗期是适合进行水分调控的生育期之一[5-6]。因此,研究干旱-复水对夏玉米苗期的生理调节机制对探索作物生理活动有着重要意义。渗透调节是植物适应干旱的关键过程,可以维持组织的代谢活动。脯氨酸的亲水性极强,能稳定原生质胶体及组织内的代谢过程[7]。在干旱胁迫下,叶片和茎的脯氨酸含量明显增加,以维持细胞膨压,但随着胁迫时间的延长而有所下降[8-9]。这种调节不是无限制的,在受胁迫程度大于一定范围时,植物组织的渗透调节能力下降[10-11]。干旱胁迫下植物细胞积累的活性氧破坏细胞功能,对作物生长和产量造成负面影响[12-13]。在正常条件下,植物细胞中产生的活性氧与其清除系统能够保持平衡,超氧化物歧化酶(SOD)、过氧化物酶(POD)以及过氧化氢酶(CAT)等抗氧化保护酶在清除活性氧的过程中发挥重要的作用[13-14]。而当环境胁迫长期作用于植株,产生的活性氧超出了活性氧清除系统的清除能力时,便引起活性氧累积产生氧化伤害,从而使细胞功能失常[15-16]。研究表明,丙二醛(MDA)含量在作物受到胁迫后会逐渐增大,且随着胁迫程度的加剧,丙二醛含量上升趋势更加明显[17-18]。叶片净光合速率、蒸腾速率、气孔导度以及胞间 CO2浓度反映了植株的光合能力[19-20]。研究表明,干旱胁迫在较短的时间里能够造成部分气孔的关闭,导致光合、蒸腾速率在胁迫初期就有了较大幅度的降低,随着胁迫时间的延长,干旱胁迫可能引起光合结构的异常,从而导致光合能力以及气体交换参数的持续下降[21-22];复水后,作物的光合以及气体交换能力逐步恢复,而重度干旱胁迫将对作物光合能力产生不可逆影响[23]。
本研究在夏玉米苗期进行不同时长和程度的干旱处理,以期进一步阐明夏玉米在苗期干旱-复水环境下的生理调控机制,探讨作物在干旱胁迫下光合、渗透调节以及抗氧化酶系统的动态变化过程,为实现农业生产高效用水提供理论依据。
1 材料与方法
1.1 试验区概况
试验在西北农林科技大学中国旱区节水农业研究院农田水分转化试验场(108°04′E,34°18′N)遮雨棚下进行。试验区为大陆性暖温带季风气候,海拔506 m,多年平均温度12.5℃,多年平均降水量609 mm,多年平均蒸发量1 500 mm,全年无霜期210 d。2018年夏玉米苗期内(7月)最高气温与平均气温动态变化如图1。
1.2 试验设计
供试玉米品种为吉祥1号,试验采用桶栽方式,土壤类型为中壤土,取自西北农林科技大学附近耕地0~20 cm土层。土壤砂粒(0.05~1.00 mm)、粉砂颗粒(0.05~0.005 mm)和粘粒(≤0.005 mm)分别占23.0%、30.0%和47.0%。1 m土层内的田间持水量为24%,土壤干容重为1.31 g·cm-3。试验采用长×宽×深度=30 cm×30 cm×40 cm的试验方桶,每桶种植2株玉米,于出苗后选取长势相同的定苗1株。为保证根部通气性良好,在桶底打5个直径为1 cm的小孔,铺1 cm的砂过滤层来保证透气性,为避免夏季桶内土壤温度过高,在桶四周包裹一层隔热膜;每桶装干土52.10 kg,在土的最上层铺1 cm珍珠岩以减少水分蒸发。按200 kg·hm-2的施氮量、100 kg·hm-2的施磷量,换算出每桶施尿素2.37 g,磷酸二铵4.75 g。
本试验一共7个处理,每个处理有5个重复,于2018年6月20日播种,五叶期开始进行干旱-复水处理。水分处理分别为:轻度干旱胁迫(W1):60%~70%θf(田间持水量);中度干旱胁迫(W2):50%~60%θf;W1和W2处理各设3个胁迫时长处理,分别是7 d(T1),10 d(T2),13 d(T3),其余时间充分供水。以正常供水(75%~85%θf)作为对照(CK)。控水方法为称重法,每次在早上8∶00 进行灌水,具体灌水量见表1,其中CK为总灌水量。分别在胁迫前、胁迫期间每3 d取1次叶片样品,在复水第3天、第6天取1次叶片样品进行测定。
1.3 测定指标与方法
叶片光合参数:选择气候稳定的晴天,于上午10∶00进行观测,采用Li-6400便携式光合仪测定
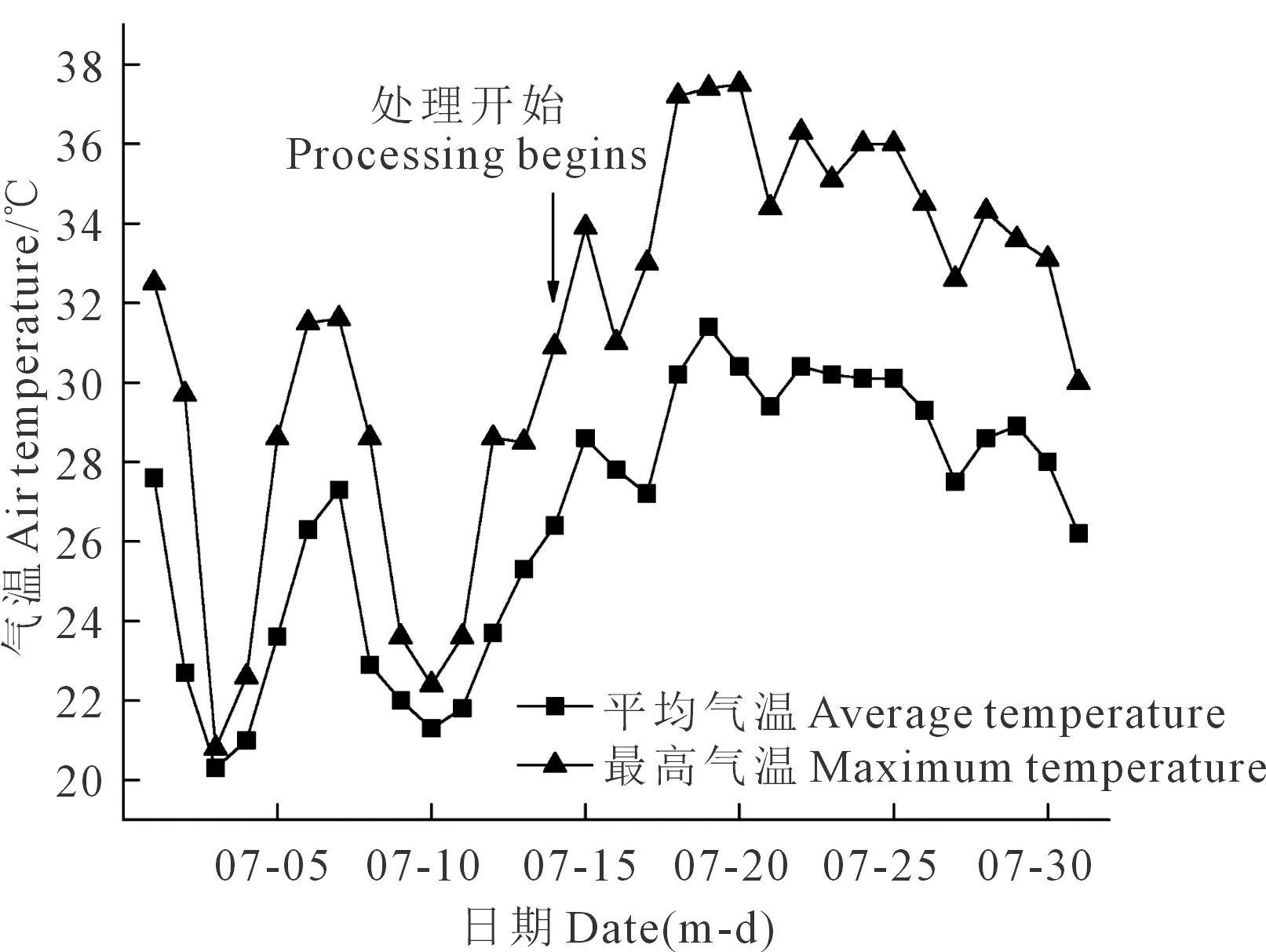
图1 2018年7月夏玉米苗期最高气温与平均气温变化Fig.1 Variation of maximum temperature and averagetemperature in summer maize seedling in July 2018
光合参数,主要包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)。
胁迫生理指标:脯氨酸(Pro)含量使用磺基水杨酸法测定[24];丙二醛(MDA)含量使用硫代巴比妥酸法测定[25];超氧化物歧化酶(SOD)活性使用NBT光还原法测定[26];过氧化物酶(POD)活性采用愈创木酚比色法测定[27]。
采用SPSS 22、Microsoft Excel 2007和DPS 7.05进行数据处理分析,并且采用Origin 85进行绘图。
2 结果与分析
2.1 干旱胁迫-复水对夏玉米Pro含量的影响
如图2所示,相较于CK处理,所有干旱处理组叶片中Pro含量均增高,在胁迫第3天达到峰值,W1和W2相较于CK分别提高32.7%和44.1%,且差异显著。随着胁迫时长增加胁迫的程度加剧,Pro含量逐渐减少,均在胁迫周期结束降低至CK水平。复水后,胁迫7 d和胁迫10 d的处理更有利于夏玉米Pro的累积,T1处理组在复水后呈先增后减趋势,第3天累积含量最高;T2处理组在复水后呈增长趋势,第6天累积量最高;而T3处理组在复水后的累积量无明显变化。说明10 d胁迫时长更有利于刺激复水后的夏玉米提高渗透调节能力。
2.2 干旱胁迫-复水对夏玉米SOD和POD活性的影响
SOD是作物抗氧化系统抵御伤害的第一道防线,将胁迫产生的超氧阴离子反应生成O2和H2O2。如图3所示,胁迫期间,所有处理的SOD活性均呈先增后减趋势,在胁迫3 d达到峰值,W1和W2相较于CK分别提高22.0%、12.4%,且差异显著。随着胁迫时长增加,胁迫程度加剧,T1处理组在第7天SOD活性降为最小值;T2处理组在第10天SOD活性降为最小值;而T3处理组在第10天SOD活性最小,到胁迫13 d便开始回升。复水后,SOD活性回升,T1处理组在第6天恢复至CK处理的活性水平;T2处理组在第6天W1和W2的SOD活性分别是CK处理的1.21倍和1.62倍;T3处理组在复水3 d活
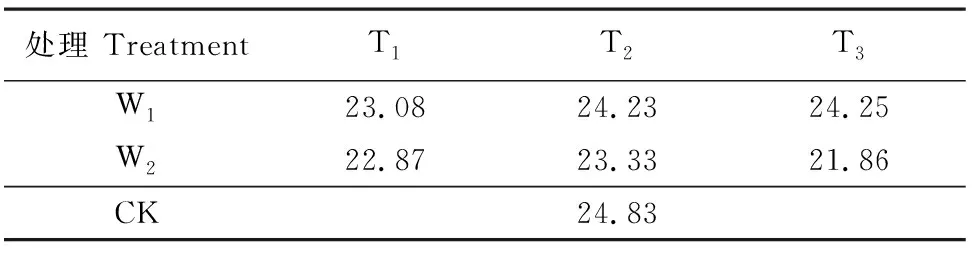
表1 夏玉米苗期灌水量统计表/kg
性最高,W1和W2分别是CK处理的1.47倍和1.88倍,之后呈下降趋势。长时间的水分胁迫加剧作物受到的胁迫程度,复水后SOD活性的升高也体现出作物的补偿效应。
POD 可以将SOD 产生的H2O2还原成为H2O,清除细胞内H2O2。由图4可见,叶片POD活性在轻度胁迫下有一定上升,在第3天相较于CK提高了28.5%,3 d之后便呈下降趋势。而W2的POD活性呈下降趋势,且低于CK水平。各处理下的POD活性均在胁迫周期结束降低至最小值;且胁迫时间越长,胁迫程度越大,POD酶的活性越低。复水后,POD活性均有回升,除W1T1处理下的玉米有显著的升高,在复水第3天达到567.60 ug·g-1·min-1之外,其他处理均基本恢复至与CK处理相同的水平。
2.3 干旱胁迫-复水对夏玉米MDA含量的影响
MDA是植物细胞膜酯化最重要的产物之一,其大小可用来反映植物膜系统的受损程度。由图5可得知,胁迫0~3 d,MDA含量基本不变,从3 d之后随着胁迫时长和胁迫程度的加大,MDA累积量逐渐增多,3个试验组均于胁迫最后1 d达到峰值,此时T1处理组的W1和W2处理分别为CK处理的1.5倍和2.02倍(图5A),T2处理组的W1和W2处理分别为CK处理的1.47倍和2.2倍(图5B),T3处理组的W1和W2处理分别为CK处理的2.4倍和3.81倍(图5C),且均达显著性差异。复水后MDA含量迅速降低,逐渐恢复至与CK处理相同的水平。水分胁迫处理下,W2的MDA含量高于W1,且胁迫时间越长,MDA含量越高,细胞膜酯化程度更为严重。

注:BS:胁迫前;S3:胁迫3 d;S7:胁迫7 d;S10:胁迫10 d;S13:胁迫13 d;R3:复水3 d;R6:复水6 d。下同。Note: BS: Before stress; S3: Stress for 3 days; S7: Stress for 7 days; S10: Stress for 10 days; S13: Stress for 13 days; R3: Rehydration for 3 dasy; R6: Rehydration for 6 days. The same below.图2 干旱胁迫-复水对夏玉米苗期脯氨酸(Pro)含量的影响Fig.2 Effects of drought stress and rewatering on proline content of maize in seedling stage
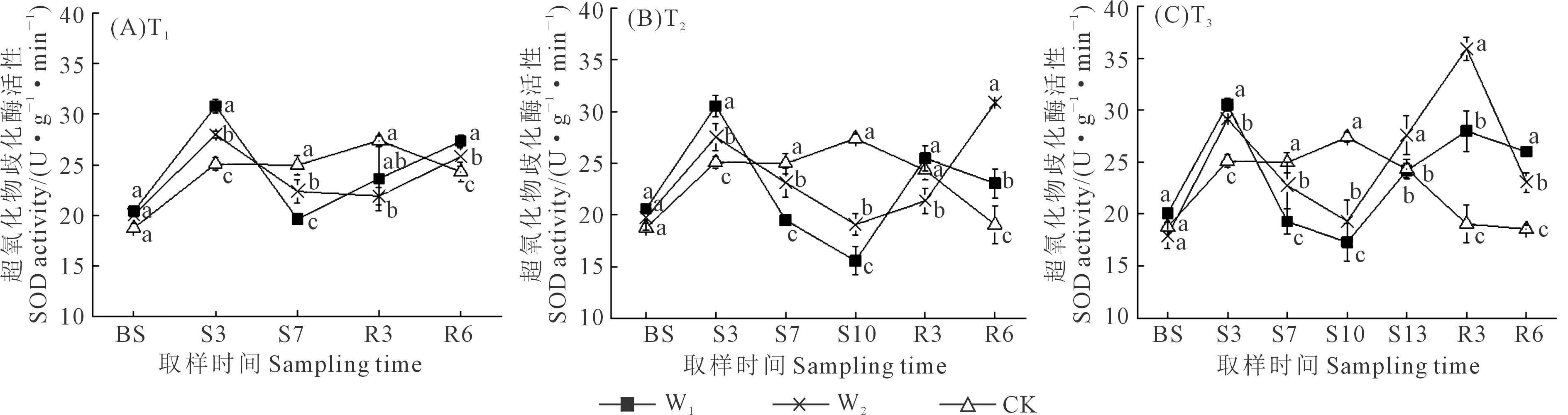
图3 干旱胁迫-复水对夏玉米苗期超氧化物歧化酶(SOD)活性的影响Fig.3 Effects of drought stress and rewatering on SOD activities of maize in seedling stage

图4 干旱胁迫-复水对夏玉米苗期过氧化物酶(POD)活性的影响Fig.4 Effects of drought stress and rewatering on POD activities of maize in seedling stage
2.4 干旱胁迫-复水对夏玉米苗期叶片光合参数的影响
由图6可见,干旱胁迫期间,夏玉米在不同胁迫水平下的Pn、Gs、Tr大小均表现为:CK >W1>W2,复水后各处理各参数值均高于其胁迫期间,且存在光合补偿效应,Pn表现为:W2>W1>CK。复水后T1处理组Pn、Gs、Tr均表现为W2>W1>CK,W1、W2显著高于CK;T2处理组除Pn的W2外,其余指标在处理间无显著性差异;而T3处理组各指标大小均表现为W2最高,且显著高于W1及CK处理。由此可知,经过中度水分胁迫锻炼的夏玉米复水后的光合补偿效应更显著,代谢能力更强。
2.5 夏玉米苗期抗性生理指标及光合参数的相关性分析
由表2可知,不同的干旱-复水处理下,Pro含量与SOD、POD活性呈极显著正相关,POD活性与MDA含量呈极显著负相关;SOD活性与Pn、Gs和Tr呈极显著正相关,MDA和Pro含量与Pn、Gs和Tr呈极显著负相关,POD活性与净光合速率、气孔导度和蒸腾速率无显著相关关系;净光合速率、气孔导度和蒸腾速率3个光合指标之间均呈现极显著正相关。

图5 干旱胁迫-复水对夏玉米苗期丙二醛(MDA)含量的影响Fig.5 Effects of drought stress and rewatering on MDA content of maize in seedling stage
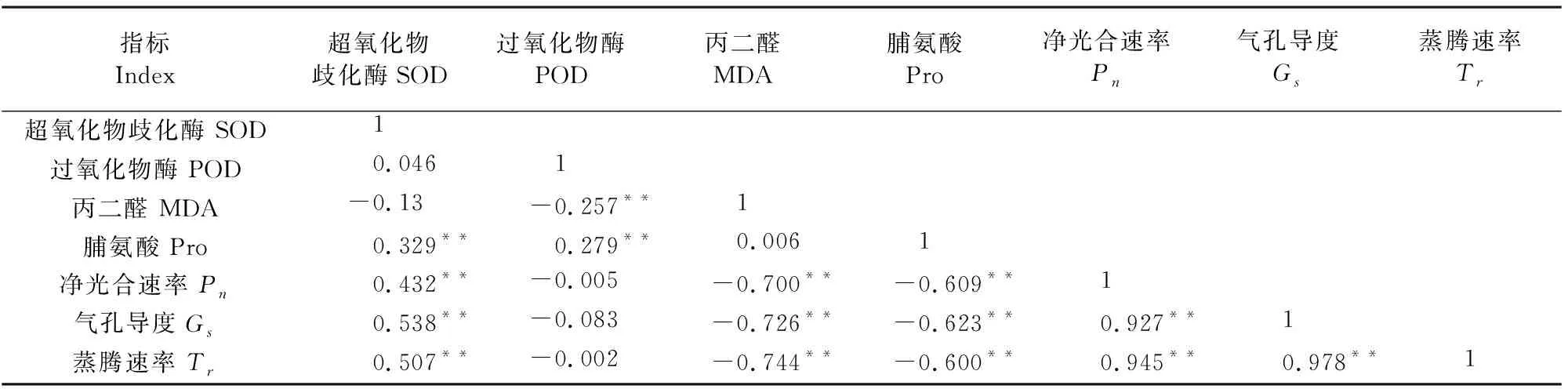
表2 夏玉米苗期抗氧化系统、脯氨酸以及光合参数的相关关系
3 讨 论
本试验结果表明,夏玉米苗期受到不同水平、不同时长的干旱胁迫时,植株体内脯氨酸(Pro)含量在胁迫期间先增大后减小,且W2>W1> CK;随着胁迫时长增加,胁迫程度加剧,Pro含量降低明显,胁迫13 d的累积量显著低于胁迫10 d和胁迫7 d的累积量。说明在胁迫初期,水分亏缺可诱导植株提高体内渗透调节能力,避免对自身产生伤害,但随胁迫程度的加剧,植物自身的渗透调节能力随之降低;复水后,前期遭受伤害的植株进一步提高渗透调节能力,维持系统与胁迫伤害的平衡,产生补偿累积效应。本试验胁迫10 d复水后的脯氨酸含量呈上升趋势,可能此时已达峰值,后期还是会回落。T1胁迫时间短,因此峰值出现在复水第3天,T2峰值可能出现在复水第6天。由此可见,高强度的水分胁迫尤其是长时间胁迫后,夏玉米的补偿效应会减弱,甚至造成伤害。
超氧化物歧化酶(SOD)活性在胁迫期间呈先增大后减小的趋势,在胁迫最后一天表现为CK>W2>W1,复水之后有明显回升,且胁迫时间越长,复水后的SOD活性越高。刘佳等[28]对辣椒的研究中,随着水分胁迫时间的延长,SOD活性也呈先增大后减小趋势,而郝舒雪等[29]对番茄幼苗的研究中发现SOD活性在水分胁迫下呈持续升高趋势。过氧化物酶(POD)活性在轻度胁迫下0~3 d有所提高,但在7 d 之后呈下降趋势,而遭受中度胁迫的植株POD活性持续呈下降趋势,复水后虽有回升,但也仅达到与CK基本相同的水平。丙二醛(MDA)在胁迫期间呈增大趋势,且W2>W1> CK,复水后迅速下降。杜彩艳等[30]在夏玉米苗期的研究中也得到相同结论。对比SOD和POD与MDA的变化趋势,本研究发现,SOD、POD(W1)从胁迫前到胁迫3 d呈增大趋势,有效抑制胁迫伤害(MDA基本保持不变);随着胁迫程度加剧,植物体内保护酶系统活力和平衡遭到破坏,MDA含量增加,酶活性和Pro含量开始降低,复水后MDA含量回到正常水平。由此也可进一步证明上述结论。
结合对比气温变化和SOD、POD活性以及Pro、MDA含量变化, CK处理下Pro、MDA含量基本保持不变,而SOD活性表现为先增大后减小的趋势,POD活性表现为持续减小趋势。施春晖等[31]在对猕猴桃的研究中表明,SOD活性对温度比较敏感,且与温度呈正相关;方芳等[32]对夏玉米幼苗的研究表明,在高温胁迫下,SOD 活性存在先上升后下降的现象,而POD活性在高温处理中呈下降趋势。本试验发现,SOD在T3试验组中胁迫第10天后活性开始增大,且基本达到与CK处理相同的水平;推测是因为胁迫第10天到胁迫第13天之间(7月24日至7月27日)气温降低,最高气温由36℃降低至32.6℃,平均气温由30.1℃降低至27.5℃。在水分胁迫和温度的双重影响下,气温降低减少温度对作物的胁迫,减缓对SOD活性的抑制,故SOD活性有增高趋势,复水后SOD的高活性仍在修复细胞膜所受到的损伤。
本试验发现,水分胁迫下,夏玉米叶片的Pn、Gs、Tr均小于CK,且W1>W2。其原因是水分胁迫影响气孔的开张度,进而影响细胞内外的气体交换,导致进入细胞的CO2受阻,光合原料不足,光合能力下降。毕建杰等[21]也得出叶片净光合速率随着叶片含水量的下降而减小的结论。在复水过后,叶片的Pn、Gs、Tr均高于胁迫期间,且W2>W1,可见中度胁迫处理后夏玉米光合的补偿作用相较于轻度胁迫处理更加明显。
由表2可看出,SOD活性和Pro含量呈极显著正相关,且在胁迫13 d的处理下,Pro的累积含量无明显变化,SOD活性出现上升趋势。由此可猜测在胁迫10 d夏玉米能承受的干旱胁迫达到阈值,可对此作进一步研究,探索夏玉米生理调节响应机制所对应的干旱阈值。
4 结 论
在苗期进行水分胁迫处理,夏玉米体内Pro含量呈先增大后减少的趋势,复水后回升,存在累积效应,胁迫10 d的累积量最大,且中度胁迫的累积量大于轻度胁迫;SOD在胁迫期间呈先增大后减小趋势,复水后有轻微增长,而且T1处理的SOD活性在复水6 d仍有明显升高;POD在W1处理下呈先增大后减小趋势,在第3天达到峰值,而W2处理下POD活性呈减少趋势;MDA在胁迫期间呈上升趋势,胁迫时长越长,MDA含量越高,复水后含量下降至正常水平。通过MDA的变化可以了解植物的受损情况,也能从侧面验证植物的渗透调节与抗氧化系统的调控机制。结合夏玉米的光合参数,中度胁迫(W1)处理下试验组在复水后Pn、Gs、Tr均高于W1组和CK对照组。故在灌溉水量受到限制的条件下,从植物生理的角度来看,维持10 d土壤50%~60%田间持水量的灌水量较利于作物生长,作物在复水后的光合补偿效果更加显著,有利于实现农业高效用水。
致谢:感谢徐家屯、李亮、丁怡博等在试验中以及论文写作中给予我的支持与帮助。
猜你喜欢
作物学报(2022年8期)2022-05-29
今日农业(2021年9期)2021-11-26
今日农业(2021年9期)2021-11-26
——以2020年为例
大麦与谷类科学(2021年5期)2021-11-09
古今农业(2021年2期)2021-08-14
当代水产(2021年6期)2021-08-13
今日农业(2020年20期)2020-12-15
今日农业(2020年17期)2020-12-15
今日农业(2020年23期)2020-12-15
今日农业(2020年23期)2020-12-15
