
耕作措施对玉米田耕层土壤微生物群落功能多样性的影响
2020-11-21 09:44陆思旭贺丽燕王旭东
干旱地区农业研究 2020年5期
陆思旭,贺丽燕,王旭东,2
(1.西北农林科技大学资源环境学院,陕西 杨凌 712100;2.农业部西北植物营养与农业环境重点实验室,陕西 杨凌 712100)
土壤微生物作为生态系统中最重要的活跃组分[1-2],参与土壤中的生化反应,是衡量土壤质量和作物生产力的重要指标之一[3-4]。丰富的微生物群落能够维持土壤生态系统的稳定,但土壤微生物易受到土壤环境的影响,尤其是耕作措施的改变在调控土壤理化性质的同时,常会对土壤微生物多样性产生不同的影响,这主要是由群落代谢功能的差异引起的。因此,耕作方式对土壤微生物功能多样性的影响也越来越受到国内外学者们的关注。
渭北旱塬位于黄土高原南部台塬和残垣沟壑区,该地区长期以来的耕作以传统翻耕为主,不仅加速土壤有机质的矿化,造成土壤肥力降低,而且也容易引起水蚀和风蚀,不利于土壤水分的保持[5]和作物的生长。为了降低长期翻耕带来的不利因素,免耕、深松和秸秆还田等保护性耕作在该地区逐渐推广应用,并且在蓄水保墒、培肥地力、作物增产方面已产生了较好的效果[5-6],土壤耕作在改变土壤结构的同时常会对土壤微生物群落产生影响,因此研究保护性耕作下土壤微生物多样性的变化,对渭北旱区农田土壤的可持续发展具有重要意义。目前国内外针对耕作措施对微生物多样性的影响已有大量研究,黄国勤[7]、杨江山[8]和Lynne[9]等学者研究发现,免耕、少耕等措施有利于土壤大团聚体的形成,秸秆覆盖在地表既可以为土壤微生物提供能源又减少了土壤水分的蒸发,为微生物的生长繁殖创造了良好的生境条件,并可在一定程度上增加土壤表层微生物量及大量的根系分泌物,增加土壤微生物群落的丰富度和多样性,增强土壤微生物群落的稳定性[10]。张志政[11]、何润兵[12]和李景[13]等通过研究发现深松可以降低土壤容重,增强土壤通透性,改变土壤的物理结构,引起土壤水、肥、气、热等变化,进而影响土壤微生物学特性,增加微生物数量和活性。而传统的翻耕措施对土壤翻动频率高、强度大,破坏了土壤结构,减少土壤大团聚体数量,加速了土壤呼吸作用和土壤水分的蒸发,降低土壤有机质含量[14],导致微生物活性降低。
不同耕作影响到土壤有机质含量的高低,而有机质和微生物方面存在着相互影响。因此,本文以渭北旱塬玉米田长期不同耕作定位试验为研究对象,运用Biolog-ECO微平板方法研究分析了不同耕作模式下(单一耕作模式如翻耕、免耕、深松;轮耕模式如免耕/深松、深松/翻耕、翻耕/免耕)土壤微生物功能多样性的变化,同时探讨了微生物碳源代谢多样性与土壤有机碳及其活性组分之间的相关关系,揭示不同耕作方式对土壤微生物多样性的影响,以期从微生物角度为旱作农田耕作模式的优化以及构建合理的耕层土壤结构提供理论依据。
1 材料与方法
1.1 试验区概况
本定位试验位于渭北旱塬东部的陕西省合阳县甘井镇(106°04′E ,35°19′N)。试验区属典型的黄土残塬沟壑半湿润易旱区,年均气温11.5℃,年降水量500~600 mm,降雨年际和季节变异性较大,主要集中在7—9月,是典型的黄土高原旱作雨养农业区。试验地地势平坦,全年无灌溉,土壤类型为黑垆土,属于中壤土,黄土母质疏松、深厚并含有丰富的矿质养分,蓄水保肥能力强。试验前0~10 cm土层土壤理化性质为:土壤pH值7.9,容重1.38 g·cm-3,有机质7.1 g·kg-1,全氮0.74 g·kg-1,碱解氮40.62 mg·kg-1,速效磷3.77 mg·kg-1。
1.2 试验设计
本试验于2007—2018年进行长期定位试验,采用春玉米连作一年一熟制的种植制度。试验采用随机区组设计,在作物收获后秸秆全部粉碎还田基础上实施6种耕作处理:连年翻耕(C/C)、连年深松(S/S)、连年免耕(N/N)、免耕/深松轮耕(N/S)、深松/翻耕轮耕(S/C)和翻耕/免耕轮耕(C/N)。其中,翻耕(C)处理为采用传统铧式犁(型号为1LF-230型双向反转二铧犁,耕宽60 cm,耕深20~30 cm)将土壤全面翻耕20~25 cm,秸秆翻埋于土壤中,地表疏松裸露度过休闲期;深松(S)处理采用深松机(型号为1S2200,耕宽50 cm,耕深30~40 cm)进行深松,深度为30~35 cm,秸秆覆盖地表度过休闲期;免耕(N)处理在玉米收获后将秸秆粉碎覆盖地表度过休闲期,不采取任何土壤耕作措施。每种耕作处理设3个重复,小区面积为112.5 m2(22.5 m×5 m),2007—2018年土壤耕作顺序见表1。
春玉米每年在4月下旬播种,9月下旬收获。供试品种为郑单958,播种量为75 kg·hm-2,种植密度为67 500株·hm-2。各耕作处理的施肥量相同。施肥量为:N 150 kg·hm-2、P2O5120 kg·hm-2、K2O 90 kg·hm-2,其中氮、磷、钾肥分别为尿素、磷酸二铵和氯化钾。50%氮肥、全部磷肥和钾肥在播前施用,剩余50%氮肥在大喇叭口期追施。其他田间管理措施同当地大田生产。

表1 2007—2018年土壤耕作次序
1.3 土壤样品的采集与分析
于2018年春玉米收获后,用土钻采集0~10 cm土层的样品,其中每个小区按“S”形采样法采集6个点,去除土壤中石砾和植物残茬等杂物现场混匀为1份土壤样品,3个重复小区采集3个重复样品。置于密封袋放入冰盒中带回实验室。土样带回实验室后分成两份:一份过2 mm筛后置于4℃冰箱保存,用于测定微生物功能多样性、微生物量碳和水溶性有机碳含量;另一份置于通风、干燥、阴凉的室内风干,将风干样过0.25 mm和1 mm筛用于测定有机碳和易氧化有机碳含量。
1.3.1 土壤因子测定 土壤微生物量碳用氯仿熏蒸浸提法[15]测定;水溶性碳用去离子水浸提法[16]测定;易氧化有机碳用333 mmol·L-1KMnO4氧化法[17]测定;土壤颗粒有机碳的提取采用六偏磷酸钠分散法[18]测定,土壤总有机碳用重铬酸钾-外加热法[19]测定。
1.3.2 土壤微生物群落功能多样性的测定 土壤微生物功能多样性采用Biolog-ECO平板法测定,具体操作步骤参照Classen[20]等的方法。称取相当于5.0 g风干土壤的新鲜土壤样品置于无菌三角瓶中,加入45 mL 0.85%灭菌的NaCl溶液,封口,震荡30 min(160 r·min-1),静置后去其上清液,在超净工作台上,采用十倍稀释法,用无菌NaCl溶液(0.85%)稀释至10-3,随后用8通道移液器将上述稀释液加入Biolog-ECO微平板(ECO Micro Plate, 美国Marix Technologies Corporation)上,每孔150 μL。将接种好的微平板放入25℃条件下培养,每24 h用Biolog微生物自动读数装置读取Biolog-ECO微平板在590 nm波长下的吸光值,连续测定10 d。
1.4 数据处理与统计分析
Biolog-ECO平板每孔的平均颜色变化率(average well color development,AWCD)是反映土壤中可培养微生物对不同底物(碳源)的利用能力,在一定程度上反映了土壤微生物的代谢活性,AWCD值越大,表示土壤微生物利用碳源的能力越强,代谢活性越高[21],计算公式如下:
AWCD=∑(Ci-R)/n
(1)
式中,Ci为各反应孔的吸光值;R为对照孔的吸光值;Ci-R<0的反应孔在计算中默认为零;n为碳源数目,为31。
土壤微生物群落丰富度指数(S)表征土壤微生物的丰富度,其值越高表明微生物群落越丰富,采用反应孔(吸光值≥0.25则代表该孔的碳源被利用)的数目表示。
Shannon-Wiener多样性指数(H)、均匀度指数(E)和Simpson指数(D)表征微生物多样性程度,计算公式如下:
Shannon-Wiener多样性指数(H):
H=-∑(Pi×lnPi)
(2)
式中,Pi表示第i孔相对吸光值与整板平均相对吸光值总和的比值,即:
Pi=(Ci-R)/∑(Ci-R)
(3)
均匀度指数(E):
E=H/lnS
(4)
Simpson指数(D):
D=1-∑(Pi)2
(5)
本试验采用培养168 h的数据进行土壤微生物功能多样性的计算.
试验数据采用Excel 2010进行初步处理分析,采用SPSS 19.0进行方差分析及差异显著性检验(LSD法)和主成分分析,Canoco 5.0软件进行冗余(RDA)分析,图形采用Origin 8.5、Canoco 5.0绘制。
2 结果与分析
2.1 不同耕作处理下土壤微生物群落的碳源代谢活性
6种耕作处理的土壤AWCD值均随着培养时间的延长呈现逐渐上升的趋势(图1)。0~24 h内各处理的AWCD值一直趋近于零,表明碳源在24 h之内基本未被利用,处于微生物适应期;培养24~168 h内,AWCD值呈现急速上升趋势,表明土壤微生物逐渐适应了Biolog微平板的基质环境,进入对数生长期,此时碳源被微生物快速消耗,微生物代谢活性较强;培养168 h后,各处理的AWCD值增长速度逐渐减缓,土壤微生物的生长趋于稳定。不同耕作处理间土壤微生物群落的AWCD值在培养48 h开始出现显著差异,N/S和N/N处理的AWCD值在48~96 h内接近,均显著高于S/S、S/C、C/N和C/C处理,在120 h时N/S处理的AWCD值达到最高,显著高于其他处理。在168 h处微生物进入生长稳定期,此时N/S处理的AWCD值较N/N、S/S、S/C、C/N和C/C处理分别高出9.51%、21.90%、29.89%、37.13%和54.54%(P<0.05)。同时,N/N、S/S、S/C和C/N处理的AWCD值也较C/C处理显著提高41.12%、26.78%、18.98%和12.70%。因此,少耕和免耕的耕作方式可以显著提高土壤微生物的碳源代谢活性,且N/S处理的效果最佳。
2.2 不同耕作处理下土壤微生物群落的功能多样性指数
不同的多样性指数反映了土壤微生物群落功能多样性的不同方面。Shannon-Wiener指数和均匀度指数越高表明微生物群落的多样性越高;Simpson指数越大,表示微生物群落内的优势种越突出。由表2可知不同耕作措施下的土壤微生物群落功能多样性指数存在一定差异。与AWCD值相似,N/S、N/N可以提高土壤的丰富度指数(S)、Shannon-Wiener指数(H)和Simpson指数(D),且均与C/C处理达到显著差异(P<0.05),而均匀度指数(E)较C/C处理有所降低(P<0.05)。表明N/S和N/N处理的土壤微生物种群丰富,优势种群突出,对碳源的利用程度较高,但降低了群落的均匀度指数。
2.3 不同耕作处理下土壤微生物的碳源利用特征
根据Biolog-ECO微平板上碳源官能团的不同将31种碳源划分为6类,包括糖类(10种)、氨基酸类(6种)、羧酸类(7种)、多聚物类(4种)、多酚化合物类(2种)和多胺类(2种)[22]。本试验对168 h处的吸光值进行土壤微生物碳源利用特征的计算,结果表明(图2):各处理土壤微生物对6类碳源的利用程度依次为:氨基酸类>糖类>羧酸类>多聚物类>多胺类>多酚化合物类,可见氨基酸类、糖类和羧酸类物质是该地区土壤微生物利用的主要碳源。其中N/S、N/N和S/S处理土壤微生物对6类碳源的利用能力均大于C/C处理。与C/C处理相比,S/C和S/S处理显著提高了对糖类、多聚物类和多胺类碳源的利用能力;N/N处理显著提高了对糖类、羧酸类、多聚物类和多胺类碳源的利用能力;C/N处理显著提高了对多胺类碳源的利用能力;N/S处理显著提高了对各类碳源的利用能力。同时,N/S处理对氨基酸类、多聚物类、多酚化合物类和多胺类碳源的利用能力也显著高于N/N、S/C和C/N处理。

注:图中不同小写字母表示同一时间点不同耕作处理间差异显著(P<0.05)。Note:Different lowercase letters in the figure indicate that there are significant differences (P<0.05) between different tillage treatments at the same time point.图1 不同耕作处理下土壤微生物群落的平均颜色变化率Fig.1 AWCD of the microbial community in different treatments

表2 不同耕作处理的土壤微生物功能多样性指数
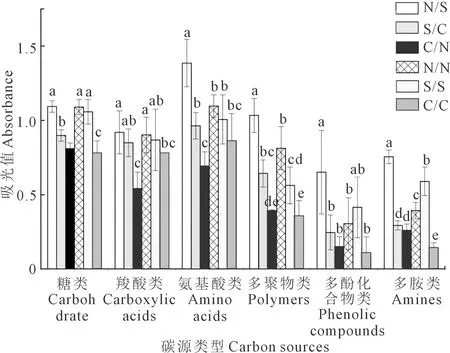
注:不同小写字母表示同一碳源在不同耕作处理间差异显著(P<0.05)。Note: Different lowercase letters indicate significant differences among tillage treatments in the same carbon sources (P<0.05).图2 不同耕作处理下土壤微生物对六类碳源的利用状况Fig.2 The utilization of six kinds of carbon sourcesin different treatments
2.4 土壤微生物碳源利用的主成分分析
采用主成分分析来研究不同耕作处理下土壤微生物群落对31种碳源的利用情况。根据主成分个数的提取原则(相对应特征值大于1的主成分)共提取了前10个主成分,累积方差贡献率达到90.71%,可以解释大部分原变量特征。其中第一主成分(PC1)的贡献率为27.34%,权重最大;第二主成分(PC2)次之,贡献率分别为12.71%,第3~10主成分的贡献率较小,分别为3.65%~10.57%,因此本文选取前2个主成分来表征微生物群落的碳源代谢特征。
主成分分析还表明不同耕作处理的土壤微生物群落在PC轴上有明显的分布差异(图3)。在PC1轴上,S/S、N/S和N/N处理分布在正方向,C/C、C/N和S/C处理分布在负方向。在PC2轴上,N/N、N/S、S/C和C/C处理主要分布在正方向,C/N和S/S处理主要分布在负方向。进一步进行方差分析(表3)可知不同耕作处理在PC1和PC2上的得分系数出现显著差异。在PC1轴上,N/S、N/N、S/S、S/C处理与C/C处理的分布差异显著,表明碳源利用方式不同。在PC2轴上,N/S、S/C、C/N、N/N处理与C/C处理的分布差异不显著,表明碳源利用方式相近。
2.5 土壤微生物碳源利用与环境因子的冗余分析
选取土壤总有机碳(SOC)及其活性组分指标(易氧化有机碳EOC、水溶性有机碳WSOC、微生物量碳MBC、颗粒有机碳POC)作为环境因子变量与土壤微生物碳源利用特征进行冗余分析(RDA)。表4是不同耕作措施下土壤有机碳及活性组分指标,基本表现为免耕、深松以及两者相结合轮耕模式下的有机碳及其活性组分的含量高于传统翻耕措施,其中N/S处理下的EOC、MBC和POC与C/C处理相比均达到显著性差异。

注:同一耕作处理的3个点代表3个重复。Note: Three points of the same tillage treatment represent three duplicates.图3 不同耕作处理下土壤微生物群落主成分分析Fig.3 Principal component analysis of soil microbialcommunity in different treatments

表3 不同耕作处理的主成分得分系数
冗余分析结果显示Axis1和Axis2的累积解释率达到65.69%,特征根分别为0.611和0.046,相关系数分别为0.915和0.815,两排序轴对应的环境变量对于响应变量的解释贡献均达到统计学上的显著水平。如图4所示,Axis1与SOC、EOC、WSOC、MBC和POC均呈正相关,Axis2与WSOC、POC和MBC呈正相关,与SOC和EOC呈负相关。5个碳指标的箭头连线均较长,且SOC和EOC与Axis1轴的夹角较小,表明土壤微生物碳源代谢与这两个碳指标的相关性较大,其中SOC、EOC对碳源利用类型具有显著影响。MBC和POC与羧酸类碳源的相关性较大,SOC、EOC与糖类、氨基酸类和多酚化合物类碳源的相关性较大,但WSOC与各类碳源的夹角相对较大,相关度较低。

表4 不同耕作措施对土壤有机碳及其活性组分的影响

注:实线代表土壤微生物对6类碳源的利用,虚线代表环境因子(土壤有机碳及其组分指标)。SOC:土壤有机碳;EOC:易氧化有机碳;WSOC:水溶性有机碳;MBC:微生物量碳;POC:颗粒有机碳。Note:The solid line represents the utilization of 6 types of carbon sources by soil microorganisms, and the dotted line represents environmental factors (soil organic carbon and its fractions). SOC: Soil organic carbon; EOC: Easily oxidizing organic carbon; WSOC: Water soluble organic carbon; MBC: Microbial biomass carbon; POC: Particulate organic carbon.图4 土壤微生物功能多样性的冗余分析Fig.4 Redundancy analysis on the functional diversityindexes of soil microbial community
3 讨 论
微生物在农田生态系统中起重要作用,其结构和功能的多样性在一定程度上反映了农田生态系统的基本状况[4]。本研究发现,免耕和少耕等轮耕的耕作方式较传统翻耕显著提高了土壤微生物的碳源代谢活性(图1),碳源代谢活性较高的土壤也具有较高的微生物多样性指数,N/S和N/N处理的Shannon-Wiener指数、丰富度指数和Simpson指数均较高(表2)。这主要与土壤扰动程度引起的土壤环境变化以及秸秆的还田量有关。有研究表明[23],土壤耕作对土壤含水量、容重、pH值和土壤全氮等有显著影响,土壤理化性质等的变化影响着微生物生存的微环境,进而影响土壤微生物群落的种类、活性和分布的关键因素。C/C处理长期频繁地扰动土壤,加快了表层土壤有机质的矿化[24],导致大团聚体比例降低[25],同时会引起水分的大量损失,破坏了微生物生存的土壤环境,使微生物的代谢活性降低。免耕和深松措施较翻耕减少了对土壤的扰动,改善了土壤结构和理化性质,有效降低耕作层的土壤容重,增加毛管孔隙的比例,使土壤能够容纳更多的水分,进而提高了微生物的代谢活性[26]。N/S处理的轮耕模式既避免了对耕层土壤过度扰动,也对土壤的通气以及透水性等理化性状和生物性状起到调控作用[27],更大程度地保持了土壤原有的结构体,有利于土壤微生物代谢活性的维持,且N/S的秸秆还田量(14 433 kg·hm-2)高于其他处理(12 636~14 205 kg·hm-2),大量秸秆覆盖于地表,为土壤微生物提供了丰富的碳源和物质来源,因此微生物的碳源代谢活性达到最高。这与姬艳艳[28]和邵泱峰[29]等研究结果相似。但也有研究认为,过高的秸秆还田量(13 000 kg·hm-2)会对土壤微生物碳源代谢活性产生不利影响[30],这与试验地的土壤类型、土壤肥力、气候条件等有关,东北地区黑土有机质含量高,秸秆还田量过高会导致土壤碳、氮比例失调,从而抑制了微生物代谢活性,而在本试验西北半湿润易旱区,黑垆土有机质含量低,接近全量秸秆还田后,可以增加土壤有机碳、活性有机碳和全氮含量,继而增加微生物活性。土壤微生物多样性的变化也通常受到土壤理化因子等环境因素的驱动[31-33],前人研究结果显示[5-6],西北黑垆土区秸秆还田后的少耕和免耕措施可以有效增加土壤有机碳含量,改善土壤理化性质,减少土壤中水分的蒸发,使植物生长代谢加快、根系分泌物增多,因此会促进土壤微生物的生长,增加其多样性。土壤微生物量与微生物群落结构具有显著相关性[34]。一些研究也表明,免耕土壤的微生物Shannon-Wiener指数、丰富度指数和Simpson指数都显著高于传统耕作[35-36]。如Balota等[37]研究认为耕作强度的降低会增加土壤微生物多样性。张志政等[11]的研究也发现,深松改善了耕层土壤物理结构,使土壤的微生物Shannon-Wiener指数、丰富度指数和Simpson指数都显著提高。所以在秸秆还田下,少耕、免耕等耕作措施比传统翻耕更有利于维持土壤微生物的活性及多样性。
微生物对不同类型碳源的相对利用程度可以反映微生物的代谢功能类群[38]。主成分分析结果显示,在秸秆还田基础上的6种耕作措施下土壤微生物利用的主要碳源为氨基酸类、糖类和羧酸类物质(图2),可能是因为秸秆还田后显著增加了氨基酸和单糖等物质的含量[39],进而促进了以偏好氨基酸类、单糖类物质为碳源的微生物群落的生长发育,这与李春格[40]和邵丽[41]等的研究结果一致。根据已有研究认为玉米根系分泌物组分有利于土壤中偏好羧酸类碳源的微生物种群增长[38,42]。不同耕作处理在PC轴坐标上的分布存在显著差异与在该轴上的碳源利用能力是相互对应的[43]。其中N/S对氨基酸类、糖类和羧酸类碳源利用能力的显著提高是导致其代谢活性最强的原因之一,而N/N仅显著提高了对糖类和羧酸类碳源利用能力,S/S和S/C仅显著提高了对糖类碳源的利用能力,这也是导致其碳源代谢活性较强的原因之一。充分说明氨基酸、糖类和羧酸这3类碳源是导致微生物代谢呈现差异的敏感碳源,这与吴迪等[44]的研究结果一致。冗余分析结果表明,耕作方式对土壤微生物群落结构产生了影响;SOC和EOC主要通过影响土壤微生物对糖类、多酚化合物类和氨基酸类碳源的利用来影响土壤微生物的碳源代谢特征,而土壤有机碳是影响微生物群落结构变化的关键因素[45],所以以糖类、多酚化合物类和氨基酸类碳源为主要能源物质的微生物较活跃。这与Li等[46]的研究结果一致,即土壤有机碳是引起微生物碳源代谢发生分异的重要因素。Chen等[34]研究发现,总有机碳和微生物生物量碳是影响土壤微生物群落结构的主要因素,土壤有机碳能够为土壤微生物提供丰富的碳源,影响土壤微生物群落结构。本研究结果与其相一致,也表明微生物碳源代谢与SOC的相关性较好,在一定条件下,土壤中较高的有机碳含量与较高的微生物活性和生物量是一致的。
4 结 论
秸秆还田基础上,免耕、少耕的土壤微生物碳源代谢活性显著高于翻耕,且相应的微生物多样性指数(丰富度指数、Shannon-Wiener指数和Simpson指数)也显著高于翻耕处理,其中N/S处理的土壤微生物代谢活性和多样性指数均较高。土壤微生物对碳源利用的偏好因耕作方式不同有所差异,其中氨基酸类、糖类和羧酸类物质是微生物主要利用的碳源,SOC和MBC主要通过影响土壤微生物对糖类、氨基酸类和羧酸类碳源的利用来影响土壤微生物的碳源代谢,其中N/S轮耕处理对各类碳源的利用率都高于C/C处理。所以,N/S轮耕是适宜该地区的推荐耕作模式。
猜你喜欢
油气田地面工程(2022年9期)2022-10-26
农业技术与装备(2022年3期)2022-05-31
山东农机化(2022年1期)2022-03-05
腐植酸(2021年2期)2021-12-04
中国土壤与肥料(2021年5期)2021-12-02
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
当代水产(2020年11期)2021-01-04
红领巾·探索(2018年12期)2018-01-26
家庭用药(2018年11期)2018-01-23
