
接穗对油茶砧木根生理特性及解剖结构的影响
2020-11-20 00:53郑芳奕杨志坚廖鹏辉黄蓝明冯金玲
森林与环境学报 2020年6期
郑芳奕,杨志坚,陈 辉,廖鹏辉,黄蓝明,冯金玲
(1.福建农林大学林学院,福建 福州 350002;2.福建省林业科学技术推广总站,福建 福州 350003)
接穗与砧木在嫁接后形成一个完整共生体,彼此间产生竞争与共生关系[1]。砧木决定植物的水分和养分吸收,接穗为砧木提供所需的光合同化物,而养分的吸收、积累和分配取决于两者的耦合反应,影响着嫁接植物的生长发育[2-4]。砧穗间存在一定的互作,选择不同砧木与相同接穗或者不同接穗与相同砧木嫁接后,植物的生长发育特性均表现出差异性[5-6]。目前,嫁接砧穗互作研究主要集中在嫁接亲和性和砧木对接穗的影响等方面,而接穗对砧木影响的研究还相对较少。根是植物重要的营养器官,只有通过根的不断生长,植物才能达到吸收水分和无机营养物质的目的[7]。接穗不仅影响砧木根的生长,而且影响其生理生化代谢,导致嫁接体的移栽成活率和抗逆性等不同[8-9]。不同接穗既然对相同砧木在客观上存在不同影响,就得认识接穗与砧木亲缘关系对砧木生长、生理代谢等方面的影响规律,从而制定合理的栽培管理措施使砧木根系与接穗代谢协调。根尖作为实现吸收功能的主要部位,是根系中生命活动最旺盛的区域[10]。虽然关于接穗对砧木根系差异的报道有一些,但直接针对接穗诱导砧木根尖的研究并不多,尤其缺乏其对根尖结构影响的报道。
油茶(CamelliaoleiferaAbel.),又名茶子树、茶油树等,是山茶科(Theaceae)山茶属(Camellia)的多年生常绿小乔木或灌木[11]。山茶科植物全球共28属约700种,中国目前有15属约 400种,其中60余种可作为木本油料树种,其中,普通油茶种植面积最大,分布区域最广,适应性也最强[12]。油茶作为我国特有的木本油料树种[3],不仅寿命长、适应性强,而且还能保持水土、绿化荒山,具有良好的社会、经济和生态效益[13]。芽苗砧嫁接苗是油茶生产上的主要苗木来源,鉴于油茶芽苗砧嫁接苗的广阔应用前景和砧木根系对嫁接苗生长的重要性,开展不同接穗对芽苗砧嫁接体的根系影响试验,通过测定根系生长、生理生化指标,观察根尖解剖结构,分析与砧木不同亲缘关系的接穗对普通油茶砧木根的差异影响,从而探讨相同砧木具有不同栽培表现的原因,以期为油茶生产提供技术参考和理论基础。
1 材料与方法
1.1 材料
2016年11月,在福建省闽侯桐口林场华南工区,选取1棵生长良好,产量高的15年生普通油茶(闽48)植株,采集所有果实,阴干,自然开裂,脱壳,收集所有粒大、饱满、无病虫害的种子,贮藏于4 ℃冰箱。2017年1月,取出,培育芽苗砧,至5月下旬,起芽苗砧,着手嫁接。茶花(CamelliajaponicaL.)和普通油茶的接穗均为当年生、健壮、半木质化的新梢,茶花接穗采自福建农林大学校园内15年生的茶花树,普通油茶的接穗采自与砧木种子来源相同的母树。
1.2 试验设计
2017年5月下旬,在福建农林大学科技园田间试验地进行油茶芽苗砧嫁接,砧木均来自同一母树的普通油茶。接穗设3个处理:处理1为东方亮品种茶花的接穗(异源嫁接苗);处理2为普通油茶接穗(同源嫁接苗);处理3为无接穗(CK,自根苗)。3个处理的砧木均采用相同的处理方法。嫁接和处理后的植株移栽至大田,采用随机区组试验设计,每个处理3次重复,每个重复30株,共270株,采用相同田间管理。2018年8月,每个重复随机选10棵幼苗作样株,测定各项指标。
1.3 测定项目与方法
1.3.1 生长指标测定 2018年8月,用游标卡尺测每株的地径(mm)。取样株,数单株总主根数和侧根数,用直尺测单株最长主根长(cm)和侧根长(cm)。
1.3.2 根尖显微结构 取根尖,用标准固定液(formalin-acetic acid-alcohol,FAA)固定,经酒精脱水,二甲苯透明及石蜡包埋后切片,切片经1.0%的番红和苯胺蓝染色,显微镜下观察根系解剖结构并拍照。
1.3.3 生理指标测定 取0.2 g根,采用蒽酮比色法测定根的可溶性糖含量。称取0.5 g根,液氮速冻,研磨,加入4 mL磷酸钠缓冲液(0.05 mol·L-1,pH值为7.8),移至10 mL离心管中,离心10 min (4 ℃,6 000 r·min-1),取上清液,放入4 ℃冰箱保存,待测。采用考马斯亮蓝法测定可溶性蛋白质含量;采用紫外线吸收法测定过氧化氢酶(catalase,CAT)活性;采用邻苯二酚法测定多酚氧化酶(polyphenol oxidase,PPO)活性;采用愈创木酚法测定过氧化物酶(peroxidase,POD)活性;采用硫代巴比妥酸比色法测定丙二醛(malondialdehyde,MDA)含量[14-15]。
1.4 数据统计与分析
采用Excel 2010软件进行数据的整理与图表的绘制,采用SPSS 19.0软件进行差异性检验、多重比较(Duncan′s)和相关性分析。
2 结果与分析
2.1 接穗对砧木生长的影响
由图1可见,不同接穗处理对普通油茶砧木的地径、主根数、侧根数、主根长和侧根长均有显著影响。处理3的砧木地径最大,处理2的次之,处理1的最小;处理1的砧木主根数、侧根数最多,处理2的次之,处理3的最少;处理1的砧木侧根长最长,处理2的次之,处理3的最短;处理1的砧木主根长最长,处理3的次之,处理2的最短。表明不同接穗嫁接处理影响普通油茶砧木根系生长,嫁接促进砧木根生长,接穗与砧木间的亲缘关系越远,越有利于根系生长,但嫁接不利于砧木地径生长。

(a)地径Ground diameter (b)根数Number of roots (c)根长Root length
2.2 接穗对砧木根尖解剖结构的影响
由图2可见,不同接穗处理后普通油茶砧木根尖横切细胞发生明显变化。处理1,细胞排列紊乱,扁平,明显膨胀,细胞内细胞质淡稀[图2(a)]。处理2,横切面细胞排列规则,均匀,细胞饱满,皮层的最内2~3层细胞壁加厚,呈马蹄形[图2(b)]。处理3,细胞状态界于处理2和处理1之间,皮层的最内一层细胞胞壁加厚[图2(c)]。由图2、表 1可得出,不同接穗处理对普通油茶砧木根尖的皮层厚度、中柱鞘厚度、维管柱直径和根尖直径均有显著影响。处理2的砧木根尖中柱鞘厚度最厚,处理1的次之,处理3的最薄,说明嫁接提高中柱鞘的分生能力,促进根横向生长,接穗与砧木的亲缘关系越近,横向生长能力越强。处理2的砧木根尖皮层厚度最厚、根尖直径最大,处理1的次之,处理3的最小;处理3的砧木根尖维管柱直径最大,处理2的其次,处理1的最小,说明嫁接减弱了砧木根的吸收效率和运输能力,同源嫁接体的根吸收效率和横向运输能力弱于异源嫁接体,纵向运输能力则强于异源嫁接体。不同接穗处理普通油茶砧木根尖表皮厚度没有差异,说明其不受接穗影响。

(a)处理1 Treatment 1 (b)处理2 Treatment 2 (c)处理3 Treatment 3

表1 不同接穗处理普通油茶砧木根尖的显微结构指标
2.3 接穗对砧木根生理生化指标的影响
由图 3 可见,不同接穗处理对普通油茶砧木根的可溶性糖含量、可溶性蛋白含量、POD活性、CAT活性、PPO活性和MDA含量均有显著影响。处理3的砧木根可溶性糖含量和可溶性蛋白含量最多,处理2的次之,处理1的最少,说明嫁接降低根系营养物质贮藏,接穗与砧木的亲缘关系越远,越不利于营养物质贮藏。处理1的砧木根POD活性最大,处理2的次之,处理3的最小,说明嫁接提高了POD活性,且接穗和砧木亲缘关系越远,POD活性越大。处理3的砧木根CAT活性最大,处理1的次之,处理2的最小,说明嫁接降低了CAT活性,接穗和砧木的亲缘关系越远,CAT活性越大。处理2的砧木根PPO活性最大,处理3的次之,处理1的最小,说明接穗与砧木亲缘关系越远,PPO活性越小。处理1的砧木根MDA含量最多,处理3的次之,处理2的最少,说明接穗与砧木的亲缘关系的远近决定了MDA含量,关系越远,胁迫越强。

注:不同小写字母表示差异显著(P<0.05)。Note: different lowercase letters indicate significant difference (P<0.05).
2.4 各指标相关性分析
表2相关分析表明,砧木根的可溶性蛋白质含量(X11)与根尖表皮厚度(X1)呈显著正相关,与根尖维管柱直径(X5)和砧木地径(X10)均呈极显著正相关,与砧木的侧根长度(X7)、主根数(X8)和侧根数(X9)均呈显著负相关。砧木根的可溶性糖含量(X12)与根尖维管柱直径(X5)和砧木地径(X10)均呈极显著正相关,与砧木的侧根长度(X7)和主根数(X8)均呈显著负相关,与砧木的侧根数(X9)呈极显著负相关。砧木根的PPO活性(X13)与根尖中柱鞘厚度(X3)和根尖直径(X4)均呈极显著正相关。砧木根的POD活性(X14)与根尖维管柱直径(X5)呈显著负相关,与砧木的主根长度(X6)、侧根长度(X7)、主根数(X8)和侧根数(X9)呈极显著正相关,与砧木地径(X10)呈极显著负相关。砧木根的CAT活性(X15)与根尖皮层厚度(X2)呈显著负相关,与根尖中柱鞘厚度(X3)和根尖直径(X4)呈极显著负相关。砧木根的MDA含量(X16)与根尖中柱鞘厚度(X3)呈极显著负相关,与砧木的主根长度(X6)呈极显著正相关。
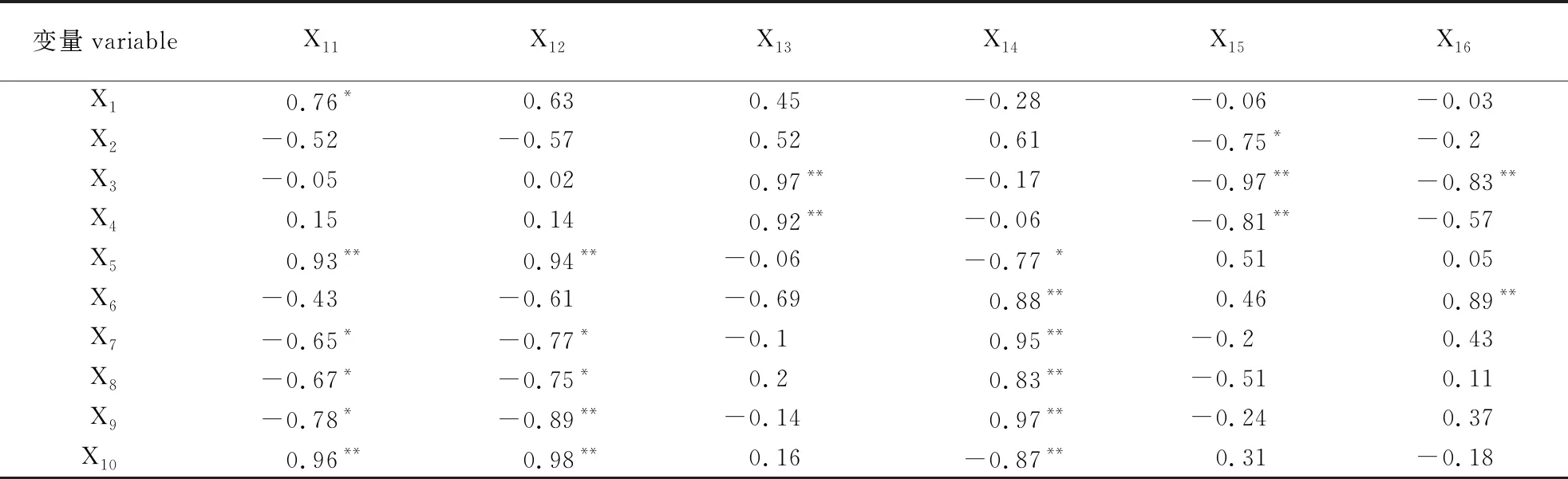
表2 接穗处理普通油茶砧木各指标间的相关分析
由表3可见,根尖的皮层厚度(X2)与中柱鞘厚度(X3)、根尖直径(X4)、砧木的侧根长度(X7)和侧根数(X9)均呈显著正相关,与根尖维管束直径(X5)和砧木地径(X10)均呈显著负相关,与砧木的主根数(X8)呈极显著正相关。根尖中柱鞘厚度(X3)与根尖直径(X4)呈极显著正相关。根尖维管束直径(X5)与砧木的侧根长度(X7)呈显著负相关,与砧木的主根数(X8)和侧根数(X9)均呈极显著负相关,与砧木地径(X10)呈极显著正相关。砧木的主根长度(X6)与侧根长度(X7)和侧根数(X9)均呈显著正相关。砧木的侧根长度(X7)与主根数(X8)和侧根数(X9)均呈极显著正相关,与砧木地径(X10)呈极显著负相关。砧木的主根数(X8)与侧根数(X9)呈极显著正相关,与砧木地径(X10)呈极显著负相关。砧木的侧根数(X9)与砧木地径(X10)呈极显著负相关。
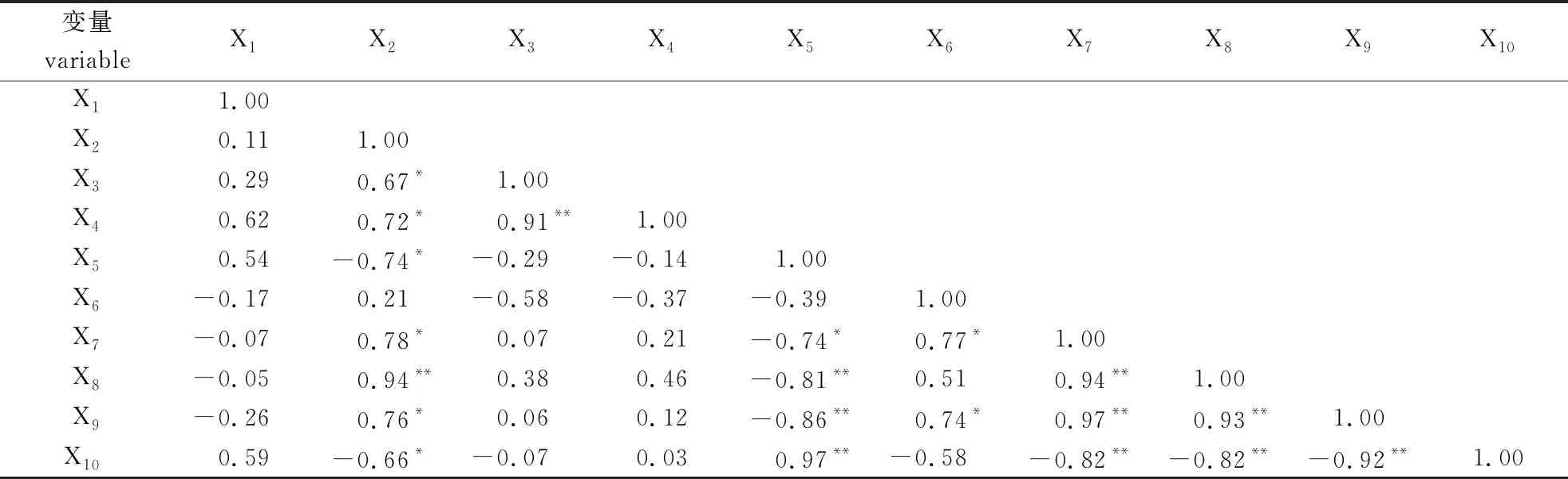
表3 接穗处理砧木生长和解剖指标间的相关分析
3 讨论与结论
嫁接被广泛地应用于农林生产,能提高植物耐胁迫能力而正常生长[16],嫁接体的砧木根系吸收提供其生长所需的水分和矿质元素[17]。而本研究发现,不同接穗显著影响砧木主根和侧根的生长,与彰武松嫁接到樟子松后的结果相同[18],嫁接促进油茶芽苗砧嫁接体根系生长,接穗与砧木亲缘关系越远,越有利于根系的生长。
根系解剖特征不但能够直接呈现出植物组织的发育状况,而且与其功能存在密切联系[10]。根尖中柱鞘是由根维管柱外围的1~2层的薄壁细胞组成,仍保留分生能力[19]。文中研究得出,嫁接提高了中柱鞘的分生能力,促进根横向生长,接穗与砧木的亲缘关系越近,横向生长能力越强;根尖的中柱鞘厚度与皮层厚度呈显著正相关,与根尖直径呈极显著正相关,这可能与接穗提供砧木根的生长素含量有关。根直径越小,吸收效率越大,皮层越薄,根系横向运输功能越强,维管柱直径越大,根系纵向运输功能越强,嫁接减弱了砧木根的吸收效率和运输能力,同源嫁接体的根吸收效率和横向运输能力弱于异源嫁接体,纵向运输能力则强于异源嫁接体,这可能与接穗和砧木的亲和性有关,接穗和砧木亲缘关系越近,亲和性越大,越有利于接穗和砧木维管束的愈合[7,10,20]。根尖内皮层细胞加厚有利于增强根细的抗逆能力,研究中砧木抗逆性与接穗的亲缘关系有关,异源嫁接减弱了砧木的抗逆能力,而同源嫁接却能增强砧木的抗逆能力[21]。
可溶性糖和蛋白是细胞生长的物质基础[22]。本研究得出,根的可溶性蛋白含量与根尖维管柱直径和砧木地径均呈极显著正相关,与砧木的侧根长度、主根数和侧根数均呈显著负相关;根的可溶性糖含量与根尖维管柱直径和砧木地径均呈极显著正相关,与砧木的侧根长度和主根数均呈显著负相关,与砧木的侧根数呈极显著负相关;嫁接减少根系营养物质贮藏,接穗与砧木的亲缘关系越远,越不利于营养物质贮藏,说明根系营养物质促进根横向生长和砧木地径生长,抑制根伸长生长和分枝,这可能是与不同的接穗影响砧木根横向输送能力和纵向输送能力有关,同源嫁接体纵向运输能力强,利于生长物质输送到地径,促进生长,而异源嫁接体横向输送能力强,利于生长物质留在根中,促进生长。MDA含量能反映受胁迫程度[23],本研究中接穗与砧木的亲缘关系的远近决定了MDA含量,关系越远,胁迫越强。CAT是植物保护机制的关键酶之一,酶活性升高,有利于提高植物的各种抗逆性[24],砧木根的CAT活性与根尖皮层厚度呈显著负相关,与根尖中柱鞘厚度和根尖直径呈极显著负相关,嫁接降低了CAT活性,接穗和砧木的亲缘关系越远,CAT活性越大,这与王淑杰等的研究相似[25],说明接穗与砧木的亲缘关系的远近决定了对砧木生长胁迫程度,关系越远,胁迫越强,通过提高根尖的厚度和CAT活性提高抗逆性。PPO是苯丙烷代谢的限速酶,有利于木质素的形成,而木质素是细胞壁的组成成分[26]。接穗与砧木亲缘关系越远,PPO活性越小,砧木根的PPO活性与根尖中柱鞘厚度和根尖直径均呈极显著正相关,说明PPO有利于根尖变粗和内皮层变薄,根尖越细,内皮层细胞加厚层次越少。POD作为一种诱导酶,存在于植物细胞壁和细胞的质外体之中,随着植株体内细胞分化的增加而提高[27]。砧木根的POD活性与根尖维管柱直径呈显著负相关,与砧木的主根长度、侧根长度、主根数和侧根数呈极显著正相关,与砧木地径呈极显著负相关,嫁接提高了POD活性,且接穗和砧木亲缘关系越远,POD活性越大,越有利于根伸长生长和分枝,越不利于根维管柱和砧木地径生长,这与王淑杰等[25]的研究结果相似。
嫁接促进油茶芽苗砧嫁接体根系生长,接穗与砧木亲缘关系越远,越有利于根系的生长。嫁接提高了中柱鞘的分生能力,促进根的横向生长,减弱了砧木细根的吸收效率和运输能力;接穗与砧木的亲缘关系越近,根横向生长能力越强,根细吸收效率和横向运输能力越弱,纵向运输能力越强,抗逆性越强。嫁接降低根系营养物质贮藏,接穗与砧木的亲缘关系越远,越不利于营养物质贮藏。嫁接降低了CAT活性,提高了POD活性;接穗与砧木的亲缘关系决定了对砧木生长胁迫程度,接穗与砧木亲缘关系越远,胁迫越强,CAT活性越大;PPO活性越小,根越细,内皮层细胞加厚层次越少;POD活性越大,越有利于根的伸长生长和分枝,越不利于根的横向生长和砧木地径的生长。因此,在立地条件优越的环境下可选与砧木亲缘关系远的接穗,在立地条件差的环境下则选与砧木亲缘关系近的接穗。
猜你喜欢
作物学报(2022年2期)2022-11-06
农业环境科学学报(2022年9期)2022-10-14
社会科学战线(2022年7期)2022-08-26
农业现代化研究(2022年3期)2022-06-29
广西植物(2022年5期)2022-06-18
文山学院学报(2022年2期)2022-05-27
课外生活·趣知识(2022年5期)2022-05-14
种子科技(2021年21期)2021-01-16
亚热带农业研究(2019年3期)2019-11-28
北方文学(2019年21期)2019-08-18
