
入境盐渍白尾鹿皮中截获的颈利虱蝇(双翅目:虱蝇科)*
2020-11-18 14:12曹秀娟高云霞何振毅
寄生虫与医学昆虫学报 2020年3期
赵 爽 曹秀娟 高云霞 何振毅
(1.中华人民共和国南沙海关,广东广州 510000;2.中华人民共和国佛山海关驻南海办事处,广东佛山 528000;3.中华人民共和国广州海关,广东广州 510000;4.中华人民共和国佛山海关驻禅城办事处,广东佛山 528000)
2018年南海口岸对来自美国明尼苏达州的白尾鹿皮集装箱进行开柜查验时,截获到虱蝇8头,经形态学和分子生物学鉴定,确定其为颈利虱蝇Lipoptenacervi(Linnaeus,1758),均为雌性。颈利虱蝇隶属于双翅目Diptera虱蝇科Hippoboscidae利虱蝇属LipoptenaNitzsch,俗称鹿蜱蝇,是欧洲鹿科动物最常见的吸血性体表寄生虫(Haighetal.,2002;Kaunistoetal.,2009)。虱蝇科全世界已知有21属213种(Dick,2006),我国已知12属44种(Maa, 1969;薛万琦等,1996;徐保海,2011)。颈利虱蝇在北欧Fennescandia半岛和美国都是入侵种类,在我国北部、东部和甘肃也有文献记录颈利虱蝇的分布(Maa, 1969;徐保海,2011)。本文将对截获样品进行详细描述,结合现有的研究资料,对该蝇的世界分布、宿主、生物学及经济和卫生重要性进行详细介绍,以便于国内口岸监管部门和鹿科动物养殖从业人员参考。
1 材料与方法
1.1 材料来源
以佛山海关南海口岸2018年6月28日截获的8头虱蝇标本为研究对象,其中1头用于分子生物学鉴定,其余标本用75%酒精浸制,保存在佛山海关驻南海办事处的医学媒介生物标本库中。
1.2 方法
1.2.1形态鉴定 使用体式显微镜(型号:奥林巴斯SZ51)进行观察,利用莱卡DVM6数码视频显微镜(Digital Microscope Leica DVM6)拍摄外部形态图片,根据标本特征和相关文献(Maa,1965;Petersonetal.,1970;Andreanietal., 2019;Salvettietal., 2020)对样品进行形态学鉴定和描述,雄性特征描述见上述文献。
1.2.2DNA提取 鉴定完毕选取其中一头标本,按照DNA提取试剂盒(TIANamp Genomic DNA Kit,目录号:DP304-02)说明书进行基因组DNA的提取。
1.2.3PCR扩增 以提取的基因组DNA为模板,正向引物LCO1490(5′-G G T C A A C A A A T C A T A A A G A T A T T G G-3′;反向引物HCO21985′-T A A A C T T C A G G G T G A C C A A A A A A T C A-3′(多细胞无脊椎动物DNA条形码通用引物,由生工生物工程(上海)股份有限公司合成,以下简称生工)。采用PCR扩增试剂盒(Taq)Ready-To-Use PCR Kit(Taq DNA polymerase)(with Loading Dye)(生工,编号:B532081-0020)进行PCR扩增,扩增条件为:94 ℃预变性5 min;94 ℃变性30 s,54 ℃退火30 s,72 ℃延伸30 s,30个循环;最后72 ℃延伸10 min。
1.2.4测序及结果比对 将PCR产物直接送至生工进行双向测序,测序结果通过Mega7.0软件进行校对拼接,去除序列两端的引物序列,得到目的片段。将所得序列在GenBank和BOLD system v4中进行同源性分析,然后使用Mega7.0软件以邻接法构建系统发育树(Neighbor-joining tree,NJ树)。
1.3 文献整理
搜集并整理颈利虱蝇相关文献,对其世界分布、寄主、生物学及携带病原体情况进行介绍。
2 结果
2.1 形态描述
鉴定特征 头宽约为长的2倍;额唇基区后缘宽平,略呈方形。前胸腹板突上的刚毛几乎分布在整个表面;肩鬃6,前4(1+3)后2排成2行;腹部背面骨片3(Tergite 3, T3)明显小于骨片4(Tergite 4, T4)。雌性生殖前板分3个骨片,两侧骨片长约为宽的2倍,略短于中央骨片。(图1~2)。

图1 颈利虱蝇雌性背面观(左)和腹面观(右)
雌性 体扁平,棕黄色,成虫翅脱落,密被深褐色鬃毛,体长6.0~7.0 mm,头部短小阔扁,前缘宽弧形,头部嵌入前胸窝内,与胸部连接紧密,活动范围极小;额唇基区(frontoclypeus area,为额囊缝至口器上片的区域)略呈方形;前口式,刺吸式口器。侧额宽,约为复眼宽的1/2;间额约与复眼等宽。额鬃3,侧额鬃4,顶鬃1。触角短,深陷入触角沟内(图2-a)。前胸背板带状,宽度较均一;前胸腹板突上的刚毛分布均匀。中胸背板具中刻线和1对侧纵刻线,侧纵刻线中间略平行,向前弧形渐宽至端部略收缩,向后微弱变宽末端不达小盾沟。肩鬃(humeral bristles)6,前4(1+3)后2排成2行,(此处所指肩鬃指中胸背板的前侧角的区域,其上鬃序排列特征是该种区别于其他种的重要特征之一)。中鬃(Acrostical bristles)8,侧中鬃(Laterocentral bristles)15~18;小盾鬃6(图2-c)。平衡棒淡黄色。足粗壮,具较密的鬃。前足基节矩退化;各足具急剧弯曲的爪,具明显爪垫和爪间突。腹部背板和侧板界限模糊,背面有明显的中央骨片,各骨片近平均分布,骨片3明显小于骨片4。雌性生殖前板(Pregenital plate)分3个骨片,两侧骨片各具2或3鬃,中间骨片具4鬃(图2-b)。
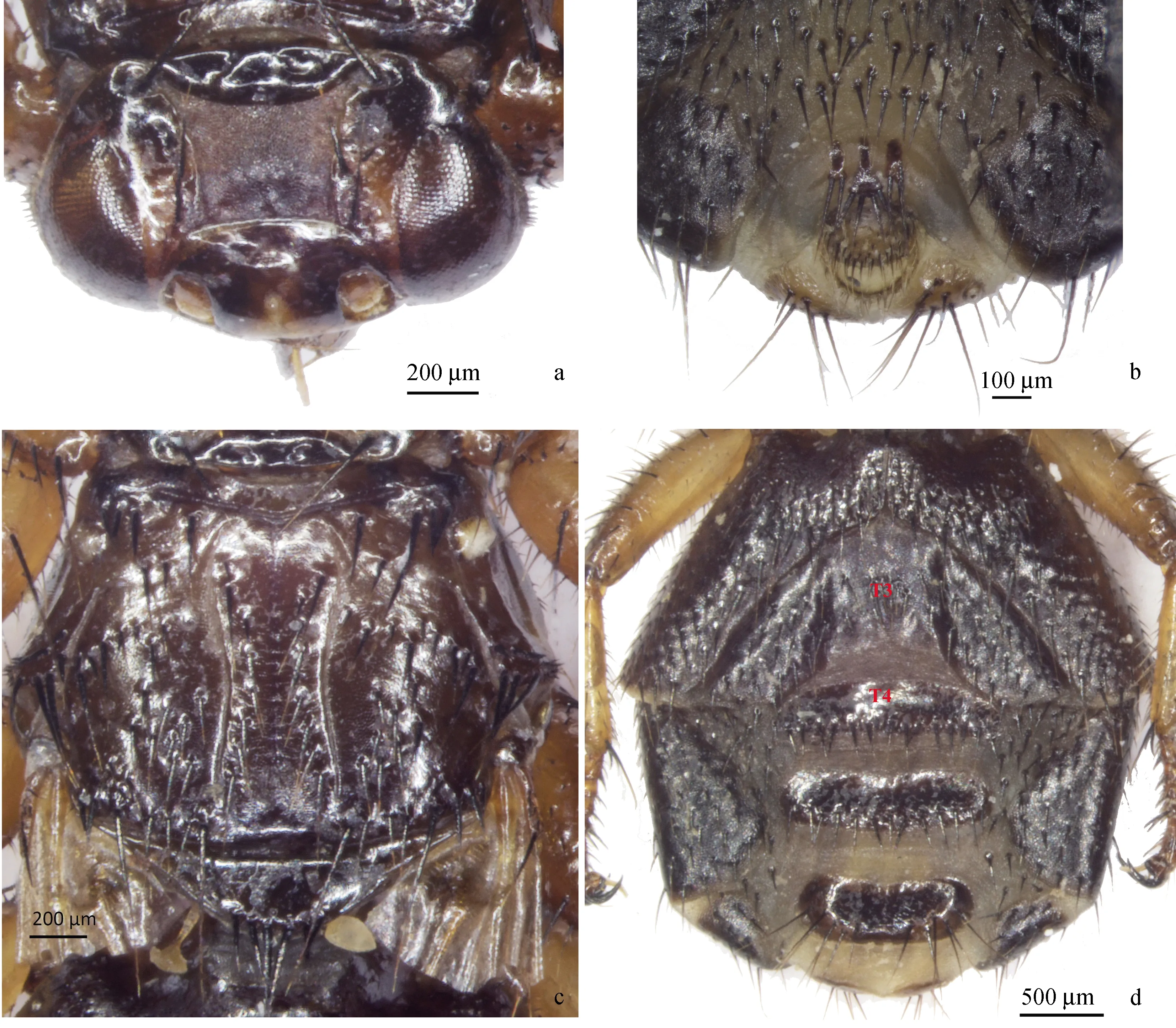
图2 颈利虱蝇雌性
2.2 序列结果及分析
截获样本的COI序列片段为661 bp,将序列输入NCBI和BOLD数据库中进行比对,与GenBank中公布的L.cervi(登录号分别为:KR362275.1和KR362270.1)的COI序列相似度为99.85%,覆盖度为98%;与BOLD System v4中的L.cervi(BIN ID: ABX1452)的COI序列相似度同样高达99.85%。
用Mega7.0软件对所得序列及NCBI中搜索到相似度高的其他虱蝇科的蝇种的序列,以须舌蝇Glossinapalpalis为外群,以邻接法构建系统发育树(NJ树)。如图3所示,通过COI序列鉴定的结果呈现出与现行传统分类系统中属级分类关系较一致的格局,除去L.mazamae(序列号:EF531205.1)的结果异常之外,NJ树中大多种的临近序列都来自同属的物种。此外,南海截获的颈利虱蝇(NHF)与来自北欧Fennescandia半岛的颈利虱蝇并为一支,并与其他利虱蝇属的种类区分开,但本研究所用样本来自北美种群,与北欧样本间出现一定的差异,这可能是由于颈利虱蝇大约在19世纪从欧洲传入美国(Bequaert,1953),在过去的一百多年间与欧洲的种群产生了地理隔断,逐渐发生了一定的遗传分化有关,但是该分化尚未达到种级阶元的差别。
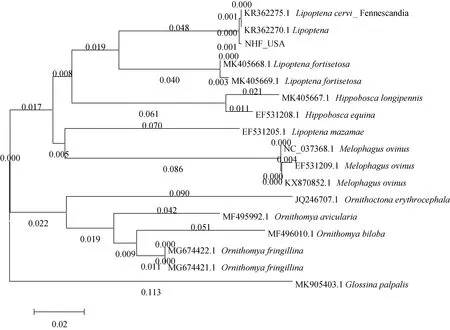
图3 基于COI基因序列构建的NJ树
2.3 颈利虱蝇的分布、宿主、生物学及医学重要性
2.3.1分布 欧洲(英国、瑞典、丹麦、荷兰、比利时、法国、德国、爱沙尼亚、拉脱维亚、立陶宛、波兰、捷克、斯洛伐克、奥地利、瑞士、匈牙利、南斯拉夫、阿尔巴尼亚、保加利亚、意大利、西班牙)、非洲(阿尔及利亚)、亚洲(俄罗斯(东西伯利亚)、白俄罗斯、中国)、北美洲(美国)(Maa, 1969;Ivanov,1974;徐保海,2011)。
2.3.2宿主 在北欧Fennescandia半岛颈利虱蝇的宿主主要是驼鹿Alcesalces,在芬兰发现其也能够在野生驯鹿Rangifertarandusfennicus体上繁殖发育,在中欧该蝇的宿主主要是红鹿Cervuselaphus、獐鹿Capreoluscapreolus和欧洲小鹿Damadama(Haarløv,1964;Hackmanetal.,1983;Välimäkietal.,2010)。在北美颈利虱蝇主要寄生于其获得性宿主白尾鹿Odocoileusvirginianus(Matsumotoetal.,2008)体表。颈利虱蝇的雌蝇随时可侵袭任何潜在的寄主(Ivanov,1974;Hermosillaetal.,2006;Kortetetal.,2010),但是其他鹿类对其宿主选择的重要性尚不清楚,仅被当做颈利虱蝇的替代宿主或潜在宿主。也有记载称牛、马等是该蝇的宿主(徐保海,2011)。偶有颈利虱蝇侵袭其他动物(如狗)甚至是人类的报道,但没有证据显示该蝇能够在鹿科以外的动物体表完成繁殖和发育(Rantanenetal.,1982)。
2.3.3生物学 颈利虱蝇是一化性吸血昆虫(Haarløv,1964)。成虫不论雌雄均可吸血,胎生,成虫交配后,幼虫在雌蝇体内发育至老熟幼虫,待该化蛹时产出。雌蝇每次仅产一头幼虫,幼虫产出后几小时内即化蛹,蛹在动物皮毛内或落到地面(Bequaert,1939),在北欧夏末或秋初时,蛹经过3~6周后即可羽化为具翅成虫。成虫可到处飞舞寻找鹿科寄主,一旦寻找到合适的寄主后,雌雄蝇的翅便会脱落并开始吸血生活(Bequaert,1953;Haarløv,1964;Hackmanetal.,1983)。雌蝇一生可产12~15头幼虫,每次间隔6~8 d。颈利虱蝇主要在鹿科动物体表毛发内繁殖,无翅成虫几乎一直生活在宿主体表毛内(Bequaert,1953)。当成蝇从宿主体表脱落后仅能存活7 d,但它们会努力爬上其他物体的高处来寻找新的合适的宿主。
2.3.4医学重要性 颈利虱蝇对宿主的侵害主要是造成失血、瘙痒、抓挠以及因此可能产生的斑秃、脱毛、甚至继发感染,最终损害宿主的皮毛质量和健康(Dehioetal.2004,Madslienetal.,2011)。被颈利虱蝇叮咬后,人和犬类也会出现长期持续的皮肤炎症,如瘙痒、丘疹,症状较顽固,可持续2周至12个月(Rantanenetal.,1982)。
近年来不时有报道从颈利虱蝇体内检出各种病原体,如在颈利虱蝇的中肠曾检出巴尔通体Bartonellaschoenbuchensis,表明该细菌能够在颈利虱蝇体内增殖,该蝇极可能是一种潜在的病媒生物(Dehioetal.,2004;Matsumotoetal.,2008;Szewczyketal.,2017),美国报道从颈利虱蝇体内检出了伯氏疏螺旋体Borreliaburgdorferi和嗜吞噬细胞无形体Anaplasmaphagocytophylum(Víchováetal., 2011;Bussetal.,2016),二者都是蜱传疾病的病原体。迄今尚没有证据证明颈利虱蝇可将病原体传播给人类,但是该蝇吸食动物血液并具有虫媒病原体宿主潜力,这增加了将病原传播给人类的风险,尤其是户外活动的人和其他野生动物。
3 讨论
颈利虱蝇在美国是一种入侵物种,大约在19世纪从欧洲传入,1907年分别在美国2个不同的地方宾夕法尼亚州和新罕布什尔州的鹿身上发现,后逐渐扩散至曼彻斯特、纽约、宾夕法尼亚和新罕布什尔州等地(Bequaert,1953,1957;Matsumotoetal.,2018)。最新的数据发现该蝇新增了康涅狄格州、新泽西州、佛蒙特州和弗吉尼亚州等4个分布地(Michaeletal.,2019),该研究也指出由于数据主要来源于文献和各博物馆的收藏以及民间狩猎组织而不是来自于全面的调查,颈利虱蝇的实际分布可能更广。
根据文献资料记载颈利虱蝇在我国东部、北部(徐保海,2011)有分布,且寄主是牛、马等,其中指出确切分布地点的是甘肃省(Maa,1969)。我国鹿科动物具有较广泛的分布地,东部、北部和甘肃等地均有鹿类的分布,颈利虱蝇可能在这些地区有分布,国外的研究也认为颈利虱蝇在鹿科之外的动物体表不能完成整个繁殖发育过程(Rantanenetal., 1982)。现阶段国内没有有关颈利虱蝇的其他记述。
颈利虱蝇在北欧和美国都有较多细致的研究,从其输入扩散的动态、对当地鹿科动物的侵染状况到该蝇的生物学及在动物皮毛内的分布以及其可能携带和传播病原体的情况等都有诸多的报道。然而,该蝇在我国是否确定分布及分布地、对我国鹿科动物的感染情况以及对鹿科动物养殖业有何影响等均无报道,这也表明我国相关物种的本底调查相当匮乏。虽然迄今还没有颈利虱蝇作为人类疾病传播媒介的确切证据,但是在该蝇体内检测出了多种病原体,并且在北美它们跟莱姆病的传播蜱媒同时寄生于白尾鹿体表,在其体内也检出了蜱传疾病病原体,这都增加了该蝇是病媒生物的潜力。考虑到该蝇在北欧和美国的输入和定殖情况,结合本次的截获,我们有理由相信该种在我国存在极高的输入风险。加强本底调查,全面掌握本底物种的物种组成和种群资料,对于尽早发现输入性物种和及时做出有效控制措施具有十分重要的意义。同时,为保护国内鹿科动物养殖产业和应对潜在的虱蝇传播疾病的风险,口岸部门应进一步加强对相关进境动物皮张的检疫监管,保卫国门卫生安全。
猜你喜欢
科学(2020年3期)2020-11-26
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
现代园艺(2017年13期)2018-01-19
小学阅读指南·低年级版(2016年12期)2017-01-05
科技知识动漫(2016年7期)2016-07-29
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年21期)2015-03-01
