
水稻灌浆期穗颈瘟发生对产量及茎节代谢产物的影响
2020-11-16 02:51:14潘争艳刘博蒋洪波姚继攀白元俊徐正进
中国农业科学 2020年20期
潘争艳,刘博,蒋洪波,姚继攀,白元俊,徐正进
(1辽宁省水稻研究所,沈阳110101;2沈阳农业大学农学院,沈阳110161)
0 引言
【研究意义】水稻(Oryza sativa)是全球约50%人口的主粮[1],由稻瘟病菌(Magnaporthe oryzae)引起的稻瘟病是世界各水稻产区普遍存在的一种病害[2],对水稻造成大幅减产,严重时可减产 40%—50%[3]。稻瘟病在水稻的各个生育时期均可发生,穗颈瘟是其中危害最大的一种[4]。植物体内代谢产物的变化是生物系统对遗传和环境变异的最终反映[5],代谢组学(metabonomics)可以对植物体内代谢产物的整体组成进行动态分析[6]。应用代谢组学方法分析穗颈瘟对水稻茎秆代谢产物的影响,可为解析穗颈瘟感病机理提供理论依据。【前人研究进展】代谢组学是连接最上游基因功能和最下游表观型变化的纽带[5],已广泛应用于水稻各个领域。ISHIKAWA等[7]利用代谢组学方法研究发现,氧化胁迫促使水稻细胞的中枢代谢途径耗竭和氨基酸积累;SANA等[8]利用代谢组学方法研究细菌性白叶枯病菌(Xanthomonas oryzaepv.oryzae)与水稻的互作发现,细菌性白叶枯病菌可以显著影响水稻的苯丙氨酸代谢;PENG等[9]研究水稻叶鞘和褐飞虱(Nilaparvata lugens)互作发现,褐飞虱对抗/感品种的诱导方式不同,褐飞虱可以诱导感病品种脂肪酸氧化、乙醛酸循环、糖异生和GABA作用的增强,还可以诱导抗病品种糖酵解和莽草酸途径的增强。稻瘟病菌是研究植物病原菌重要的模式生物,在全基因组、转录组以及抗病基因定位与克隆[10]等方面都对其有大量的研究,代谢组学方法也成功应用到稻瘟病菌与水稻互作的研究中,JONES等[11]对稻瘟病菌侵染感病型与抗病型水稻叶片的代谢产物进行分析发现,感病型与抗病型水稻叶片差别最大的代谢产物为丙氨酸。水稻茎鞘中贮藏的可溶性糖等非结构碳水化合物(nonstructural carbohydrate,NSC)和氮素是水稻增产的重要物质[12-13],抽穗前茎鞘中积累的NSC在后期被高效率转运到籽粒中是水稻产量提高的重要因素[14],魏凤桐等[15]研究表明,施氮后 NSC积累不足和转移效率降低限制了千粒重和结实率的提高。稻瘟病菌侵染茎节后,叶片和茎中的营养物质不能运输到籽粒中[16],影响水稻植株营养物质的积累、运转和分配[17],导致水稻减产且无法补救[18]。【本研究切入点】选用两个感病品种辽星1号和C418,其中辽星1号为高产优质常规稻品种,在辽宁省曾大面积种植,C418为通过“籼粳架桥”技术选育的粳型恢复系,因具有较高的配合力在全国范围内培育出较多强优势组合,随着稻瘟病菌小种变化等原因,辽星1号和C418抗病性逐渐下降。代谢组学是研究水稻响应生物胁迫和非生物胁迫的重要手段[19],在稻瘟病方面的研究多集中在叶瘟与寄主之间的互作,而自然条件下穗颈瘟发生对茎节代谢产物的变化及其对产量的影响鲜见报道。【拟解决的关键问题】以自然条件下辽星1号和C418感染穗颈瘟的茎节为研究对象,考察产量性状并利用气相色谱-质谱(GC-MS)非靶向代谢组学方法检测茎节代谢产物,分析感病茎节与正常茎节之间的差异代谢产物种类,比较糖类物质、氨基酸类物质相对含量,从代谢物角度解析水稻品种感病和产量降低的原因,为感病机理研究提供理论依据。
1 材料与方法
1.1 供试品种
选用水稻感病品种辽星1号和C418为材料,种子来源于辽宁省水稻研究所。
1.2 样品采集
试验于2019年4—10月在辽宁省水稻研究所试验基地进行。辽星1号和C418生育期田间正常管理,无药剂防治。灌浆期间进行穗颈瘟调查,计算发病率。选择仅有穗颈瘟发生的植株进行标记,无任何病害的正常植株为对照(分别为LX1-CK和C418-CK),因辽星1号和C418抽穗时期不一致,9月15日对辽星1号穗颈瘟发生的茎节(LX1-disease)和正常茎节进行取样,9月 18日对 C418穗颈瘟发生的茎节(C418-disease)和正常茎节进行取样,茎节取样后经液氮冷冻研磨成粉末,-80℃冰箱保存备用。两个样品均取样完成后进行代谢物检测。每个处理4次重复,每个重复20穗;另一部分成熟期取样进行产量及品质构成因素测定。
1.3 茎节代谢产物定量、定性分析
茎节代谢产物定量、定性测定委托具有资质的专业第三方检测机构上海鹿明生物科技有限公司完成。
1.3.1 仪器设备 代谢产物测定所需仪器包括,超声波清洗机(SB-5200DT),旋涡振荡器(TYXH-I)、研磨仪(JXFSTPRP-24/32)、离心干燥器(LNG-T98)、高速冷冻离心机(TGL-16MS)、气浴恒温振荡器(THZ-82A)、真空干燥箱(DZF-6021)、气相色谱-质谱联用仪(7890B-5977B)和色谱柱(HP-5MS(30 m×0.25 mm×0.25 μm))。
1.3.2 样品处理 精密称取 60 mg 茎秆粉末样本,放入 1.5 mL 的离心管中,加入 40 μL 内标(L-2-氯-苯丙氨酸,0.3 mg·mL-1,甲醇配置)和 360 μL 冷甲醇,在-20℃冰箱中放置2 min,冰水浴超声提取30 min,加入 200 μL 的氯仿,漩涡机中涡旋 2 min,再加入 400 μL的水,漩涡机中涡旋2 min,冰水浴超声提取30 min;-20℃静置 30 min 后低温离心 10 min,取 300 μL 的上清液装入玻璃衍生瓶中,用离心浓缩干燥器挥干样本,向玻璃衍生小瓶中加入 15 mg·mL-1的甲氧胺盐酸盐吡啶溶液80 μL,涡旋振荡2 min后,于37℃振荡培养箱中放置90 min进行肟化反应。将样本取出后再加入80 μL BSTFA(含1%TMCS)衍生试剂、20 μL 正己烷和 11种内标(C8/C9/C10/C12/C14/C16,0.8 mg·mL-1;C18/C20/C22/C24/C26,0.4 mg·mL-1,均为氯仿配置)10 μL,涡旋振荡2 min后,于70℃反应60 min。取出样本后,在室温放置30 min,进行GC-MS代谢组学分析。每6个样品中插入1个质控(quality control,QC)样本,最后4个样品插入1个质控样本,评价整个实验过程中系统质谱平台的稳定性。
1.3.3 色谱条件 HP-5MS 毛细管柱(30 m×0.25 mm×0.25 μm,Agilent J&W Scientific,Folsom,CA,USA),载气为高纯氦气(纯度不小于 99.999%),流速1.0 mL·min-1,进样口的温度为260℃。进样量为1 μL,不分流进样,溶剂延迟5 min。柱温箱的初始温度为60℃,以8℃·min-1程序升温至125℃,5℃·min-1升温至 210℃,10℃·min-1升温至 270℃,20℃·min-1升温至305℃保持 5 min。
1.3.4 质谱条件 电子轰击离子源(EI),离子源温度230℃,四级杆温度150℃,电子能量70 eV。扫描方式为全扫描模式(SCAN),质量扫描范围:m/z 50-500。
1.3.5 数据分析 将GC-MS原始数据经软件转换,导入MS-DIAL软件进行峰识别,与公共数据库Fiehn匹配相似度实现化合物定性,每个物质在质谱中的响应信息经过转换得到丰度值即为相对定量值。
1.4 产量及品质性状测定
水稻成熟后取样,测定单穗产量性状、外观品质和碾磨品质,包括结实率、千粒重、粒长、粒宽、糙米率、精米率、整精米率和垩白率等。
1.5 统计分析
运用Excel软件进行产量性状的方差分析。代谢产物数据通过在线软件(http://www.metaboanalyst.ca/)进行主成分分析(principal components analyses,PCA)、偏最小二乘法判别分析(partial least-squares discriminant analysis,PLS-DA)和差异代谢产物等分析。
2 结果
2.1 辽星1号和C418穗颈瘟发生调查
灌浆期对辽星1号和C418穗颈瘟发病率进行调查,结果表明,C418的发病率和发病部位长度均大于辽星1号。辽星1号穗颈瘟发病率为57.23%,穗颈瘟发病部位长度介于1.10—2.35 cm。C418穗颈瘟发病率为82.94%,发病部位长度介于1.52—3.64 cm。
2.2 穗颈瘟发生对辽星1号和C418产量构成因素的影响
对辽星1号和C418产量构成因素进行测定,结果表明穗颈瘟发生降低了辽星1号和C418的结实率和千粒重。辽星1号穗颈瘟发生极显著(显著)降低了一次枝梗结实率、二次枝梗结实率和千粒重,分别降低了2.91%、5.78%和 5.41%;一次枝梗的千粒重降低了2.95%,但差异不显著。C418穗颈瘟发生极显著(显著)降低了一次枝梗千粒重、二次枝梗千粒重和结实率,分别降低了 5.93%、8.22%和 7.44%,一次枝梗结实率降低了3.17%,但差异不显著(表1)。辽星1号和C418穗颈瘟发生的不同穗之间结实率和千粒重变化幅度较大,可能是由于穗颈瘟发生在不同时期造成的。
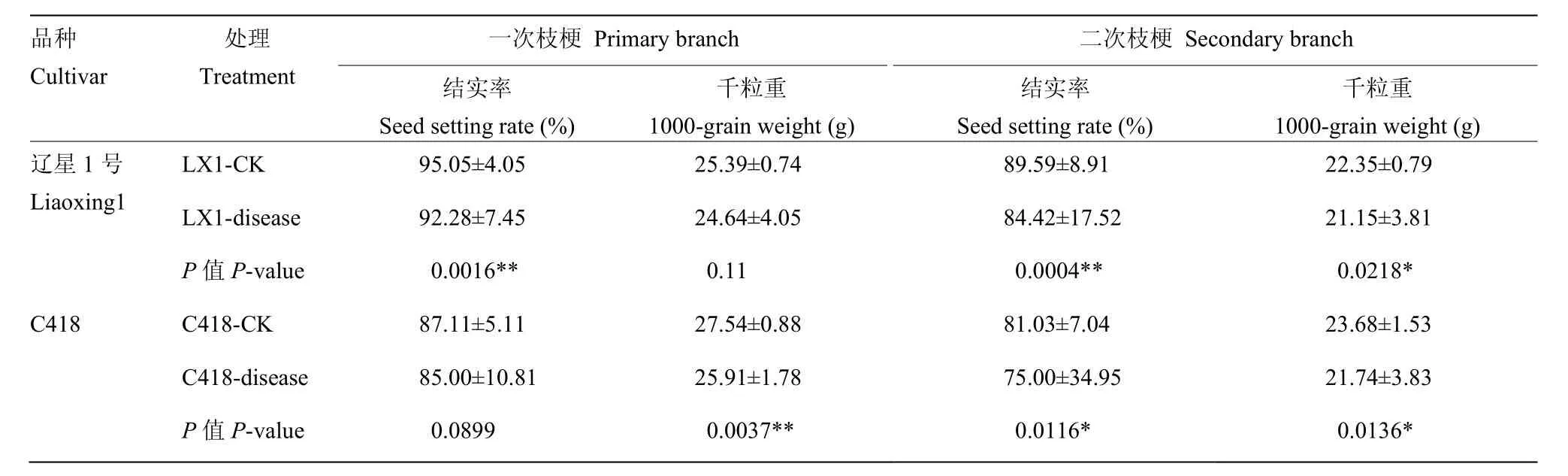
表1 穗颈瘟发生对辽星1号和C418产量构成因素的影响Table 1 Effect of panicle neck blast on Liaoxing 1 and C418 yield components
2.3 穗颈瘟发生对辽星1号和C418外观品质和碾磨品质的影响
对辽星1号和C418碾磨品质和外观品质进行测定,结果表明辽星1号穗颈瘟发生显著降低了一次枝梗和二次枝梗稻粒长度,分别降低了1.51%和1.54%,稻粒宽度略有增加,差异不显著。C418穗颈瘟发生显著降低了一次枝梗和二次枝梗稻粒宽度,分别降低3.53%和3.27%,稻粒长度差异不显著。穗颈瘟发生显著(极显著)降低了辽星1号和C418的二次枝梗稻粒的长宽比,分别降低了 8.68%和 4.83%。同时,穗颈瘟发生显著增加了辽星1号的一次枝梗垩白率,垩白率增加了54.08%(表2)。
穗颈瘟发生降低了辽星1号和C418的糙米率、精米率和整精米率。糙米率和精米率与正常差异不显著,整精米率显著(极显著)降低,一次枝梗整精米率分别降低了 7.93%和 8.79%,二次枝梗整精米率分别降低了5.15%和9.74%(表3)。
2.4 穗颈瘟发生对辽星1号和C418茎节代谢产物的影响
采用GC-MS对辽星1号和C418感病茎节和正常茎节代谢产物进行检测,均定性到358种代谢产物,辽星1号茎节感病后上调的代谢产物有263种,占检测到的代谢产物总数的73.46%;C418茎节感病后上调的代谢产物有243种,占检测到的代谢产物总数的67.88%。两组分析均上调的代谢产物有220种,占检测到的代谢产物总数的61.45%。
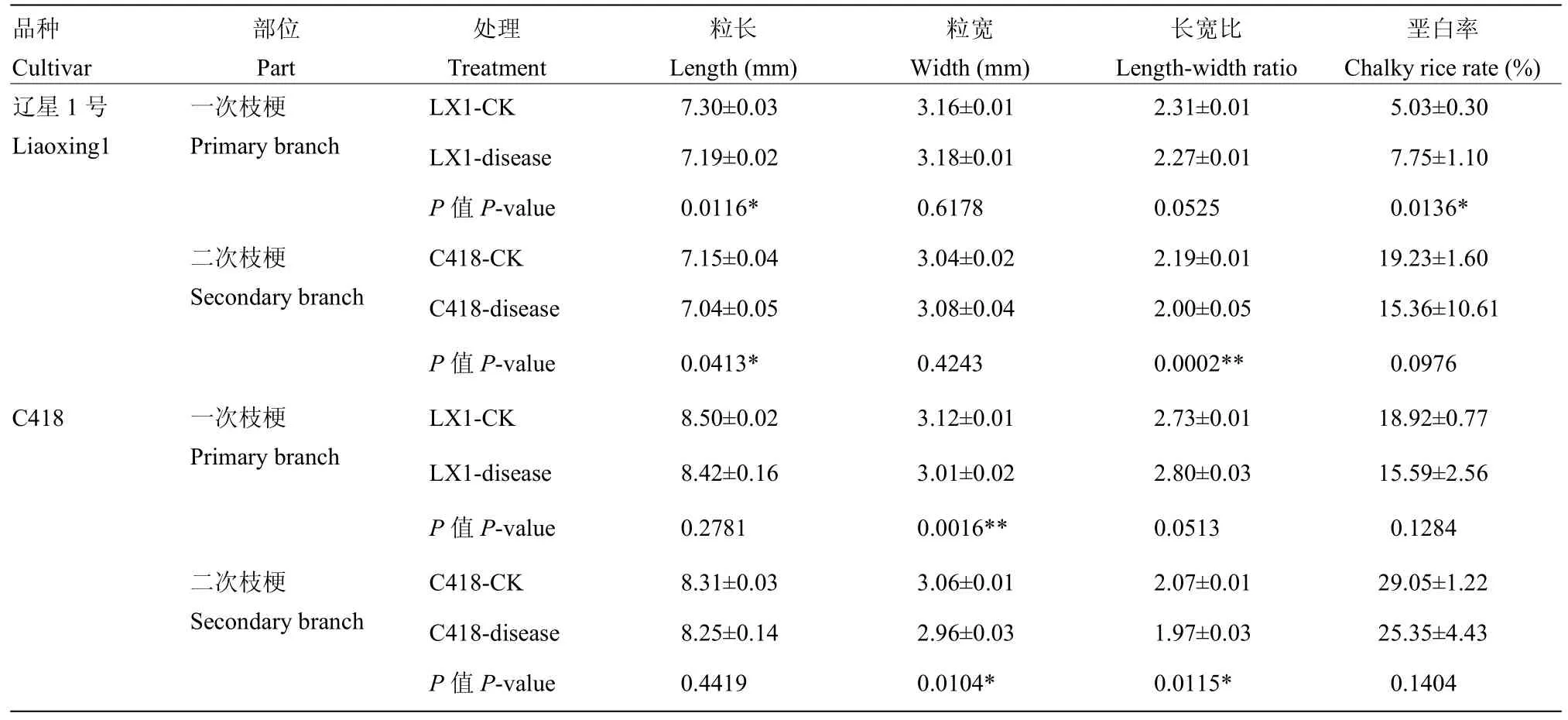
表2 穗颈瘟发生对辽星1号和C418外观品质的影响Table 2 Effect of panicle neck blast on appearance quality of Liaoxing 1 and C418
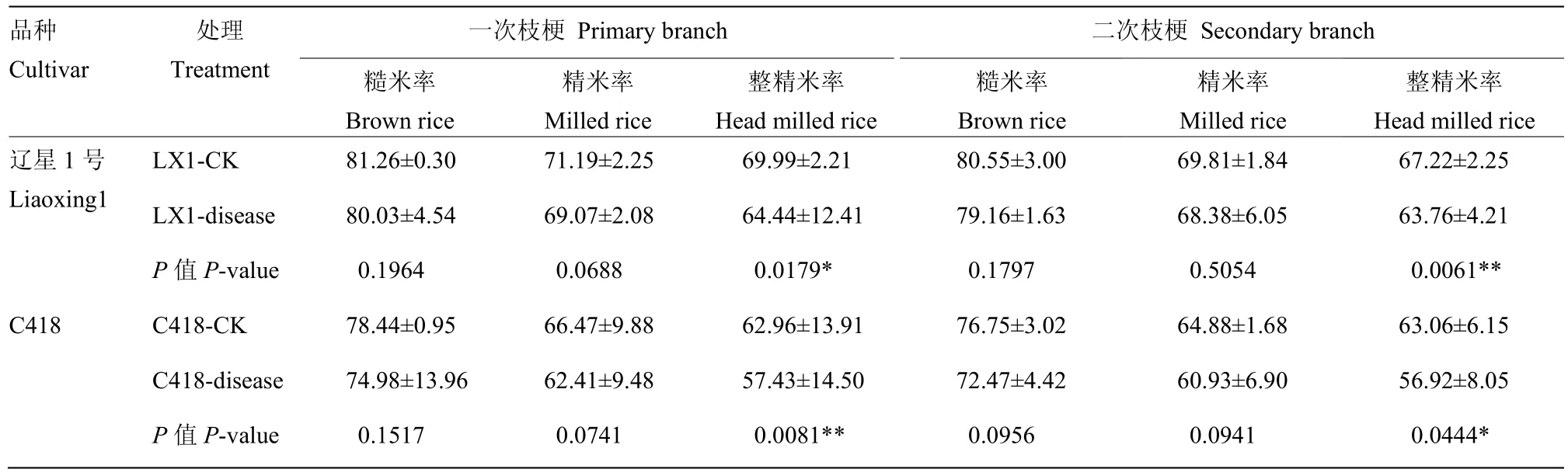
表3 穗颈瘟发生对辽星1号和C418碾磨品质影响Table 3 Effect of panicle neck blast on milling quality of Liaoxing 1 and C418 (%)
采用 PCA分析来评估所有样本的整体分布和整个分析过程的稳定性。如图1-A所示,PCA分析的两个主成分解释了总方差信息的 75.0%。所有的质控样品(QC)聚在一起,表明所有检测具有良好的分析稳定性和实验重现性。在PCA模型中,辽星1号和C418的感病茎节和正常茎节在第一个成分中被很好地分离,表明处理之间的代谢差异显著。
对检测到的代谢产物进行PSL-DA分析,辽星1号和 C418的感病茎节和正常茎节的代谢产物均沿着第一主成分清楚的分开,分别解释了总变异的 78.9%(图1-B)和82.0%(图1-C)。辽星1号的模型累积解释率R2=0.9947,Q2=0.9879;C418模型累积解释率R2=0.99726,Q2=0.99063,数值均>0.5,表明模型的拟合准确性较好。结果表明,辽星1号和C418的感病茎节和正常茎节之间具有明显不同的代谢产物表型,穗颈瘟发生严重影响了茎节的代谢物组成。
2.5 感病茎节与正常茎节之间差异代谢产物分析
利用PLS-DA分析的变量权重值(VIP>1.0)进行感病茎节与正常茎节之间差异代谢产物分析,明确促使分离的代谢产物。与正常茎节相比,辽星1号感病茎节诱导61种代谢产物显著变化,其中49种代谢产物的相对含量显著上调;C418感病茎节诱导47种代谢产物显著变化,其中24种代谢产物的相对含量显著上调。两组处理共有的差异代谢产物30种,β-谷甾醇、二甲基尿酸、α-生育酚、正壬醇、香草酸和磷酸己酸在两组分析的VIP值均>2.0,表明这6种代谢产物在感病茎节和正常茎节的代谢产物分离中起到重要作用。
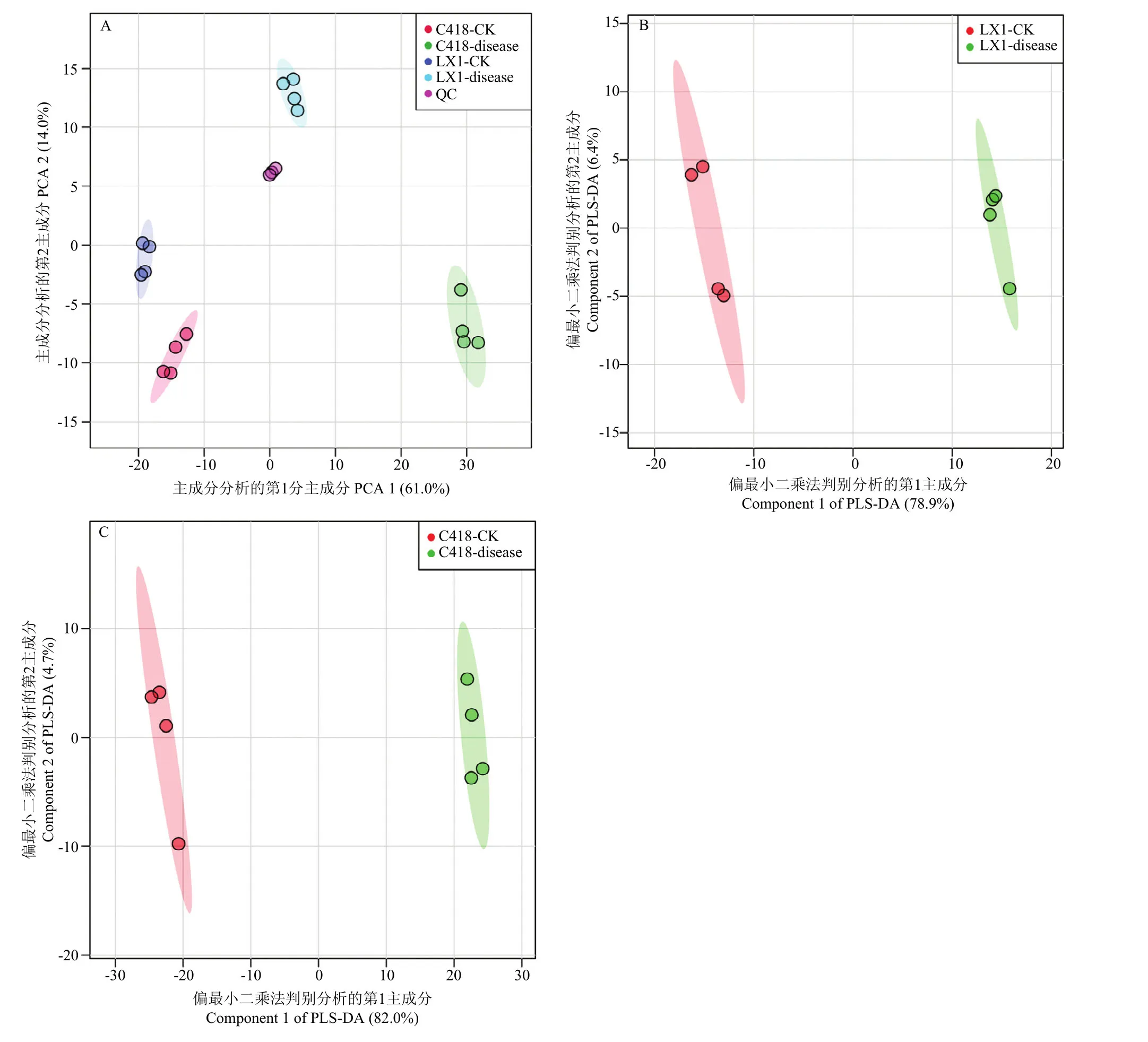
图 1 穗颈瘟发生茎节与正常茎节之间代谢产物 PCA和PLS-DA分析Fig. 1 PCA and PLS-DA analysis of the metabolites between diseased stem nodes and normal stem nodes
根据代谢产物相对含量的变化可以将 30种代谢产物分为4组(图2)。第1组19种代谢物,包括高丝氨酸、别嘌呤醇、D-红鞘氨醇、阿拉伯糖醇、山梨醇、β-谷甾醇、半乳糖-6-磷酸、乙酰氨基葡萄糖、菊粉糖、羟基氨基甲酸酯、葡萄糖-6-磷酸、D-果糖-1,6-二磷酸、磷酸己糖酸、二甲基尿酸、2-单油酸、葡萄糖酸、尿囊酸、N-乙酰半乳糖胺和5-甲基尿苷。与正常茎节相比,辽星1号和C418穗颈瘟发生导致这19种代谢产物相对含量上调,分别上调了3.83—2 763.97和10.01—18 989.02倍。第2组9种代谢物,包括甲基α-呋喃硫醚、2,3-二羟基-2-丁烯二酸、亚甲基二磷酸盐、脱氢抗坏血酸、香草酸、N-乙酰-d-甘露糖胺、三十烷醇、α-生育酚和大豆苷元。这一组代谢产物具有两个特点,一个是与正常茎节相比,辽星 1号和C418穗颈瘟发生导致这组代谢产物相对含量下调,分别下调了73.87%—96.29%和73.02%—99.68%;二是正常茎节中这组代谢产物辽星 1号的相对含量大于C418的相对含量。第3组和第4组分别有一种代谢物,分别是 1,5-脱水葡萄糖醇和正壬醇,与对照相比,辽星1号和C418感病茎节中1,5-脱水葡萄糖醇分别上调了6.91倍和下调了86.23%,正壬醇分别下调了80.03%和85.22%。由于正常茎节中C418的正壬醇相对含量大于辽星1号的正壬醇相对含量,所以将正壬醇与第2组分开,单独独立为一组。
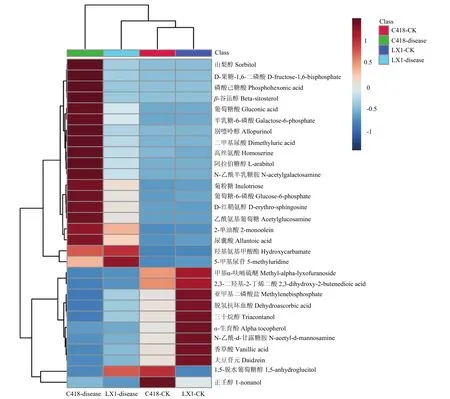
图2 差异代谢产物层次聚类分析生成的热图Fig. 2 Hierarchical cluster analysis of differential metabolites resulting in thermal energy maps
2.6 穗颈瘟发生对辽星1号和C418茎节糖类物质的影响
GC-MS共检测到糖类物质30种,从热图中可见(图3-A),与正常茎节相比,辽星1号穗颈瘟发生上调的糖类物质有21种,占糖类物质总数的70%;C418穗颈瘟发生上调的糖类物质有22种,占糖类物质总数的 73.33%。两个品种中共有的上调糖类物质19种,占糖类物质总数的63.33%。分别为阿拉伯糖、葡萄糖、异麦芽糖、纤维二糖、D-核糖、蜜二糖、三糖、菊粉糖、酮己糖、D-木糖、β-龙胆糖、槐糖、松三糖、D-果糖、D-甘露糖、D-岩藻糖、哌啶-α-d-果糖、葡庚糖和赤鲜糖。与对照相比,这19种糖类物质在辽星1号和C418感病茎节中分别上调了1.02—6.22和1.07—17.92倍。除赤藓糖和葡庚糖,其余17种代谢产物在C418感病茎节中上调的幅度更大。与对照相比,两个品种感病茎节中共有下调的代谢产物6种,包括左旋葡聚糖、棉子糖、D-塔格糖、D7-葡萄糖、海藻糖和6-脱氧葡萄糖,这6种糖类物质在辽星1号和C418感病茎节中分别下调了2.16%—21.80%和7.15%—99.77%。
2.7 穗颈瘟发生对辽星1号和C418茎节氨基酸类物质的影响
GC-MS共检测到氨基酸类物质38种,从热图中可见(图 3-B),与对照相比,辽星 1号感病茎节中上调的氨基酸类物质有31种,占氨基酸类物质总数的 81.58%;C418感病茎节中上调的氨基酸类物质有 29种,占氨基酸类物质总数的 76.32%。两个品种共有上调的氨基酸类物质26种,占氨基酸类物质总数的68.42%。上调的氨基酸有L-半胱氨酸甘氨酸、草氨酸、L-天冬氨酸、丙氨酸、β-丙氨酸、L-异亮氨酸、丝氨酸、L-苯丙氨酸、氰基丙氨酸、N-甲基丙氨酸、氧化谷氨酸、高丝氨酸、甘氨酸、L-缬氨酸、L-天冬酰胺、L-蛋氨酸、鸟氨酸、赖氨酸、邻琥珀酰高丝氨酸、L-谷氨酸、谷氨酸、L-酪氨酸、L-谷氨酰胺、L-脯氨酸、亮氨酸和苏氨酸。这些代谢物在辽星 1号感病茎节中上调了 1.05—7.98倍,在C418感病茎节中上调了1.10—13.12倍。两个品种感病茎节中共有的下调代谢产物4种,包括 N-甘氨酰脯氨酸、D3-色氨酸、N-甲基谷氨酸和N-乙酰鸟氨酸,这4种氨基酸在辽星1号感病茎节中下调了8.96%—28.19%,在C418感病茎节中下调了2.78%—32.33%。
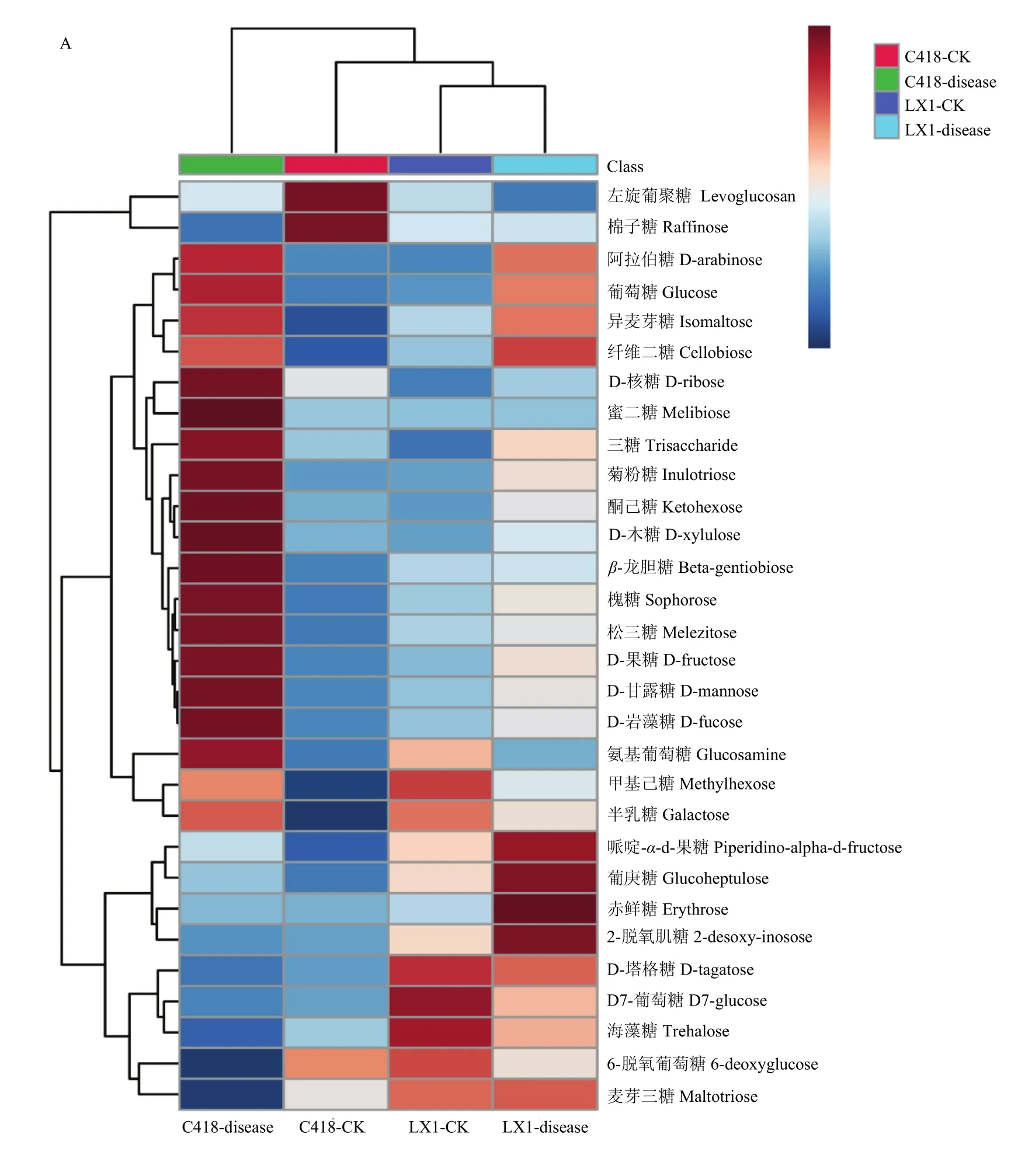
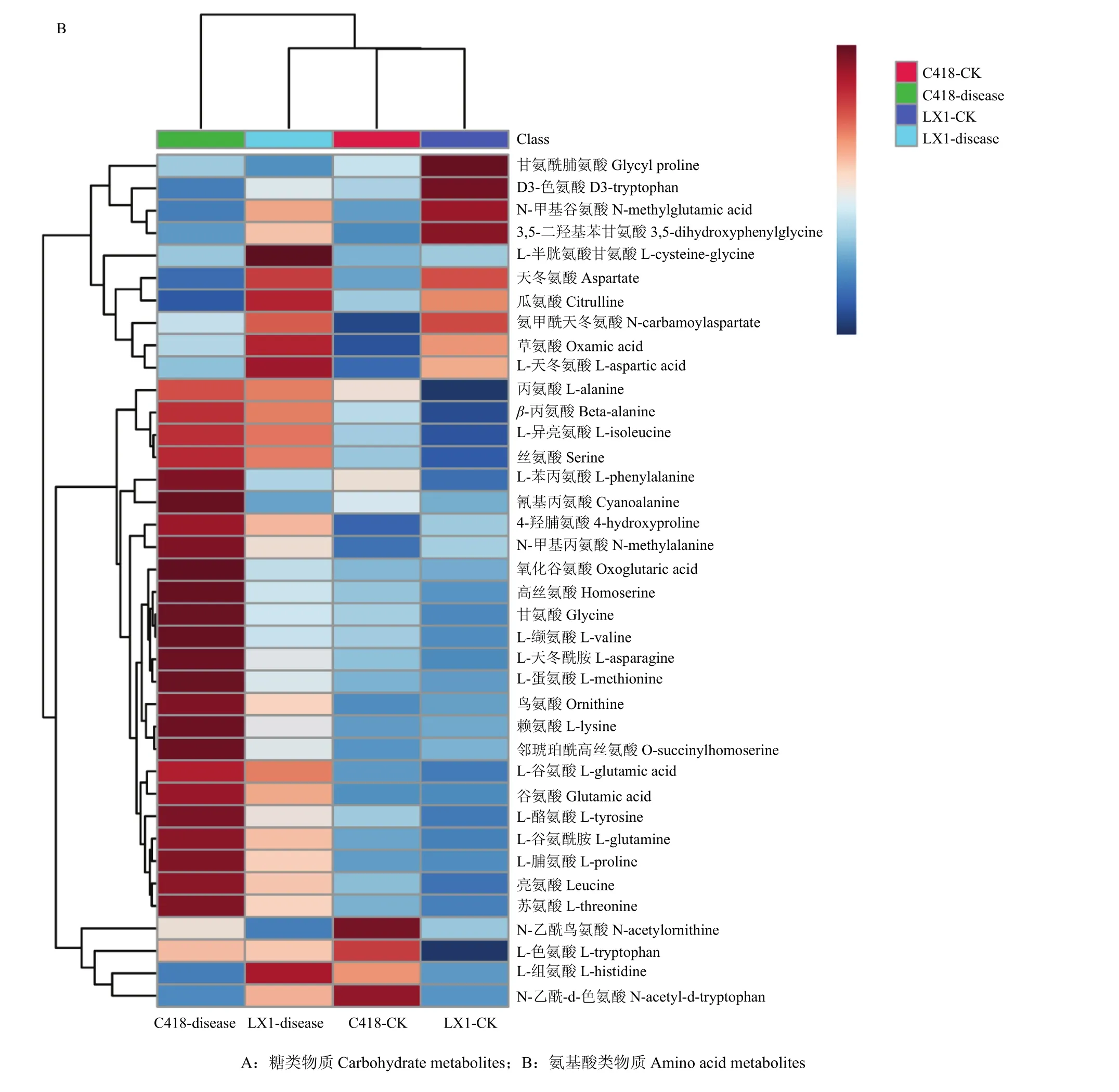
图3 糖类物质和氨基酸类物质层次聚类分析生成的热图Fig. 3 Thermal energy maps of hierarchical cluster analysis of carbohydrate metabolites and amino acid metabolites
3 讨论
次生代谢产物在植物应对生物胁迫中起着重要防御作用[20]。本研究发现,穗颈瘟发生诱导辽星1号和C418香草酸、N-乙酰-d-甘露糖胺、脱氢抗坏血酸、三十烷醇、α-生育酚和大豆苷元的相对含量显著下调,这6种物质均属于次生代谢产物范畴。生育酚和脱氢抗坏血酸作为抗氧化物质可以保护植物新陈代谢免受应激损伤[21]。三十烷醇广泛存在水稻等植物的蜡质中[22-23],能够促进碳氮代谢,加速茎中贮藏的有机物向籽粒运输[24]。大豆苷元未见水稻方面的报道,但是豆科植物中常见的次级代谢产物[25],可以提高豆科植物对病原菌的抗性。香草酸作为化感类物质在植物生长发育过程和抗逆中起到一定作用[26-27]。本研究还发现,辽星1号正常茎节中这6种次生代谢产物的相对含量分别是C418的2.64、2.39、2.39、2.27、3.09和2.56倍,穗颈瘟发病率调查结果与这一结果相反,辽星1号发病率和发病程度均小于C418。推测这6种次生代谢产物的相对含量与水稻茎节感染穗颈瘟有一定关系,稻瘟病菌可能诱导水稻茎节下调这些次生代谢产物相对含量降低茎节对稻瘟病菌的防御反应。
禾谷类作物倒1节间主要发挥物质运输通道的功能[28],抽穗后水稻茎鞘中70%左右的非结构性碳水化合物(NSC)通过倒1节转运到籽粒中,对最终产量的表观贡献率可达 24%—27%[29-30]。本研究发现,穗颈瘟发生上调了辽星 1号和 C418茎节中 70%和73.33%的糖类物质,其中阿拉伯糖上调了 6.22%和7.45%,菊粉糖上调了5.68%和17.92%,葡萄糖上调了3.0和4.80倍,甘露糖上调了1.39和3.43倍,果糖上调了1.92和2.97倍。这些可溶性糖的上调表明水稻“源”向“库”输入的强度和“库”将糖转化为淀粉的能力降低,同时果糖的上调也预示着茎中蔗糖转化酶异常活跃[31]。本研究还发现,C418穗颈瘟发生有17种糖类物质上调幅度大于辽星 1号,并且,C418穗颈瘟发生二次枝梗结实率和千粒重下降幅度也大于辽星1号,分析原因,可能是感病茎节碳水化合积累,降低了茎节光合作用,茎秆运输到籽粒上的碳水化合物量减少,导致产量下降,尤其C418产量下降更多。
水稻籽粒的形成除了需要累积大量的碳水化合物外还需要大量的有机氮类代谢产物,茎鞘中的氮素对籽粒氮的贡献率达到30%以上[32]。植物体内游离氨基酸直接或是间接对生物胁迫和水稻生长发育作出响应[33-34]。本研究表明,穗颈瘟发生上调了辽星1号和C418茎秆中81.58%和76.32%的氨基酸类物质,破坏了氮素在营养器官和生殖器官的分配比例,降低了氮素向穗部的再运转量,导致产量的降低。有研究也表明,大量丙氨酸产生可能导致植物细胞死亡,促进稻瘟病菌侵染[10],本研究结果与这一结论一致,稻瘟病菌侵染诱导了辽星1号和C418茎节中丙氨酸上调了3.34和3.74倍。
穗颈瘟发生后,辽星1号和C418二次枝梗的结实率和千粒重较一次枝梗下降显著,分析原因可能是强势粒在不同环境下存在遗传稳定性,而弱势粒灌浆受环境因素影响大[35]。穗颈瘟对精米率和糙米率没有显著影响,但对整精米率影响较大,可能是一方面碳水化合物合成受阻易造成籽粒充实不良[36],另一方面,穗颈瘟发生造成“源”输出不足,籽粒灌浆速率极慢,充实度差容重极轻,造成整精米率大幅度降低[37]。
4 结论
穗颈瘟发生显著(极显著)影响了辽星 1号和C418的二次枝梗结实率和千粒重,改变了茎节代谢产物组成,穗颈瘟可能通过下调脱氢抗坏血酸、α-生育酚、N-乙酰-d-甘露糖胺等次生代谢产物相对含量降低了水稻茎节对稻瘟病的防御反应。感病茎节通过积累大量的糖类物质和氨基酸类物质,导致碳氮代谢受阻,降低了其向籽粒转运,从而抑制了水稻产量。
猜你喜欢
湖南农业科学(2021年11期)2021-12-21 09:28:24
作物学报(2021年10期)2021-08-05 11:30:40
北方水稻(2021年2期)2021-04-26 08:25:56
中国稻米(2021年2期)2021-04-04 15:44:10
长江大学学报(自科版)(2018年22期)2018-12-03 03:31:42
湖北农业科学(2017年6期)2017-04-26 14:37:05
农学学报(2016年2期)2016-12-22 01:43:16
中国稻米(2016年4期)2016-08-15 11:53:05
分析测试学报(2015年4期)2016-01-13 06:18:27
中国烟草学报(2012年6期)2012-04-09 07:41:40
