
不同肥力潮土的酶活计量比特征及其与微生物量的关系
2020-11-16 02:51:06张露张水清任科宇李俊杰段英华徐明岗
中国农业科学 2020年20期
张露,张水清,任科宇,李俊杰,段英华,徐明岗,4
(1吉林农业大学资源与环境学院,长春 130118;2中国农业科学院农业资源与农业区划研究所/耕地培育技术国家工程实验室,北京 100081;3河南省农业科学院植物营养与资源环境研究所,郑州 450002;4中国热带农业科学院南亚热带作物研究所,广东湛江 524091)
0 引言
【研究意义】土壤作为植物生产的根本,是农业的基本生产资料,其本质是肥力。土壤肥力一方面影响土壤蓄纳和稳定供应养分的能力,另一方面影响微生物活性和根系生长,进而影响养分在土壤-作物系统中的高效循环利用。我国土壤肥力差异大,一般来说,高肥力土壤上作物产量和养分转化利用能力均较高。相同施肥量下,高肥力土壤上的玉米产量较中肥力和低肥力上高12.4%和23.1%[1],且氮肥损失量可降低大约11%和22%[2]。多年来,我国相继实施了有机肥补贴、秸秆还田等一系列措施来提高土壤肥力,而这些措施的核心都是提高土壤有机质水平。土壤有机质(包含有机碳、有机氮和有机磷)的形成和矿化既直接影响养分库的构成,有机碳又以供给微生物能量来调控微生物-土壤-作物互作功能。因为植物体和土壤微生物的形成都需要一定量的碳氮磷比(C∶N∶P),因此C∶N∶P对土壤有机质的矿化或形成具有重要的指示作用,表征了土壤中养分的丰缺状况,是指示土壤肥力的一个重要指标[3]。阐明不同肥力土壤上的C∶N∶P差异及影响因素对提高土壤肥力、作物产量和养分利用效率具有重要意义。【前人研究进展】对于土壤C∶N∶P的确定,前人已进行了大量的研究。全球范围内的土壤全量C∶N∶P为186∶13∶1[4],我国整个剖面(250 cm)的土壤全量 C∶N∶P约为60∶5∶1,有机土壤表层全量C∶N∶P为134∶9∶1[5]。通常来说,土壤C∶P要比土壤C∶N变异性程度大,主要原因在于磷元素并不是腐殖酸和棕黄酸的结构组分,且不参与大气循环[6]。微生物是土壤中最为活跃的部分,微生物量C、N、P是土壤活性养分的储存库,微生物量 C∶N∶P体现了养分是否满足微生物生长所需,决定了土壤C、N、P的活化程度,因此C、N和 P的有效性归根结底取决于微生物量 C∶N∶P(SMBC∶SMBN∶SMBP)。土壤微生物生物量C∶P低,表明与土壤中的碳相比,磷素是相对富足的,此时微生物会矿化土壤有机质补充土壤碳库,使得土壤碳对微生物的限制进一步加大;而微生物生物量C∶P高,则表明与土壤中的磷素相比碳相对富足、磷素相对限制,此时微生物为了维持自身的正常生长发育需要同化更多磷素,表现为固定磷素能力的提高,这种情况一般可能会造成与植物对磷素的竞争。土壤微生物通过胞外酶将有机态转化为无机态,显著改变C∶N∶P比例和养分有效性。BG和CBH是与碳转化密切相关的酶,NAG和LAP是与氮转化密切相关的酶,AP是与磷转化密切相关的酶,这些胞外酶的活性与微生物代谢、养分的生物循环密切相关。在目前的研究中,常将(BG+CBH)∶(NAG+LAP)、(BG+CBH)∶AP和(NAG+LAP)∶AP与养分元素C∶N、C∶P和N∶P相联系,用来评价微生物生物量C、N、P养分供给情况,继而提出了土壤酶化学计量学的概念。多数研究表明,施用有机肥料,增加外源有机物质的投入可以提高土壤中微生物的活性,从而显著提高土壤微生物量碳、氮含量及土壤酶活性,根据土壤中碳氮磷元素的适宜比例,微生物会对限制元素优先进行转化,使得土壤胞外酶C∶N、C∶P和N∶P的响应不尽相同[7-8]。【本研究切入点】土壤胞外酶C∶N∶P在全球尺度上的比值约为1∶1∶1,说明在生物地球化学循环之间C、N、P营养元素存在相互耦合关系[4]。但不同的土壤有机质水平和施肥量会导致土壤C∶N∶P的差异,从而产生不同的微生物量C∶N∶P关系和群落结构,从而导致碳氮磷酶活比的不同[9-10]。那么,不同肥力土壤上胞外酶C∶N∶P差异情况如何,是受单一养分限制,还是受其比例的限制?对于因地制宜地合理施肥和培肥土壤具有重要意义,亟需进行深入分析。【拟解决的关键问题】潮土区是我国粮食生产的主产区,本研究选取了不同肥力水平的潮土,分析了土壤微生物量和胞外酶C∶N∶P的差异,结合土壤养分指标,旨在通过生态化学计量学方法,明确不同肥力土壤的酶化学计量学特征及其养分限制因素,为土壤肥力提升和肥料合理施用提供科学依据。
1 材料与方法
1.1 试验点基本情况
试验地位于河南省农业科学院河南现代农业开发基地(东经 113°41′47.66″—113°42′00″,北纬35°00′28.43″—35°50′00″),属暖温带大陆性季风型气候。四季分明,光照充足,年均气温14.5℃,年均积温5 300℃,≥10℃积温5 169℃,年均降水量615.1 mm,年蒸发量 1 450 mm,年均日照时数 2 324 h。试验地土壤为潮土,成土母质为黄河冲积物。
1.2 试验设计
本研究选取了基地内及周围5个不同养分含量的潮土(0—20 cm),依次记为S1—S5,土壤质地均为砂质壤土。其中,S1为黄河河漫滩土壤,养分含量低,试验之前长期处于撂荒状态,未种植任何作物;S2—S5 为农田土壤,氮肥施用量为 165—281 kg·hm-2,磷肥施用量和钾肥施用量均为82.5—93.8 kg·hm-2。种植作物均为小麦-玉米轮作。土壤样品采集前 S2—S4的玉米产量水平约为 6 750 kg·hm-2,S5 的玉米产量水平约为9 750 kg·hm-2,土壤基础理化性状见表1。根据农用地利用等别[11],可将 S1划分为低肥力土壤,S2—S4为中肥力土壤,S5为高肥力土壤。
1.3 测试方法
试验于2018年10月10日取样,采用五点取样法,用直径为2 cm的土钻采取0—20 cm耕层土壤,每个肥力土壤采取3次重复,新鲜土壤过2 mm筛并挑根后,4℃保存供分析SMBC和SMBN,-20℃保存供分析BG、CBH、NAG、LAP、AP酶活性。
微生物量碳和微生物量氮采用氯仿熏蒸-K2SO4浸提法[12]。土壤样品从4℃冰箱里拿出后,在25℃培养箱里培养1周使微生物恢复活性,称取12.50 g土壤样品放入真空干燥器,用氯仿熏蒸24 h,以未熏蒸土壤作为对照,同时测定土壤样品含水量,然后加50 mL(水土比 4∶1)0.5 mol·L-1优级纯 K2SO4溶液振荡、过滤,滤液中的SMBC和SMBN采用multi N/C3100总有机碳氮分析仪测定。
土壤胞外酶活性采用96孔酶标板荧光分析法[10]。取 1.00 g 新鲜土壤样品,加入 100 mL 50 mmol·L-1的醋酸钠缓冲溶液(pH为土壤样品pH的平均值,如样品之间 pH相差较大,需配置不同缓冲溶液),用磁力搅拌器使其均质化后移取 200 μL悬浮液于酶标板中,以缓冲液为空白,4-甲基羟基香豆素(MUB)为标准物(LAP采用 7-氨基-4-甲基香豆素(AMC)为标准物),不同胞外酶相应加入对应的底物,25℃黑暗培养 4 h 后(酸性土壤需加入 50 μL 1 mol·L-1的NaOH溶液终止反应),在激发光365 nm,发射光450 nm条件下,用Synergy H/M酶标仪测定其荧光度,胞外酶活性以每克每小时干物质产生底物的纳摩尔数(nmol·g-1·h-1)计算。
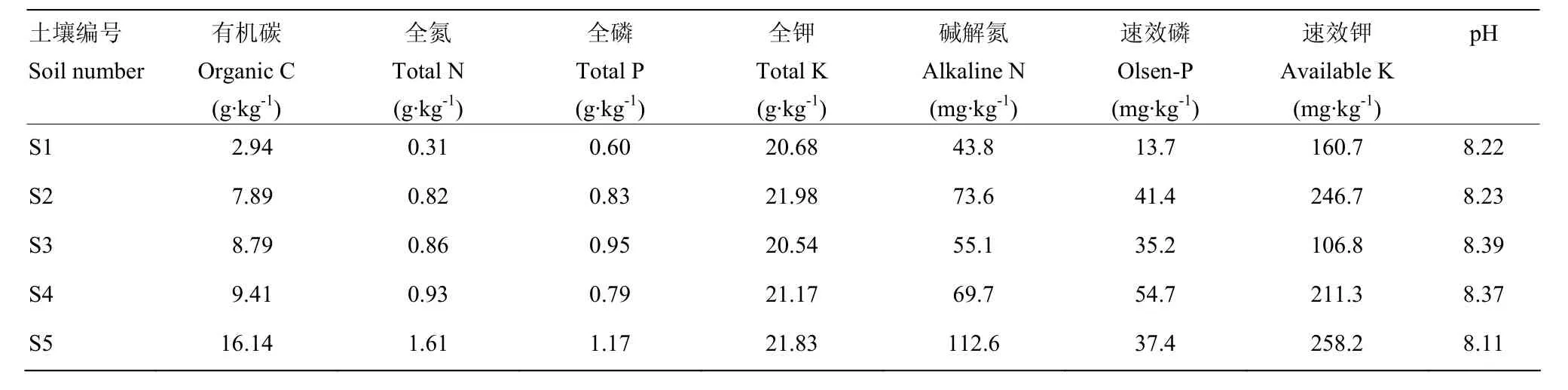
表1 不同肥力水平土壤的基础性状Table 1 Basic properties of soil with different fertility levels
土壤综合酶指数(IEI)计算[13]分为3个步骤:因子的选择、权重的确定和综合指标的获得。由于土壤因子变化具有连续性,故各评价指标采用连续性质的隶属度函数,并从主成分因子负荷量值的正负确定隶属度函数分布的升降,与各因子对植被的效应相符。对碳氮磷相关酶采用升型分布函数。IEI计算公式如下:

式中,IEI为土壤综合酶指数,IEI(xi)表示土壤酶隶属度值,wi表示土壤酶(i)的权重。
升型分布函数的计算公式如下:
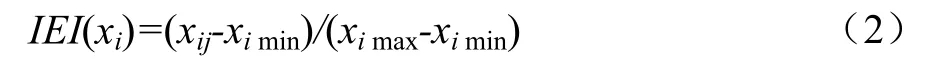
式中,xij表示土壤酶活性值,ximax和ximin分别表示土壤酶(i)活性的最大值和最小值。
由于土壤质量的各个因子的状况与重要性通常不同,所以通常用权重系数来表示各个因子的重要程度。权重系数的确定有许多方法,本研究利用主成分分析因子负荷量计算各因子作用的大小,确定其权重。利用下式计算:

式中,Ci为公因子方差,C为公因子方差之和。
土壤有机碳采用重铬酸钾容量法,全氮采用开氏定氮法,全磷采用高氯酸-硫酸-钼锑钪比色法,全钾采用高氯酸-硫酸- 火焰光度法,碱解氮采用碱解扩散法,速效磷采用NaHCO3浸提-钼锑钪比色法,速效钾采用NH4OAC浸提-火焰光度法,pH采用玻璃电极法(土水比1∶2.5)[14]。
1.4 数据处理
数据通过 Microsoft Excel 2019软件进行数据处理,差异显著性检验采用SPSS22.0软件包进行单因素方差分析(P<0.05),选择Duncan多重比较分析土壤间差异,冗余分析(RDA)采用Canoco5绘制,其他图表采用Excel和SigmaPlot12.5软件完成。
2 结果
2.1 土壤微生物量碳氮的差异
由表2可见,SMBC和SMBN均呈现出随土壤肥力提高而增加的规律:S5>S4>S3,S2>S1。总的来说,高肥力土壤的SMBC约为中肥力土壤的2.6倍,低肥力土壤的5.8倍;高肥力土壤的SMBN约为中肥力土壤的 3.1倍,低肥力土壤的 5.5倍。除了 S2的SMBC/SMBN较高外,其他土壤的 SMBC/SMBN为7.66—9.56,没有显著差异。
SMBC/SOC和SMBN/TN一定程度上表明了土壤中C和N的生物有效性。由表2可见,SMBC/SOC和SMBN/TN均为S5和S1土壤最高,且无显著差异,均显著高于中肥力土壤。中肥力土壤中,SMBN/TN为S2土壤最低,S4土壤最高。
2.2 土壤胞外酶活性的变化以及土壤酶指数
由图1-a可见,土壤碳转化相关的BG和CBH的活性在S5土壤上最高,S2—S4其次,S1土壤最低。其中,S5土壤的BG和CBH活性在为S1土壤的 8.1倍,在 S2、S3、S4土壤为 S1的 3.7、2.7、2.7倍。与碳转化相关酶活性类似,土壤氮转化相关的NAG和LAP的活性由图1-b所示,也表现为在S5土壤上最高,S1土壤上最低。其中,S5土壤的NAG和LAP活性为S1土壤的4.6倍,S2、S3、S4为S1土壤的2.2、2.4、2.2倍。从图1-c可以看出,AP的活性S5土壤最高,为S1的5.3倍;在S2、S3、S4其次,为S1的2.3、2.8、2.7倍,S1土壤最低,中等肥力土壤间差异不显著。
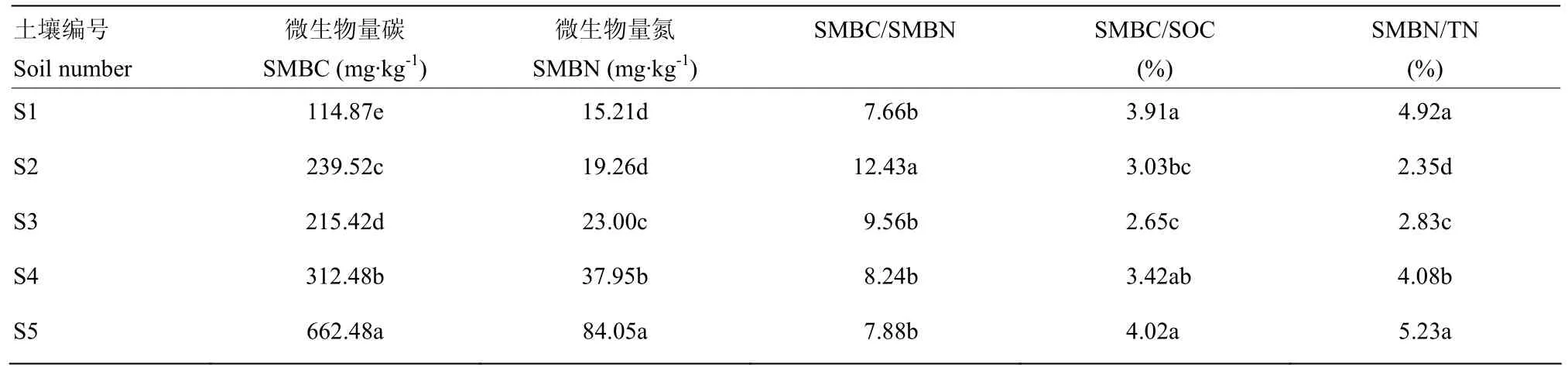
表2 不同肥力土壤的微生物量碳氮Table 2 Microbial biomass carbon and nitrogen in different fertility soils
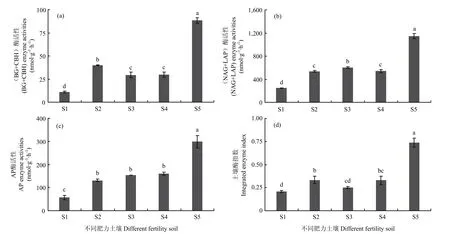
图1 不同肥力土壤碳氮磷转化相关胞外酶活性和综合酶指数Fig. 1 Extracellular enzyme activity and integrated enzyme index related to carbon, nitrogen and phosphorus transformation in different fertility soils
土壤酶指数(IEI)是所有土壤胞外酶活性的综合反映,能够全面、客观地反映土壤胞外酶活性的变化过程。从图1-d可以看出,不同肥力土壤上的IEI存在着显著性差异。高肥力土壤S5的IEI最高,为0.74;S2、S3、S4为0.33、0.25、0.33,S1土壤为0.21。
2.3 不同肥力土壤胞外酶活性与微生物量碳氮的关系
土壤碳转化相关酶(BG+CBH)活性与SMBC(图2-a),和土壤氮转化相关酶(NAG+LAP)活性与SMBN(图 2-b),均呈直线正相关关系,决定系数分别为0.924和0.847,说明SMBC和SMBN的提高会显著提高碳和氮的相关酶活性,且提高幅度在不同肥力梯度上无显著差异。SMBC和SMBN每增加1 mg·kg-1,(BG+CBH)和(NAG+LAP)活性可分别提高0.134和 10.53 nmol·g-1·h-1。
高肥力土壤S5的碳氮转化相关酶活性和SMBC、SMBN均显著高于其他肥力土壤,而低肥力土壤 S1土壤碳氮转化相关酶活性以及SMBC、SMBN含量均最低,中肥力土壤 S2—S4的碳氮转化相关酶活性以及SMBC含量相近,但SMBN含量表现为S4>S3>S2,说明在相同SMBN含量下,氮转化相关酶活性依次表现为 S2>S3>S4,即氮转化速率依次表现为 S2>S3>S4。
2.4 不同肥力土壤胞外酶化学计量学
5 种肥力土壤的 ln(BG+CBH)∶ln(NAG+LAP)、ln(BG+CBH)∶ln AP 和 ln(NAG+LAP)∶ln AP均呈直线正相关关系(图 3)。ln(BG+CBH)∶ln(NAG+LAP)和ln(BG+CBH)∶ln AP均小于1,表明相对于氮、磷来说,土壤养分转化速率受土壤碳转化相关酶活性的限制。ln(NAG+LAP)∶ln AP高于 1,说明相对于氮来说,土壤养分转化速率更受土壤磷转化酶活性的限制。另外,3个图中土壤肥力越高,其比例越接近1,说明低肥力土壤S1受养分限制最为强烈,高肥力土壤S5相比中肥力土壤S2—S4,其所受碳或磷的限制更小,养分比例更均衡。
2.5 不同肥力土壤胞外酶活性与基础理化性状的冗余分析(RDA)
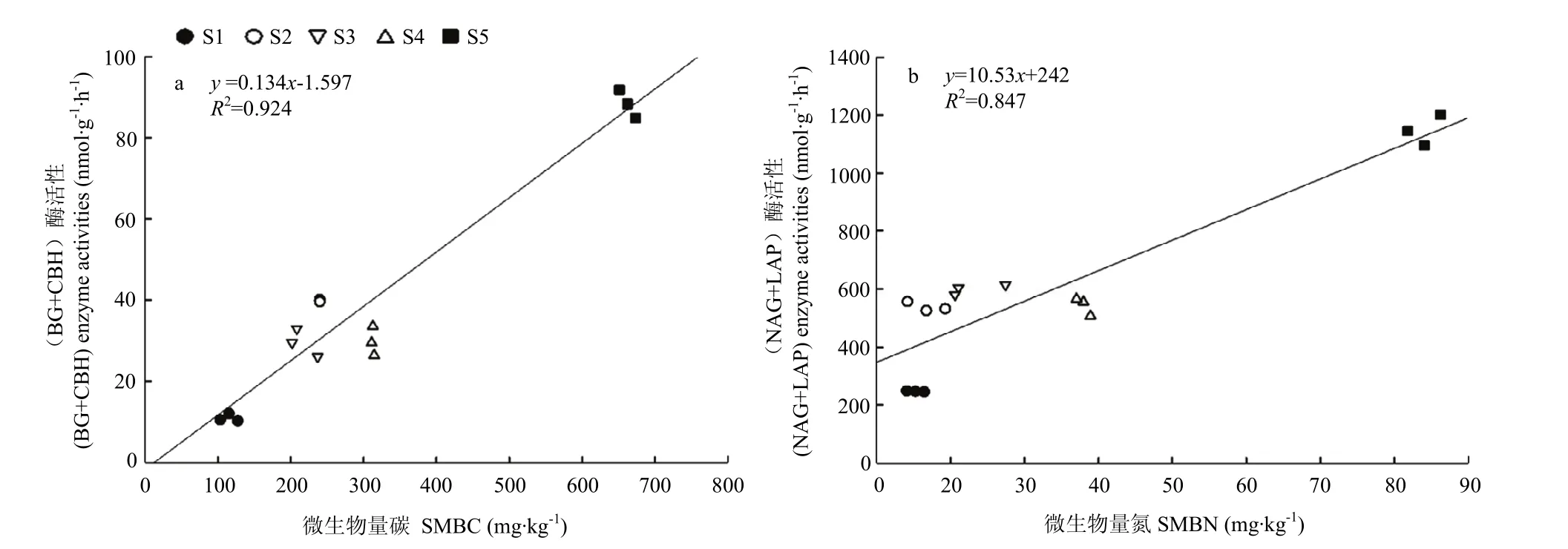
图2 不同肥力土壤碳氮转化相关胞外酶活性与微生物量碳氮的关系Fig. 2 Relationship between extracellular enzyme activities related to carbon and nitrogen transformation and microbial biomass carbon and nitrogen in different fertility soils
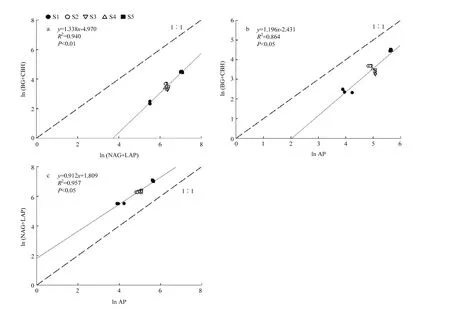
图3 不同肥力土壤胞外酶化学计量学Fig. 3 Stoichiometry of extracellular enzymes in different fertility soils
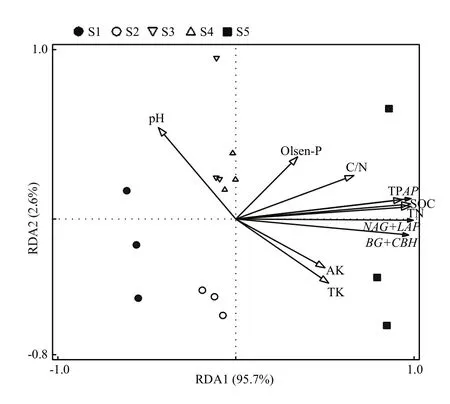
图 4 不同肥力土壤胞外酶活性与基础理化性状的冗余分析Fig. 4 Redundant analysis of extracellular enzyme activity and basic properties of different fertility soils
通过冗余分析(图 4)可以发现碳氮磷转化相关酶活性与基础理化性状的主解释率达到了95.7%,表明胞外酶活性与土壤养分有着非常紧密的联系。碳氮磷转化相关酶活性与 SOC、TN、TP存在着紧密的正相关关系,表明土壤全量碳氮磷养分含量显著影响胞外酶活性,与AP、AK、TK、C/N有着正相关关系,pH与胞外酶活性夹角为钝角,表示pH与胞外酶活性呈负相关关系,这可能是所选的5个肥力土壤的pH相差不大,土壤pH不是影响胞外酶活性的主要因素。
3 讨论
3.1 不同肥力土壤微生物量碳氮差异
本研究中SMBC和SMBN均在高肥力土壤上最高,低肥力土壤上最低。其原因可能是高肥力土壤上作物根系生物量较高且养分库容大,因此有机碳积累量高。有机碳的积累一方面增加了微生物的数量和活性[15],另一方面可促进团聚体结构的形成,改善土壤物理性状,为微生物生长繁殖提供了良好的条件[16-17],进而提高微生物量。但是,也有研究结果发现长期施用无机肥对SMBC和SMBN的影响较小[18],因为作物生长消耗大部分养分且小麦生长状况较差,根系及残留物都较少,长期施用化肥使土壤的C/N比降低,加速了土壤中原有有机碳的分解,导致土壤中积累的有机碳总量较少[19]。但是本研究中高肥力土壤在后期试验收获时作物产量较高(数据尚未发表),微生物量也较高。与化肥相比,很多研究表明,施用有机物料可更大幅度的提高微生物生物量[20-23]。主要的原因是有机物料可以促进作物根系和地上部的生长,增加外源碳的输入,增加土壤微生物数量,增强了微生物的活性。
土壤微生物量碳氮比(SMBC∶SMBN)可表征土壤中微生物群落结构信息,一般来说,真菌的C/N值在7—12之间,细菌在3—6之间[24]。有研究发现尽管施肥显著提高了微生物的总量,但并未明显改变土壤微生物的群落结构[25]。本试验结果中土壤的SMBC∶SMBN在 7.66—12.43之间,除 S2外没有显著差异(表2),说明供试土壤间整体群落结构中细菌真菌比例无显著差异。虽然细菌真菌比例没有差异,但随着土壤肥力的提升,SMBC和SMBN含量显著升高,土壤细菌和真菌含量应该都有所提高,这与之前的研究也相吻合[17],只是其比例没有变化。深入分析不同土壤肥力以及施肥对微生物群落结构,尤其是关键微生物种群的影响,对于阐明养分功能发挥的差异机制具有重要意义,值得进行深入研究。
SMBC与SOC的比值称为微生物熵,表示土壤中的碳转化效率,一定程度上表明了土壤中碳的生物有效性[26],同理,SMBN与TN的比值也代表了土壤中氮的生物可利用性[27]。施氮量的提高可以显著提高土壤微生物熵,这是因为施用氮肥弥补了土壤中氮素的消耗,加大了SMBN的固持,提高了SMBC量,但会加速SOC的分解,从而提高微生物熵[28-29]。本试验中,S1和S5的SMBC/SOC和SMBN/TN均较高,这是因为 S5的施氮量和肥力均较高,因此微生物熵高于S2、S3、S4;而低肥力土壤S1虽然土壤肥力缺乏,但是土壤养分没有作物消耗,微生物可利用碳源较为充足,利用碳源的效率较高[18]。
3.2 不同肥力土壤胞外酶活性差异
土壤酶活性代表微生物的活性,是土壤活性功能最重要的指标,在土壤有机物分解和物质循环中起到重要作用[30]。土壤胞外酶活性在3种肥力土壤上差异显著,均表现为高肥力土壤>中肥力土壤>低肥力土壤(图1)。这是因为高肥力土壤微生物量高,微生物分泌到土壤中的胞外酶数量较多,致使土壤胞外酶活性以及养分转化能力较强。本研究中碳、氮、磷的转化酶活性均在高肥力土壤上高,这是因为一方面高肥力土壤上作物生物量大,残留根茬量高;另一方面施肥通过增加根系分泌物,调节碳氮比,直接或间接增减了碳源,促进微生物活性,加速了土壤碳、氮、磷的循环。马晓霞等[31]发现,对化肥配施有机肥提高了土壤微生物量,也提高了蔗糖酶和脲酶等胞外酶的活性。所以,通过增施有机物来提高地力是促进养分循环、提高作物产量的主要途径。相对于氮磷转化酶活性,本研究中碳转化酶的活性较低,这可能是因为土壤长期施用氮磷肥,大量的肥料投入造成了土壤中氮磷的累积,从而提高了氮磷转化相关酶的活性,而外源碳投入的缺乏,导致碳转化酶的活性偏低。
本研究发现碳氮转化相关酶活性分别与SMBC和SMBN呈直线正相关关系,说明SMBC和SMBN的提高会显著提高碳氮转化酶的活性。SMBC和SMBN每增加 1 mg·kg-1,(BG+CBH)和(NAG+LAP)活性可分别提高 0.134 和 10.53 nmol·g-1·h-1。前人有关酶活的研究有的集中在酶活与土壤养分的关系,如黄土的尿酶、碱性磷酸酶和蛋白酶活性随土壤SOC含量增加而增加[32];有的分析了酶活、SMBC和SMBN随作物生育期的变化,如玉米SMBC和SMBN与土壤酶活性有在玉米生长的前中后期呈现先升高后降低再趋于平稳的趋势[33],但是却很少有人去量化胞外酶活性与SMBC和SMBN的关系。笔者认为量化它们之间的关系是很有必要的,因为酶来源于微生物,酶的活性也影响着微生物的活性,如果将研究放眼于作物的整个生育期,那么酶活性与SMBC和SMBN的关系就表征了整个生育期的养分转化量,对于研究生育期内养分的动态转化和深入理解养分转化的生物机制具有重要意义[34-36]。值得注意的是,微生物量碳氮和胞外酶活性灵敏度非常高,受到土壤类型、季节、测定方法等多种因素影响[12,37-38],其定量关系的获得还需要保证采样、测定环境条件一致并进行多次重复以降低其试验误差,保证其相关方程的准确性,更好的理解养分转化的过程机理。
3.3 不同肥力潮土的酶活计量比
阐明养分元素特定化学计量关系,有助于研究土壤微生物代谢过程和酶活性相互耦合关系及其内在机理,提升对微生物和酶活性的认识以及相关机理的完善[39-41]。全球尺度上,土壤BG∶(NAG + LAP)为1.41,BG∶AP 为 0.62,(NAG+LAP)∶AP 为 0.44[42]。本研究中,BG∶(NAG + LAP)为0.05,远低于全球平均值;BG∶AP为0.21,与全球平均值相比差异不大;(NAG+LAP)∶AP为3.91,远高于全球平均值,表明了5种肥力土壤上氮转化酶活性较强,N分解转化较快,这是由于农田土壤长期施用大量氮肥,氮素积累量较高。
ln(BG+CBH)∶ln(NAG+LAP)和 ln(BG+CBH)∶ln AP等于1被认为是土壤中最适宜于碳氮转化的酶活比例,高于1则说明该土壤受氮/磷源限制,低于1则受碳源限制[43]。本文的供试土壤中,ln(BG+CBH)∶ln(NAG+LAP)和 ln(BG+CBH)∶ln AP均小于1(图3),表明相对于氮、磷,土壤养分转化速率受土壤碳转化效率的限制。长期施用化肥的土壤,由于缺少碳源的补充,碳转化相关酶活性较低,会加剧土壤有机碳的矿化[29],所以应补充碳源,以保证微生物得到充足的能量,提高微生物活性,增强微生物的内稳性,加强其对氮磷养分的转化利用能力。ln(NAG+LAP)∶ln AP高于1,说明相对于氮,土壤养分转化速率受土壤磷转化酶活性的限制。这可能一方面是由于土壤中氮素的大量积累,另一方面磷素大多被固定而有效性低,在实际农业生产中应注意氮肥适量减施和有效磷的供给。本研究还发现,高肥力土壤上的胞外酶活性对数计量比更接近于 1(图3),说明高肥力土壤上土壤酶活计量比更接近适宜值,养分转化利用能力更强,这可能是高肥力土壤高产高效的重要机制之一。
4 结论
提高土壤肥力可提高土壤微生物量碳氮,以及碳、氮、磷转化相关胞外酶活性,但不改变微生物量碳氮比。土壤碳转化酶活性与SMBC、氮转化酶活性与SMBN均呈直线正相关关系,SMBC和SMBN每增加1 mg·kg-1,碳转化相关酶(BG和CBH)活性和氮转化相关酶(NAG和 LAP)活性分别提高 0.134和 10.53 nmol·g-1·h-1。在长期施用化肥的潮土上,碳是微生物活性的首要限制因子。高肥力土壤上碳、氮、磷相关酶活的比例更接近适宜值,说明微生物活性更高,养分的循环转化更快,这可能是高肥力土壤高产高效的主要机制之一。
猜你喜欢
今日农业(2022年14期)2022-11-10 00:27:51
江苏农业科学(2020年2期)2020-04-17 09:53:15
中国化肥信息(2018年7期)2018-08-23 09:12:42
江苏农业科学(2017年10期)2017-07-21 17:00:28
西南农业学报(2016年5期)2016-05-17 05:42:37
江西煤炭科技(2015年1期)2015-11-07 03:06:32
湖北农业科学(2015年17期)2015-10-09 22:07:40
小哥白尼·趣味科学画报(2014年9期)2015-04-07 21:01:12
湖北农业科学(2015年2期)2015-03-20 08:25:42
湖南农业科学(2014年14期)2014-02-27 14:31:52
