
玉米XYLPs基因家族表达模式及其蛋白结构分析
2020-11-14 01:45张洪权熊校勤黄丽霞
浙江农业学报 2020年10期
钟 静,谭 芬,张洪权,熊校勤,黄丽霞
(湖北第二师范学院,植物抗癌活性物质提纯与应用湖北省重点实验室,湖北 武汉 430205)
植物维管组织包括内部的木质部、外部的韧皮部和中间的形成层细胞,它们共同构成植物“骨架”[1]。维管组织在植株正常的生长发育过程中发挥着重要的作用[2],其木质部向上运输根吸收的水分和无机盐,韧皮部向下运输有机物(例如光合作用产生的糖类),从而将植物地上部分的叶、其他器官与地下部分的根联系起来[3-4]。同时,维管组织还具有机械支撑、储藏和传递信息等多种功能[2]。最新研究显示,植物维管组织发育过程受多种内外环境影响、涉及多种信号机制[5]。Motose等[6-7]在2001年首次报道了一类阿拉伯半乳糖蛋白(arabinogalactan proteins,AGPs)在百日菊(ZinniaelegansL.)叶肉细胞导管分化过程中发挥关键作用,并将其命名为木质形成素(xylogen)。随后的研究显示,木质形成素蛋白骨架同时包括AGP结构域和非典型脂转运蛋白(non-specific lipid transfer protein,nsLTP)结构域,因此又被称为木质形成素类AGP(Xylogen-like arabinogalactan protein,XYLP)[8]。拟南芥xylp1/xylp2双突变体叶片中叶脉发育不连续或者存在错误连接,同时叶脉减少;但是,双突变体幼苗胚轴中的维管系统并无显著变化[9]。对水稻XYLP7的研究结果表明,该基因在穗轴生长过程中发挥重要作用[10]。
随着我国经济和社会的快速发展,人们对粮食的需求日益增加。玉米是我国一种重要的粮食作物,其从籽粒到秸秆都具有良好的经济价值。前人的研究表明,XYLPs在植物维管组织发生过程中起着重要作用,但是对其研究主要集中在拟南芥、水稻和百日菊中[6-10]。对玉米等重要的经济作物中该类蛋白的研究十分有限。同时XYLPs的功能是否也涉及植物其他生长发育过程也并不清楚。基于此,本研究对前期从玉米全基因组中鉴定的16个XYLPs蛋白骨架结构、基因的染色体定位和复制情况、表达模式等进行了较为深入的分析。研究结果能够为进一步解析XYLPs在重要的经济作物玉米中的生物学功能提供重要线索。
1 材料与方法
1.1 玉米XYLPs序列获得
参照前期方法获得玉米XYLPs基因家族基因和蛋白序列[11]。
1.2 玉米XYLPs基因染色体定位和复制分析
将蛋白序列提交至maizeGDB数据库(https://www.maizegdb.org)中检索其基因座位,利用MapChart软件绘制染色体定位图[12-13]。在数据库中查找ZmXYLPs基因存在的片段复制关系。当基因对的共线性关系小于500 kb时,认为基因之间存在片段复制关系[8]。当基因对位于同一染色体且之间被最多5个基因所间隔时,认为基因之间存在串联复制关系[10]。
1.3 玉米XYLPs基因表达模式分析
从maizeGDB数据库中下载前人提交的玉米基因芯片(Microarray)表达数据,并从中查找ZmXYLPs的表达数据,其中ZmXYLP2/10/13/16这4个基因未查找到数据。随后从中选择以下组织的Microarray数据进行分析:萌发24 h的种子(24 h-germinating seed,24H-GS)、播种后6 d的胚芽鞘(6 days after sowing-coleoptile,6DAS-C)、播种后6 d的初根(6 days after sowing-primary root,6DAS-PR)、出苗期的整个小苗(vegetative emergence-whole seedling,VE-WS)、一叶/三叶/四叶期的茎和茎顶端分生组织(vegetative 1/3/4-stem and shoot apical meristem,V1-SS)、五叶期的茎尖(V5-shoot tip,V5-ST)、三叶期第一片叶和叶鞘(V3-first leaf and sheath,V3-LS)、三叶期最顶端的叶(V3-topmost leaf,V3-TL)、七叶期处于第二阶段叶的顶部(V7-tip of stage-2 leaf,V7-TSL)、七叶期处于第二阶段叶的基部(V7-base of stage-2 leaf,V7-BSL)、九叶期幼叶(V9-immature leaves,V9-IL)、九叶期第八片叶(V9-eighth leaf,V9-EL)、抽雄期的第十三片叶(vegetative tasseling-thirteenth leaf,VT-TL)、五叶/七叶期第一节间(V5/7-first internode,V5-FI)、九叶期第四节间(V9-fourth internode,V7-FI)、十三叶期幼雄穗(V13-immature tassel,V13-IT)、十八叶期减数分裂期雄穗(V18-meiotic tassel)、十八叶期的幼雌穗(V18-immature cob,V18-IC)、吐丝期授粉前的雌穗(R1-pre-pollination cob,R1-PC)、吐丝期的花丝(R1-silks,R1-S)、吐丝期的花药(R1-anthers,R1-A)、授粉后2/6/10天的种子(2/6/10 days after pollination-whole seed,2/6/10DAP-WS)、授粉后12~22 d的胚乳(12-22 days after pollination-endosperm,12-22DAP-En)、授粉后16~22 d的胚胎(16-22 days after pollination-embryo,16-22DAP-Em)[12]。对查找到的12个ZmXYLPs表达数据利用Cluster和Treeview软件做出热图,并对其进行聚类分析。
1.4 玉米XYLPs蛋白骨架结构分析
将蛋白序列分别提交至SMART(http://smart.embl-heidelberg.de/smart/set_mode.cgi?NORMAL=1)、SignalP4.1(http://www. cbs. dtu. dk/services/SignalP/)和Big-PI Plant Predictor(http://mendel.imp.ac.at/gpi/plant_server.html)检测nsLTP结构域、N端信号肽和C端糖基磷脂酰肌醇锚(glycosylphosphatidylinositol,GPI)[14-16]。手动检测蛋白序列中富含PAST(Hyp/Pro、Ala、Ser和Thr)的AGP结构域[17]。
1.5 玉米XYLPs糖基侧链修饰位点分析
将蛋白序列提交到NetNGlyc 1.0 Server(http://www.cbs.dtu.dk/services/NetNGlyc/)检测N连接糖基侧链修饰位点。参照前人研究手动检测蛋白序列中可能的O连接糖基侧链修饰位点:蛋白质骨架中含有Ala/Ser/Thr-Pro且相邻脯氨酸之间间隔的氨基酸少于11个,则结构中的Pro可被阿拉伯糖和半乳糖修饰;在[(Ala/Ser/Thr)-Pro]2-4结构中的Pro可被阿拉伯糖基侧链修饰位点[18-21]。
2 结果与分析
2.1 ZmXYLPs基因的染色体定位及基因复制
根据从maizeGDB数据库中获得的ZmXYLPs基因精确座位,将16个ZmXYLPs基因标识到玉米染色体上。结果显示,1号、2号、4号、5号、8号、9号和10号染色体上分别分布有2个、4个、1个、2个、1个、4个和1个ZmXYLP基因。第3号、6号和7号染色体无ZmXYLPs的分布(图1)。进一步检测了ZmXYLPs的基因复制情况,结果显示,玉米ZmXYLPs基因家族成员中存在4对片段复制基因对:ZmXYLP6-ZmXYLP8、ZmXYLP8-ZmXYLP7、ZmXYLP7-ZmXYLP9和ZmXYLP9-ZmXYLP10。3对串联复制基因对:ZmXYLP3-ZmXYLP4、ZmXYLP8-ZmXYLP9和ZmXYLP15-ZmXYLP16。
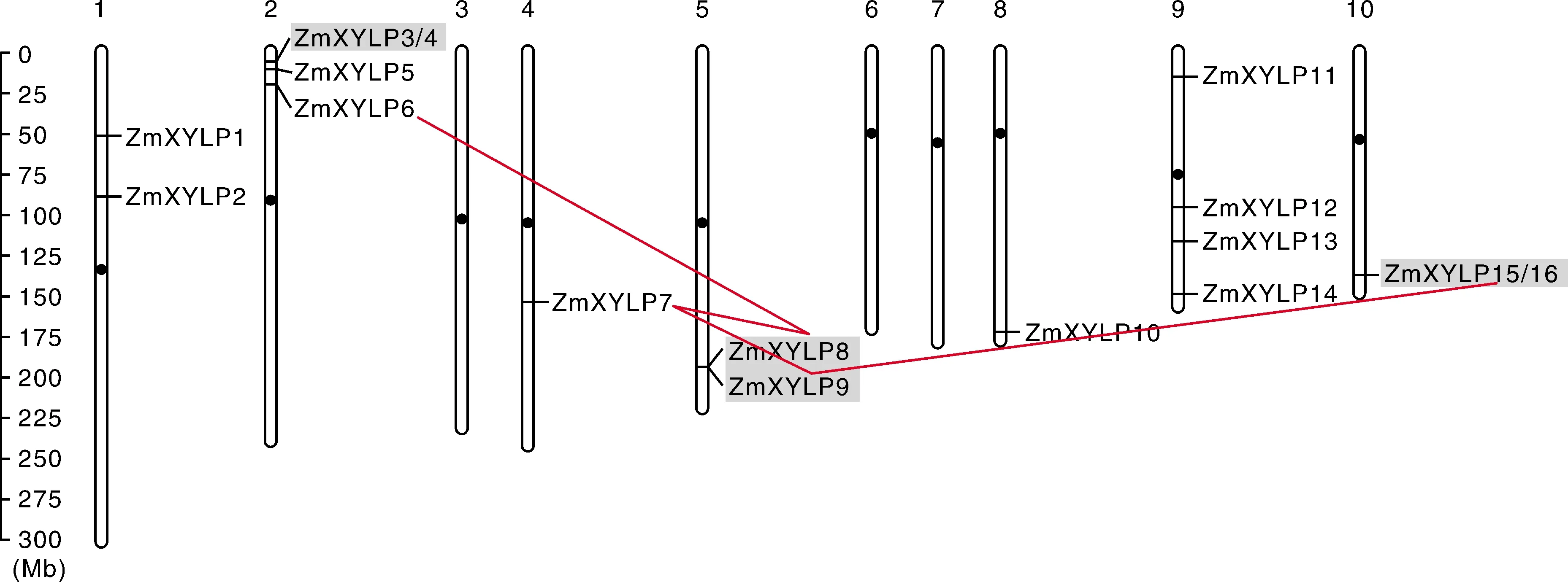
左边为染色体长度标尺,单位为Mb;顶端为染色体号,染色体上的黑色椭圆表示着丝粒位置;红色直线表示基因之间发生了片段复制事件,浅灰色背景表示基因之间发生了串联复制事件。The scale of chromosome length with the unit of Mb was located on the left; the chromosome number was located on the top; the red line indicated that there was a segment replication event between genes, and the light grey background indicated that there was a series replication event between genes.图1 ZmXYLPs基因的染色体定位和基因复制关系Fig.1 Chromosomal localization and gene duplication of ZmXYLP genes
2.2 ZmXYLPs基因表达模式分析
进一步对ZmXYLPs在37个玉米不同发育时期和不同组织中的表达模式进行了分析。结果表明,ZmXYLPs基因在玉米不同发育时期的组织和器官中的表达存在明显差异(图2、图3)。ZmXYLP1/3/5在所有组织中的表达量均很低(图2、图3)。而ZmXYLP7和ZmXYLP8则特异性的在播种后6 d的初根中表达,但是ZmXYLP8表达水平明显高于ZmXYLP7。ZmXYLP9在幼叶中高量表达,在七叶期叶的基部和十三叶期的幼雄穗中量表达(图2、图3)。ZmXYLP6/14/15表达模式较为相似,在大多数营养组织和少部分生殖器官中都有表达,但是ZmXYLP14的表达水平普遍明显低于ZmXYLP6和ZmXYLP615(图2、图3)。在三叶期最顶端的叶片、七叶期叶的基部和九叶期的幼叶中ZmXYLP6和ZmXYLP615都呈现高表达(图2)。五叶期的茎尖、十三叶期的幼雄穗、十八叶期减数分裂的雄穗和授粉后2 d的种子中ZmXYLP6表达水平高于ZmXYLP15(图2、图3)。此外,在播种后6 d的胚芽鞘中也检测到ZmXYLP15的表达(图2)。ZmXYLP4表达量普遍较低,且主要在生殖器官中低表达。ZmXYLP12在大多数组织器官中呈现泛表达模式,且表达量均较高(图2、图3)。ZmXYLP11的表达模式与其他蛋白均不相同,仅在生殖器官中表达,尤其在胚胎中的表达量较高(图3)。

顶端数字为对应的ZmXYLPs;底部色彩刻度标尺显示信号值的大小,从黑到红表示信号值从低到高。下同。The name of ZmXYLPs was located on the top; the bottom color scale corresponded to size of the signal value, from black to red indicated that the signal value was from low to high. The same as below.图2 ZmXYLPs在营养生长时期的表达模式Fig.2 The expression pattern of ZmXYLPs in vegetative growth stage
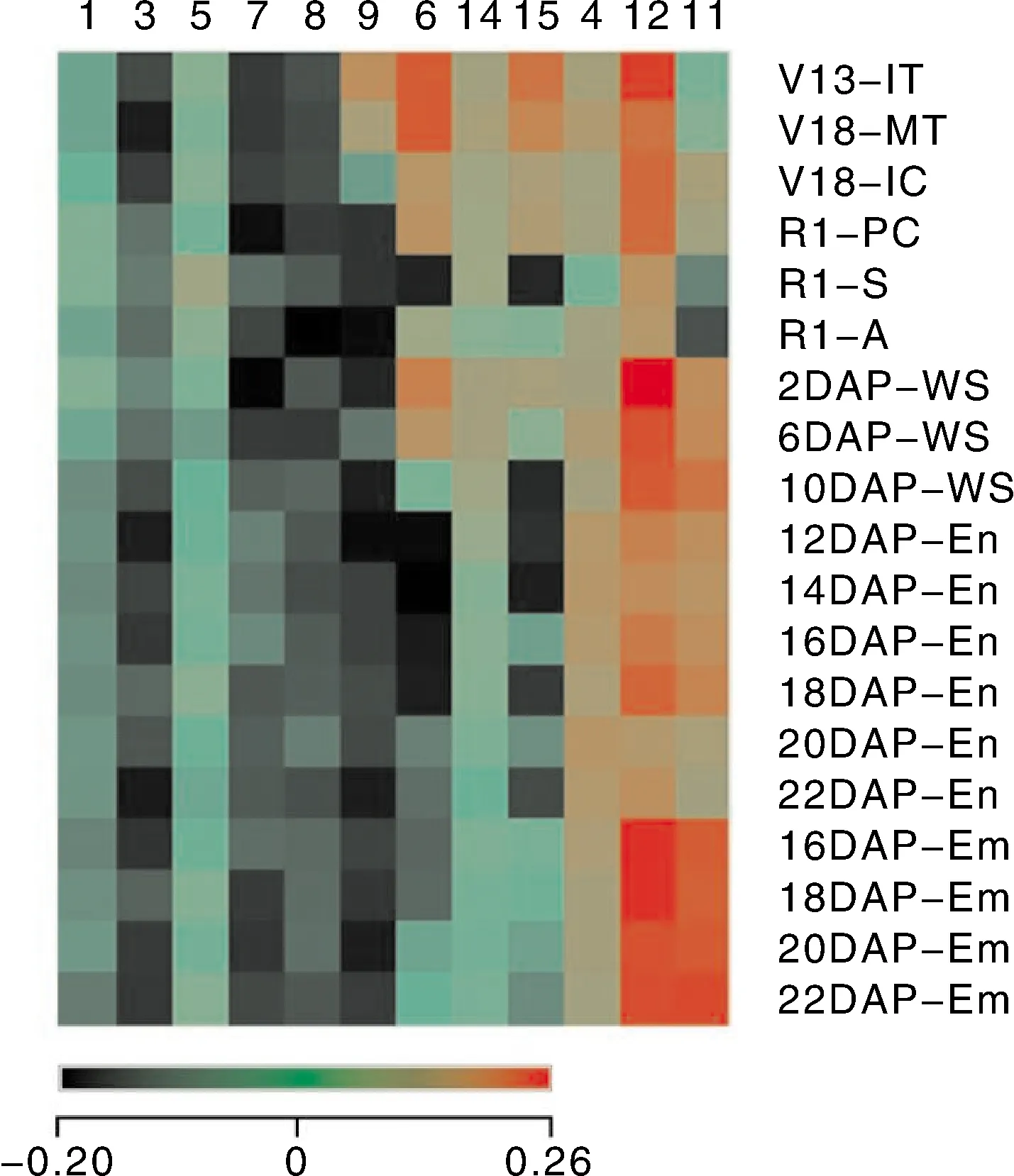
图3 ZmXYLPs在生殖生长时期的表达模式Fig.3 The expression pattern of ZmXYLPs in reproductive development stage
2.3 ZmXYLPs蛋白结构分析
对16个ZmXYLPs的蛋白序列提交到在线网站预测,结果显示,所有蛋白均含有N端信号肽,信号肽长度在21~29个氨基酸之间,其中ZmXYLP1、ZmXYLP10和ZmXYLP14具有C端的GPI锚(表1)。所有蛋白都具有1个nsLTP结构域。ZmXYLP1和ZmXYLP14含有2个AGPs结构域,其余均仅含有1个AGPs结构域(表1)。
根据N端信号肽、GPI锚、nsLTP和AGP结构域位置,可将16个ZmXYLPs分为两类(表1、图4)。第一类中,nsLTP结构域位于中间,左右两侧分别有2个AGP结构域,C端有一个GPI锚。除ZmXYLP1、ZmXYLP10和ZmXYLP14三个以外,其余ZmXYLPs均属于第二类,nsLTP结构域位于AGP结构域之后,无C端GPI锚(图4)。

绿色方块代表N端信号肽,红色方块代表AGP结构域,蓝色方块代表nsLTP结构域,紫色方块代表C端GPI锚。The green boxes represented the N-terminal signal peptide, the red boxes represented the AGP domain, the blue boxes represented the nsLTP domain and the purple boxes represented the GPI anchor signal.图4 ZmXYLPs蛋白结构示意图Fig.4 The sketch map of protein structure of ZmXYLPs
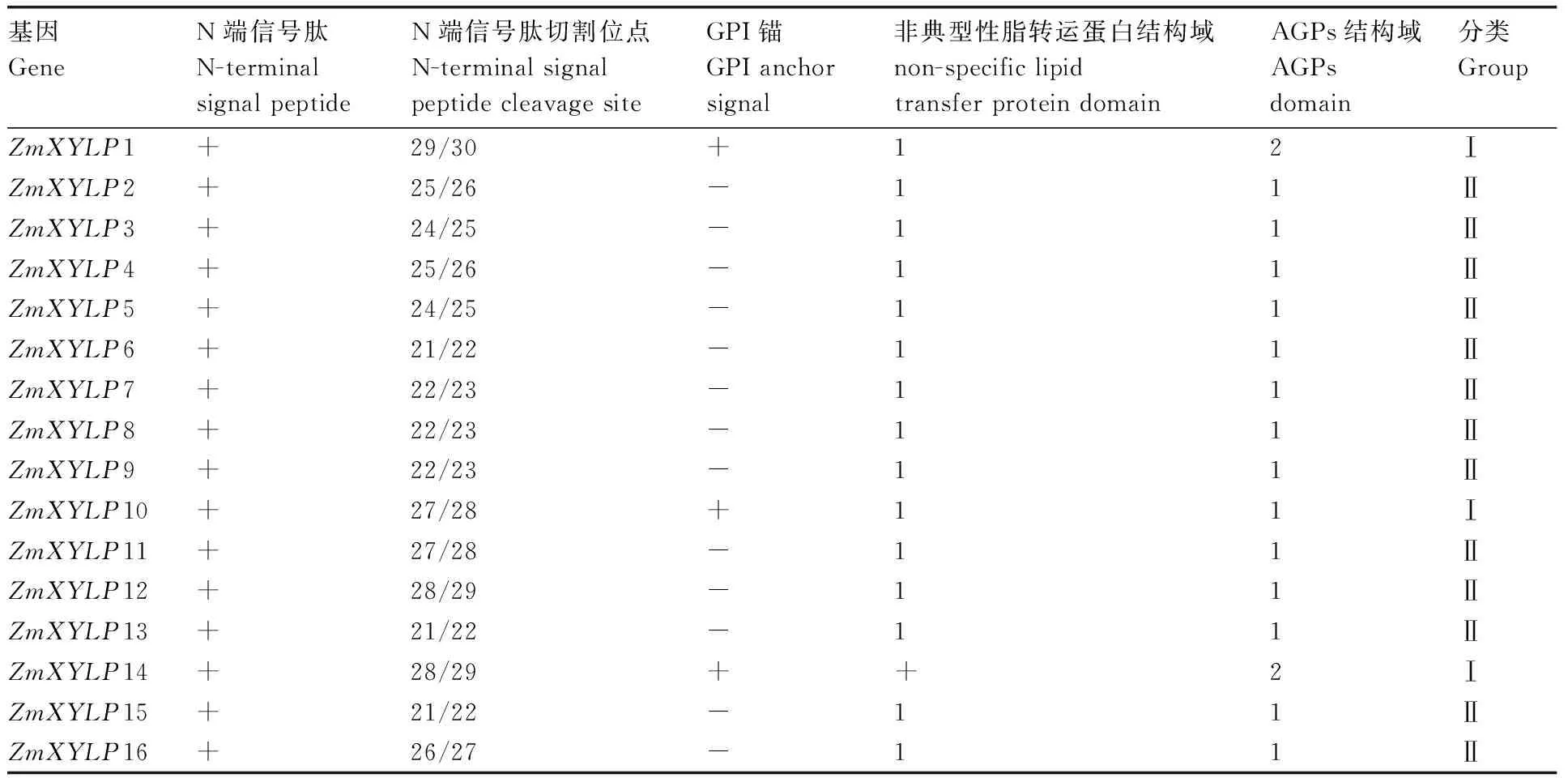
表1 玉米XYLPs蛋白结构分析Table 1 Protein structure analysis of maize XYLPs
2.4 ZmXYLPs蛋白糖基侧链修饰位点分析
对ZmXYLPs蛋白骨架糖基侧链修饰位点的检测结果显示,ZmXYLP10、ZmXYLP13和ZmXYLP14各有一个位于nsLTP或AGPs结构域的N连接糖修饰位点。所有ZmXYLPs均至少有1个O连接糖修饰位点,但是仅ZmXYLP6、ZmXYLP11、ZmXYL12和ZmXYLP15这4个蛋白中有(A/S/T)-P3结构(表2)。ZmXYLP11中O连接糖修饰位点多达19个,主要包括(A/S/T)-P和(A/S/T)-P2两种结构(表2)。
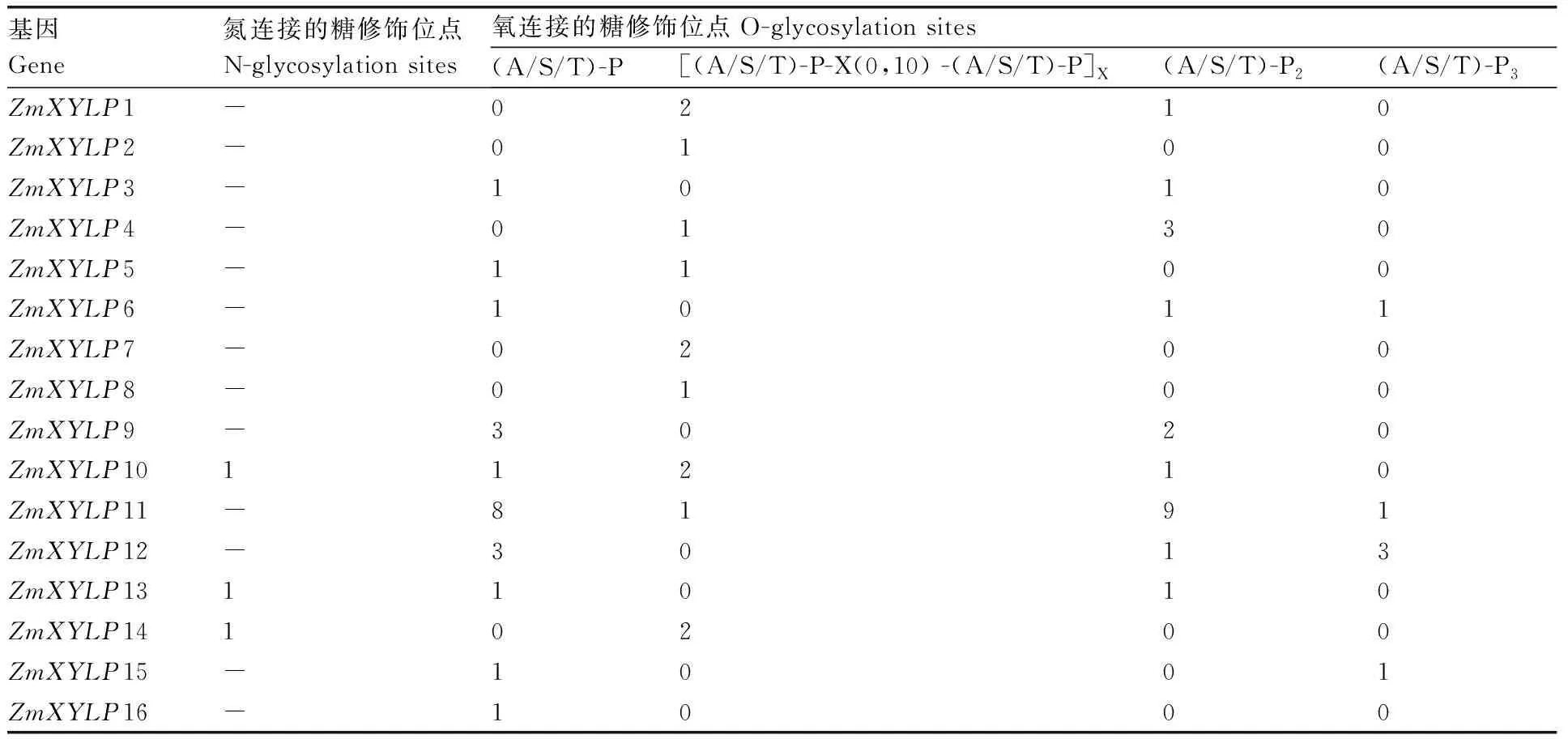
表2 玉米XYLPs蛋白糖基修饰位点分析Table 2 Analysis of glycosyl modification sites of maize XYLPs
3 讨论
本研究采用生物信息学方法对16个ZmXYLPs基因进行了染色体定位、表达和蛋白结构分析。结果显示,玉米第3号、6号和7号这3条染色体上没有ZmXYLPs基因的分布,但是2号和9号染色体上分别聚集有4个ZmXYLPs基因(图1)。这说明ZmXYLPs基因家族成员在染色体上的分布具有一定的偏好性。进一步的检测结果显示,ZmXYLPs基因家族中的5个成员均由片段复制而来,且形成4对片段复制基因对;同时有4个成员由串联复制而来,形成2对串联复制基因对。其中ZmXYLP15同时涉及片段复制和串联复制事件(图1)。共有8个(50%)ZmXYLPs涉及基因复制,说明基因复制事件在ZmXYLPs家族的扩张和进化中扮演了极其重要的角色,且片段复制和串联复制对该家族膨胀的贡献基本相同。
利用基因芯片(microarray)数据对这些基因表达模式的分析结果显示,ZmXYLP1/3/5在几乎所有组织中均呈现低表达,暗示着这3个基因可能存在功能冗余性(图2、图3)。ZmXYLP7和ZmXYLP8都特异性地仅在初根中表达,尽管两个基因之间存在片段复制关系,但是其各自的表达量有明显差异。ZmXYLP7和ZmXYLP9也是片段复制基因对,但是ZmXYLP9并未在初根中有表达且其表达的部位要多于ZmXYLP7。同样的情况也出现在ZmXYLP6和ZmXYLP8片段复制基因对以及ZmXYLP8和ZmXYLP9串联复制基因对中(图2、图3)。在ZmXYLP9和ZmXYLP15片段复制基因对中,ZmXYLP15的表达范围在覆盖ZmXYLP9的基础上有较大的扩大,且表达量有明显提高(图2、图3)。以往的研究表明,XYLPs的生物学功能可能涉及植物维管组织发生[6-7,9-10]。但是本研究结果显示,有7个ZmXYLPs基因在生殖器官中也呈现高量表达模式(图3)。其中ZmXYLP12呈泛表达模式(除九叶期第八片叶以外),且表达量普遍较高。而ZmXYLP11则仅在生殖器官中表达,尤其在授粉后16~22 d的胚胎中高量表达(图3)。此结果暗示着基因复制事件不仅扩大了该基因家族成员的数目,同时还可能导致基因表达部位的扩张,从而产生新的生物学功能。
进一步对ZmXYLPs蛋白数据进行了分析。与前人研究一致的是,ZmXYLPs蛋白序列较为保守,均含有N端信号肽和一段nsLTP结构域(表1)。但是16个ZmXYLPs蛋白中仅有ZmXYLP1/10/14三个蛋白含有被nsLTP结构域隔开的两段AGP结构域和C端GPI锚。其余13个蛋白均只含有一段AGP结构域且无C端GPI锚,并且nsLTP结构域均位于蛋白C末端(图4)。而前人报道的拟南芥13个AtXYLPs均含有C端GPI锚,21个水稻OsXYLPs中仅1个不含有C端GPI锚。同时在拟南芥和水稻XYLPs中,大多数蛋白含有两段AGP结构域[10,22]。这说明XYLPs蛋白可能存在一定的种属特异性。XYLPs是一类嵌合型的AGPs,AGPs是一类富含阿拉伯糖和半乳糖的糖蛋白。因此,本研究对ZmXYLPs蛋白的糖基侧链修饰位点进行了预测。仅3个ZmXYLPs蛋白存在N连接的糖修饰位点。其余蛋白糖修饰位点均为O连接,多数ZmXYLPs蛋白O连接糖修饰位点数目为1~5个。此外,O连接糖修饰位点的氨基酸组成也存在差异,具有(A/S/T)-P3结构的ZmXYLPs较少(表2)。进一步的研究发现,ZmXYLP1具有3个氧连接的糖修饰位点,而ZmXYLP14具有1个氮连接的糖修饰位点和2个氧连接的糖修饰位点。而这两个基因的表达模式也存在明显差异(图2和图3)。ZmXYLP7和ZmXYLP8具有同一类氧连接的糖修饰位点,但是数量存在差异。同时两者的表达部位也相同,但是表达量不同。虽然ZmXYLP9与ZmXYLP7/8同属第二类ZmXYLP,但是其氧连接的糖修饰位点和数量明显多于后两者,而ZmXYLP9的表达部位也多于后两者。ZmXYLP6/15与ZmXYLP7/8类似,具有的氧连接糖修饰位点多的ZmXYLP6表达水平高于位点少的ZmXYLP15。ZmXYLP11具有多达19个糖修饰位点,ZmXYLP12的糖修饰位点次于ZmXYLP11。这暗示着这两个蛋白的糖基化程度可能远高于其他蛋白,其总蛋白的分子结构也可能更加复杂。联系这两个蛋白的表达模式(图2、图3),上述结果推测ZmXYLPs蛋白糖基侧链对于该类蛋白的功能可能起着决定作用。本研究结果表明,ZmXYLPs可能并不仅仅在玉米维管组织发生过程中发挥作用,部分基因功能可能还涉及玉米的生殖发育过程。本研究为后继进一步通过功能研究确定ZmXYLPs基因家族成员的具体生物学功能提供重要线索。
猜你喜欢
分子催化(2022年1期)2022-11-02
湖北农业科学(2022年11期)2022-07-18
烟草科技(2021年6期)2021-06-24
实用肿瘤学杂志(2020年4期)2020-12-08
科学之谜(2019年3期)2019-03-28
生物学教学(2018年4期)2018-11-29
电脑知识与技术(2018年19期)2018-11-01
科学之谜(2018年8期)2018-09-29
中学生理科应试(2016年4期)2016-11-19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
