
生物多样性与生态系统服务情景模拟研究进展
2020-11-13 01:02刘焱序于丹丹傅伯杰曹铭昌陈吉星
生态学报 2020年17期
刘焱序,于丹丹,傅伯杰,曹铭昌,陈吉星
1 北京师范大学地理科学学部,地表过程与资源生态国家重点实验室,北京 100875 2 生态环境部南京环境科学研究所,南京 210042 3 首都师范大学资源环境与旅游学院,北京 100048
2019年联合国“生物多样性和生态系统服务政府间科学政策平台”(Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services,IPBES)发布《全球生物多样性与生态系统服务评估报告》,指出当前全球正面临着大自然衰退“史无前例”和物种灭绝率“加速”的局面,保护和恢复大自然需要“变革性改变”[1]。生物多样性与生态系统服务在概念上存在顺承关系,而在机理上探讨二者关系实质是剖析生态系统中物种数量、物种质量、物种间亲缘关系、以及生物与环境相互作用等如何影响生态系统服务的问题[2]。二者在模拟中解决的科学问题不同、模型相互独立,但输入变量有相似性,模拟得到的情景共同服务于生态系统保护决策支持。
具体而言,作为保护和恢复大自然的重要科学依据,生物多样性与生态系统服务情景模拟是指对未来生物多样性与生态系统服务变化轨迹的定量估计,一般利用对未来气候、土地利用等直接驱动力的情景数据集进行建模实现[3- 4]。尽管大量模型所输出的全球情景表明21世纪全球生物多样性与生态系统服务情景将继续下降,但情景之间差异较大,定量结果存在着高度不确定性,影响了未来生物多样性保护和恢复的变革性改变方案制定[3- 5]。揭示生物多样性与生态系统服务的直接与间接驱动机理,提升模型的预测能力以提供信度较高的未来情景,成为当前生态系统保护和恢复科学决策的重点和难点。
面对情景中众多的气候变化与社会经济预测数据源、诸多的统计算法或机理模型、跨时空的尺度和描述对象、本地化的保护目标和决策途径,亟待对当前生物多样性与生态系统服务情景分析研究进行梳理,进一步理清各类模型的应用潜力、生物多样性与生态系统服务预测的合理性、生态系统保护决策的可行性,从而在方法论层面为我国生态系统保护和恢复目标及方案的制定提供科学支撑[6-8]。基于此,本研究拟以统计模型和过程模型为主对比生物多样性相关预测模型,梳理生物多样性以及生态系统服务预测情景的核心观点,讨论基于情景模拟的生态系统保护决策支持途径,以期服务于我国生物多样性与生态系统服务情景分析研究的发展,为生态系统保护和恢复相关规划管理实践提供思路借鉴。
1 情景分析模型研究进展
模型是对系统简化和理想化的理解,当前研究中的生物多样性和生态系统服务模型一般属于统计模型或过程模型[7-9]。其中对物种分布的模拟大多属于统计模型,或称关联模型;少数研究采用过程模型,或称机理模型,刻画物种在生理层面的物理化学条件,但由于参数要求高其时空尺度相对受限[10-11]。在生态系统尺度上,气候变化和人类活动的宏观驱动作用更加明显,在植被分布、生产力与碳循环等研究中一些过程模型也取得了应用[12-13]。此外,生物多样性模拟结果往往是生态系统服务模型的重要输入参数,二者如能有效衔接将加强生物多样性模拟结果对人类社会的现实意义[7-8]。据此,本文重点关注以数理统计算法为主的物种分布模型,气候变化和区域人类活动下生态系统预测模型,并简要对基于生态过程与环境指标的生态系统服务评估模型进行论述。
1.1 以数理统计算法为主的物种分布模型
在物种灭绝、物种丰度、物种分布和遗传多样性四大主要生物多样性变化模拟指标中,物种分布的变化与前两项指标高度相关,并涉及气候等较为宏观的空间指标,因而在生物多样性情景分析中得以广泛应用[14]。一些研究采取了统计模型和过程模型相互校正的预测途径[15],但高精度数据的缺失是制约过程模型应用的重要障碍。Urban等综述百余个气候变化下生物模型应用的研究案例,发现涉及生物学机理(过程)模型的研究仅占23%,基于数理统计的建模方式依然是生物多样性情景分析的主流方式[16]。
在众多物种分布模型中,早期的BIOCLIM、HABITAT、DOMAIN等多基于规则判定,输出值为0或1布尔值,常用总体精度或kappa系数进行检验;然而考虑到空间环境信息的复杂性,这种二分法判定规则的不确定性相对较高[14]。基于机器学习思想的概率分布模型近年来取得了广泛应用,其中最大熵模型Maxent是应用最为广泛的物种分布模型,人工神经网络(Artificial Neural Network,ANN)、随机森林(Random Forest,RF)、支持向量机(Support Vector Machine,SVM)等也快速成为物种分布建模的主流算法[10]。此类算法通常将物种的适宜空间表示为0到1连续分布,可以通过RMSE(Root Mean Squared Error)、ROC(Receiver Operating Characteristic)曲线、R2等进行检验[17-18]。然而,尽管机器学习可以在大数据支持下发挥较强的空间信息挖掘与评估能力,但常见的数理统计算法对物种相互作用与物种迁移能力的表征仍有不足[19]。例如对于海洋物种,由于环境背景信息相对有限,往往会嵌入食物网模型提升模拟能力[20]。再如一些地形复杂地区,无法获得精确的预测数据或物种数据,引入专家知识对模型输出进行批判性评估,有助于识别输出结果偏离现实的具体微生境因素,从而明确模型优化的具体参数[21]。在近年研究中,多种数理统计算法结果的相互校正和生物过程的进一步融入已成为物种分布模型的重要应用趋势。
同时,尺度是影响物种分布模型精度和稳定性的关键因素。复杂的模型在较细的分辨率下可能更准确,而简单的模型在更大的尺度上可能提供简洁的通用解决方案;但复杂模型的开发仍然是必要的,其有助于确定精确性和通用性之间的最佳权衡[22]。Ferrier和 Guisan总结了群落尺度上生物多样性模型的预测能力,认为在不同建模策略下对群落组分的模拟能力不存在全部最优,而是此消彼长的关系[23]。显然,从群落尺度扩大到生态系统尺度,对各种群落的模拟能力的变化也会存在类似的规律,很难存在绝对最优的模拟策略。这也引申出另一科学问题:如何研发一个符合基本预测精度和稳定性需求的物种分布模型。对此,Araújo等基于对400余个生物多样性模型研究的评价,使用“不足”“青铜”“白银”“黄金”等形象词汇划分了模型建立、模型评估、预测变量、响应变量的等级,提出了生物多样性建模的最佳实践标准,并鼓励管理者、科学家和用户对框架内容的更新[24]。这一标准体系的提出无疑将对物种分布模型的研发和改进提供更明确的依据。
1.2 面向气候变化和人类活动的生态系统预测模型
气候变化是驱动宏观尺度上生物多样性变化的一项主导因素,气候模式输出的气候变化情景数据集也被众多陆地或海洋生态系统的生物多样性预测研究所使用[25-26]。Bellard等将可用于大尺度生态系统研究的生物多样性模型分成2大类5小类,在生物多样性范围模型大类中包括了生物气候包络模型、动态植被模型,在物种损失模型大类中包括了物种-面积关系、IUCN(International Union for Conservation of Nature,世界自然保护联盟)状态、驱动-反映关系三种模型[27]。显然,物种损失模型大类在较大空间尺度上未必能有效描述生物物理过程,而生物多样性范围模型可包含更多空间显式的过程模型[28]。值得注意的是,在生态系统或景观尺度上,自下而上从物种出发评估生物多样性往往能更有效的发挥过程模型的优势,但切实服务于大尺度多物种的过程模型毕竟有限,模型对比和交互验证的难度远高于以数理统计算法为主的物种分布模型。
土地利用变化是人类活动在空间上的直观反映,也是生物多样性变化的另一项主导因素,而未来土地利用变化情景又与气候变化直接相关。对应于4种RCPs(Representative Concentration Pathways,典型浓度路径)情景,大尺度生物多样性建模中的土地利用变化数据也分为了IMAGE 2.6、MiniCAM 4.5、AIM 6.0、MESSAGE 8.5。其中,IMAGE模型假定了城市范围不变,农业从发达国家转移到发展中国家,生物燃料种植面积大幅增加;MiniCAM模型假定了城市范围不变,作物产量的增加、农业效率的提高和饮食结构的改变导致了农田和牧场面积的减少,生物燃料种植面积小幅增加,碳价格导致原生林的保护和次生林的扩张;AIM模型假定了人口增长下城市面积增加,耕地面积增加以满足粮食需求,草场面积锐减;MESSAGE模型则假定了人口增长下城市面积增加,耕地面积和牧场面积迅速增加,生物燃料种植面积小幅增加[29]。结果显示,高强度人类活动下的未来农用地扩张成为全球生物多样性的重要威胁源。
GLOBIO是一项经典的评估人类活动对生物多样性影响的工具,目前已更新至3.5版。该模型将生物多样性的驱动力分解为植被覆盖、土地利用、气候变化、大气氮沉降、基础设施和生境破碎度,评价指标包括物种丰富度和生态系统范围[30-31]。GLOBIO模型基于指标的因果关系建构,并不涉及具体物种的生理过程,因而具有较好的拓展性,可用于评估陆地、淡水和海洋生态系统,其输入参数可直接由IMAGE模型生成,因而参数结构的匹配效果相对更好[31]。以GLOBIO淡水生态系统模型为例,其组织结构如图1所示[32]。而除了GLOBIO所代表的基于因果关系的模型研发思路外,一些基于碳氮循环与生产力的生态系统过程模型可以更有效的提供物种生理参数[33-34],另一些融合社会经济过程的模型则可以更有效的表达社会发展的生物质占用[35]。但值得注意,虽然GLOBIO等模型的养分模块、水文模块可以用于生态系统服务评估,但目前养分和水文模块的输出参量实际上是作为生物多样性模拟的驱动因素,并未实现与生态系统服务模型中营养物传输、碳固定、产水、土壤保持等模块的定量关联,生物多样性与生态系统服务情景在模型输出结果上依然未能有效衔接。
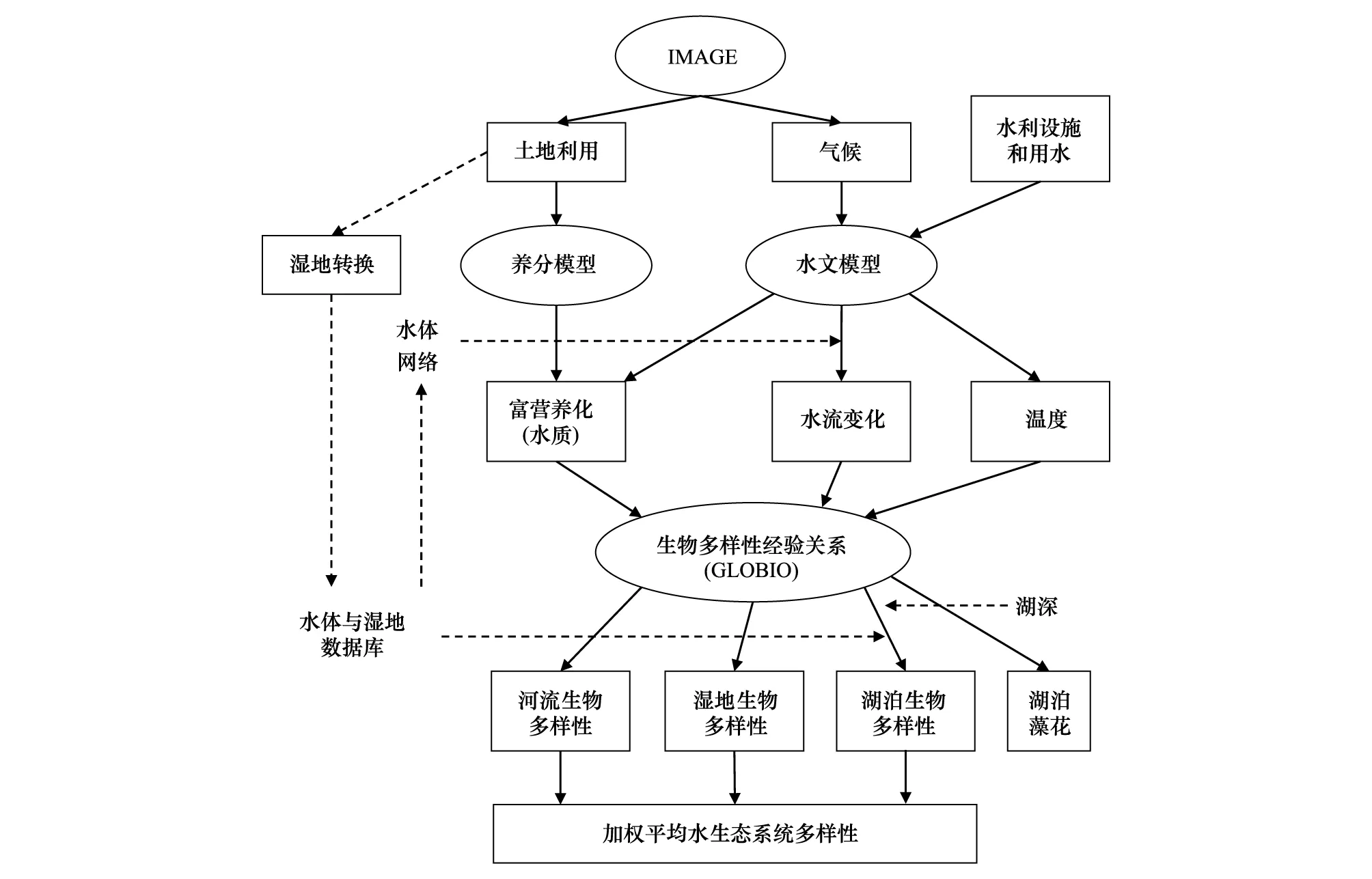
图1 GLOBIO淡水生态系统模型结构[30]Fig.1 Model structure of GLOBIO-Aquatic
1.3 基于生态过程与环境指标的生态系统服务评估模型

图2 SAORES模型结构[39]Fig.2 Model structure of SAORES
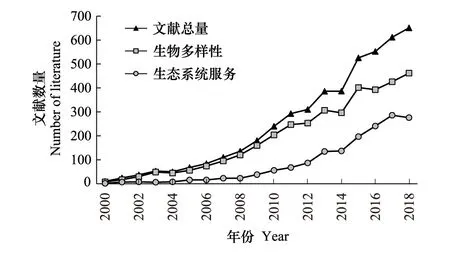
图3 生物多样性和生态系统服务情景建模文献数 Fig.3 Number of model and scenario studies on biodiversity or ecosystem service
生物多样性变化所导致的生态系统结构和过程变化直接影响着生态系统服务,然而针对生物多样性—生态系统功能—生态系统服务—人类福祉的经典级联框架,现有的评估指标体系和模型方法很难准确刻画其级联路径[36]。大部分生态系统服务评估模型是针对景观尺度、流域尺度和区域尺度研发的,因此往往较少涉及相关物种指标,多选用生态过程与环境指标作为输入参数[7]。由于生态系统服务是对生态系统功能的一种人类价值表述形式,因而生态系统服务评估的核心是对生态系统功能的量化和空间模拟,其中土地利用是生态系统服务评估的关键环境指标,尤适于景观尺度的生态系统服务评估建模[37]。其他面向气候变化和人类活动的生态系统预测模型中常用参数也大多在生态系统服务评估模型中涉及,如气温、降水、地形、土壤、生产力、碳和氮磷等营养物[36]。
关于生态系统服务模型的指标特征,现有国内研究已进行了较为详细的总结归纳[7]。其中功能最为强大的是当前一直在不断改进的InVEST模型和ARIES模型。InVEST模型内部模块大多涉及指标的专家赋权,也有部分模块直接基于水文过程,其评估生态系统服务种类多、输入参数简单、操作简便,因而成为生态系统服务快速评估的最常用模型;ARIES模型嵌入了贝叶斯概率模型、过程模型和主体模型,以刻画生态系统服务流动为目标,可以表征生态系统服务供给、需求和价值,具有广泛的决策应用前景[38]。此外,我国学者自主研发的SAORES模型也基于生态过程与环境指标(图2),依托生态系统服务权衡关系进行土地利用的空间权衡优化,在生态空间规划领域也独具优势[39]。但当前生态系统服务建模中对物种数量、质量、格局、多样性的刻画依然较为简单,尤其缺乏生物多样性组分间相互作用的关系嵌入模型[2],致使当前生态系统服务模型对生物多样性形成机理的表达相对不足。
2 生物多样性与生态系统服务情景预测核心观点
依托RCPs等全球气候变化和土地利用情景,以及一些局地土地利用预测和政策假设,生物多样性和生态系统服务情景建模案例近年来快速增加。在Web of Science核心合集中设置“主题(biodiversity or ecosystem service) AND主题(model) AND主题(scenario)”检索,得到2000—2018年出版文献4708篇,其中涉及生物多样性主题3643篇,涉及生态系统服务主题1630篇,如图3所示。考虑到文献众多、案例冗杂,本研究仅撷取近十年来十余篇影响力较大的文章梳理核心观点,以窥见生物多样性与生态系统服务空间分布预测近今研究前沿。
2.1 生物多样性情景预测
在2010年以来生物多样性预测研究中,Bellard等发表的“气候变化对未来生物多样性的影响”一文取得了极大影响力。作者在全球和次大陆尺度的大量生物多样性模型预测发现,气候变化的生物多样性后果是令人担忧的,甚至有可能成为地球上的第六次物种大灭绝[27]。但Bellard等也提到了不同模型结构导致的巨大不确定性,物种的响应可能被高估而驱动因子的协同作用可能被低估[27],从而导致结果将随评价对象、评价模型和评价区域会发生变化。例如研究者基于空间显式的过程模型iLand发现,气候变化下典型温带森林景观的膜翅目、软体动物和食蚜蝇科多样性预计将增加两倍以上[28]。针对模型间存在的巨大差异,Elith等在影响力很大的“模拟地理范围变化物种的艺术”一文中认为,模型的一致性并不能保证正确性,因为所有模型可能以同样方式出错;并以澳大利亚海蟾蜍分布为例,证明了添加机理模型相关信息的必要性[40]。总体上,生物多样性模拟预测主流观点为:不加控制的气候和土地利用变化将增加物种灭绝风险,但此类风险的程度仍是不确定的[2]。
全球气候模式产品的使用为量化生物多样性对未来气候变化的响应提供了重要数据源。Hickler等发现气候变化下2085年欧洲31%—42%的面积将被不同的植被所覆盖,在远期平衡态下76%—80%的欧洲国土可能存在另一种潜在自然植被[41]。Pearson等预测2050年北极木本植物将增加多达52%,植被变化对气候变化存在正反馈,从而可能导致更大程度的变暖[42]。Hazen等发现未来气候变化下某些海洋顶级捕食者的核心栖息地将发生高达35%的变化,北太平洋的生物多样性将大量向北迁移[43]。与气候变化产品对应,大尺度土地利用变化对生物多样性的影响也是可以基于已有模式产品量化的。Newbold等基于2100年土地利用情景发现:在一切照常情景下,样本内物种丰富度将比2005年进一步下降3.4%,损失主要集中在经济贫困国家;而强有力的减缓碳排放措施将有助于生物多样性平均提升1.9%,且与国家社会经济地位的关系相对不紧密[29]。总体就预测结果的尺度而言,气候变化所驱动的生物多样性改变是区域尺度的,而土地利用变化的驱动结果具有明显的局地差异。
2.2 生态系统服务情景预测
虽然生态系统服务预测的参数输入和模型规则与生物多样性模型不完全一致,但生物多样性预测可以为生态系统服务预测提供生态学基础。McPherson等基于洛杉矶“百万树倡议”背景,评估沿海和内陆气候区不同树木死亡率情景下的生态效益和货币价值,评估结果发现35年中100万棵树的收益分别是高死亡率情景13.3亿美元和低死亡率情景19.5亿美元;总体效益的81%是美学/其他方面,8%是减少雨水径流,6%是节能,4%是改善空气质量,不到1%是减少大气碳含量[44]。Nelson等对全球城市土地和农田的区域变化在空间格网中进行了分配,发现耕地扩张规模和格局的情景差异导致了作物生产、物种栖息地供给以及生物量碳储存等重要生态系统服务之间的不同权衡,每卡路里作物生产排放碳在5.2g至13.7g之间[45]。Chaplin-Kramer等在巴西的区域案例中得出类似结论,在农业扩张情景下生物多样性和碳储量的下降幅度将相差2—4倍,从森林内部发生变化比从边缘向森林扩张的影响更为明显[46]。Peres等同样从亚马逊森林的生物多样性出发,发现不同狩猎情景下大型果食性动物的灭绝将导致平均2.5%—5.8%的地上生物量降低[47]。同时,针对海洋和海岸带生态系统,Arkema等开发模型量化了珊瑚、红树林和海草提供的生态系统服务,发现存在既减少对沿海栖息地的影响又增加龙虾捕捞收入的共赢情景[48]。显然,对生物多样性和物种栖息地参数规则设置本身的科学性是上述生态系统服务合理预测的前提。
生态系统服务预测与生物多样性预测的重要区别在于,生态系统服务模型更加关注人类的空间决策及其导致的社会效应。土地利用作为一种重要的生态系统服务管理决策的空间载体,是陆地生态系统服务预测中的核心驱动变量。Lawler等预测发现美国潜在的高粮食需求将导致农田快速扩张,致使碳储量、木材产量、粮食产量增加,但导致25%所模拟物种的栖息地减少超过10%[49]。Polasky等对明尼苏达州的土地管理情景模拟表明,农业大规模扩张情景下土地所有者的回报最高,但社会净效益最低,生态系统碳固定、水质净化、栖息地质量都将最大程度下降[50]。Swetnam等对坦桑尼亚东部的碳储量设置“可持续发展”和“一切照常”两种社会经济情景,基于利益相关者和专家知识的土地利用制图表明碳储量在第一个情景减少4%而在第二个情景减少41%[51]。Chaplin-Kramer等则进一步构建精细化的全球情景,发现在非洲和南亚多达50亿人面临更高的水污染和导致营养不足的授粉服务,非洲、欧亚大陆和美洲数以亿计的人面临着沿海风险的加剧[4]。然而,有学者认为当前的生态系统服务评估方法和预测模型并不具备处理大尺度跨学科分析的能力,既然生态系统服务预测与空间管理决策密切相关,则模型开发中应当更注重参与性,允许大量利益相关方参与模型的训练[52]。由此引申,考虑到利益相关者一般具有局地视角,生态系统服务建模中基于生物多样性形成机制的参数本地化对保障预测合理性而言是非常重要的。
3 基于情景模拟的生态系统保护决策支持展望
从全球、国家、区域到社区,不同尺度上的生态系统保护关注重点具有很大的差异,其决策支持方式也不一而足。本研究分别从大尺度区域、景观和局地社区3个尺度出发,选取生态区划、生态安全格局、社会生态网络3个重点研究切入点,展望上述研究方向在不同尺度上对我国生态系统保护的决策支持潜力(图4)。
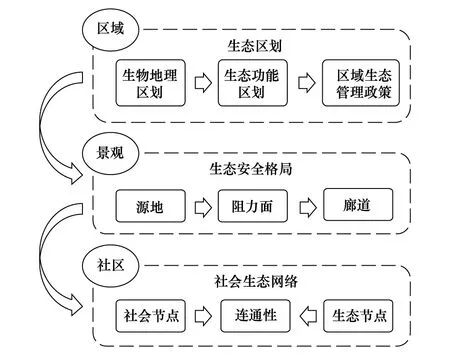
图4 生物多样性保护决策支持多尺度研究方向 Fig.4 Multi-scale research direction on the decision support for biodiversity conservation
3.1 生态区划与区域尺度情景模拟
生态区划是对生态区域和生态单元的划分或合并研究,通过揭示自然生态区域的相似或差异规律,为生态系统管理和资源持续利用提供科学支撑[53]。目前在全球生物多样性管理中,使用最多的是Olson等自下而上融合形成的全球生态区和全球优先保护区,其为大尺度生物多样性保护提供明确的边界位置[54]。目前最新更新的Ecoregions2017方案由846个生态区构成,并将生物多样性保护面积达全球陆域面积的50%作为2050年目标[55]。然而,有研究基于情景假设发现2亿多种植物、动物和真菌的空间更替是连续的,在很大程度上独立于生态区域边界[56]。另有研究也发现了当前的生态区方案并不能显示生态系统服务的空间差异,从而削弱了生物多样性保护决策与人类福祉的递进联系[57]。考虑到区域生物多样性形成机制与生态系统服务模拟能力,面向区域生态系统综合管理的生态区划方案仍有改进空间。
区域尺度生物多样性、生态系统服务情景与生态区划存在互馈关系。一方面,基于生态系统服务模拟有助于生态功能区划方案建立。目前的大多数生态区划重在描述生物地理特征,本质上属于生物地理区划;而加入生态系统服务的空间信息,有效表征生态系统对人类社会的功能,无疑将更加符合IPBES的理念,将给生态管理者提供更丰富的信息[58]。我国学者在生态功能区划领域积累的大量经验也将为“一带一路”等区域跨国生态系统管理提供有力的理论与方法指引[59]。另一方面,生态区划边界的明确有助于提升生物多样性和生态系统服务在大尺度情景模拟中的参数衔接能力。在生态系统服务模拟中,由于对生态系统参数的相互作用关系考虑不足,大尺度模拟结果的信度存疑,并未能实际应用于大尺度生态系统保护决策中。基于我国生态地理区划与生态功能区划方案良好基础,进一步率定区域生态系统尺度上的生物多样性模型参数,可以为建模提供更加统一的标准。此外,相关情节模拟结果也有必要与我国国土空间规划实践紧密结合。在决策角度上大尺度自然资源统一管理有必要更多考虑未来生态系统演化情景的不确定性[60],而在建模角度上生态空间的范围、生态红线的边界等也都是情景模拟中值得被纳入的政策变量。
3.2 景观尺度下的生态安全格局构建
生态安全是对生态系统完整性和健康水平的整体描述,我国学者提出的区域生态安全格局是针对区域生态环境问题,能够保护和恢复生物多样性、维持生态系统结构和过程的完整性、实现对区域生态环境问题有效控制和持续改善的区域性空间格局[61]。生态安全格局构建涉及的空间单元需要与生态过程发生的生态空间相一致,由于具有明确的空间位置信息,有助于提供区域生态恢复中具体的景观格局优化配置途径[62]。生态安全格局构建的主要途径分解为源地识别、阻力面设置、廊道提取等步骤,源地生态特征评估的指标、阻力面生成的数据、廊道识别的算法成为影响生态安全格局现实效果的主要因素[63]。而目前上述研究要素在生物多样性与生态系统服务形成机制的刻画上并不成熟,从而影响了生态安全格局理念在生态系统保护决策中的应用。
针对我国生态安全保障的区域诉求,以及生物多样性保护和区域社会发展的权衡关系,服务于我国景观尺度生态系统保护的生态安全格局在方法论上有待深化。第一是把握要素整体性。生态安全与粮食安全、经济安全、社会安全的权衡协同关系复杂,尤其在我国干旱区、喀斯特地区、海岸带、高海拔山地等生态脆弱区,在对自然保护区严格限制人类活动强度的同时,模拟并遴选景观发挥生产、生活、生态多种生态系统服务的协同最优情景,提升非自然保护地范围内的景观多功能性,符合区域绿色发展的基本逻辑。第二是聚焦区域异质性。区域生物多样性特征决定了不同的源地范围和廊道长度、宽度,也决定了阻力面的评价标准,生物多样性数量、质量及结构参数有必要在指标中予以强化。同时,生态系统服务空间流动的视角可以体现不同目标情景下的受益区、受益对象和受益程度,有必要从生态系统服务需求角度出发为生态安全格局构建提供定量依据。第三是强调情景动态性。由于气候变化情景下未来生物多样性特征具有不确定性,在生态安全格局构建中需要考虑更加长远的评价规则,通过模拟代表性气候和人类活动情景,兼顾生态安全格局的稳定性、持续性等特征,从而增强生态系统对未来环境变化的适应能力。
3.3 基于社会生态网络的社区适应
传统的社会生态系统适应性理论既强调政府决策的重要能力,也强调社区传统知识的应用,但如何搭建政府和社区多方协作决策的桥梁一直是适应性研究的难点。Bodin等将社会生态系统理解为生态节点和社会节点构成的空间网络结构,社会节点代表进行决策的个人、组织或其他社会实体,生态节点包括物种、生态群落或其他一些生态概念;并基于指数随机图模型,量化了决策者之间的社会协作、生态要素间的相互依赖,以及社会生态相互作用[64]。在网络的刻画中,必须明确社会生态要素间的因果关系类型,包括:网络连接影响着个体/节点如何受到其他个体/节点的影响(例如种群的迁移、知识的转播),个体/节点根据其他个体属性选择网络伙伴(例如食物网中的物种丰富度、教育背景),网络结构和节点属性分布产生了一定的全局后果(例如不同簇组成的食物网抗干扰能力强,人在密集社会网络中更倾向集体行动),网络结构和节点属性随时间推移而共同进化(例如渔民的物种选择由渔具决定,目标鱼量下降影响着海洋食物网动态)[65]。由此,个体的网络伙伴选择将通过簇的形式影响网络结构,从而产生深远的个体或全局后果。
模拟不同情景下我国社区自下而上的社会生态网络无疑将为国家生态系统保护决策提供真正意义上的机理性、结构化支撑。在我国快速城市化现状与乡村振兴政策背景下,根据网络的复杂性可以分为三层具有递进关系的适应途径。第一层是明晰城乡社区个体行为与特定动植物的因果关系,分别识别特定物种与生态系统的关联路径,以及城市、乡村、景区等社会群体与个体的关联路径。构建情景模拟不同的个体决策分别对特定物种、生态系统、群体行为产生的作用,预判相应后果用于对市民、农户、游客等个体行为的约束和引导。第二层是明晰公共政策与区域生物多样性水平、栖息地质量的因果关系,分别识别公共政策对社区群体行为的影响,社区群体行为对栖息地质量的多重影响,以及栖息地质量变化对区域生物多样性水平的影响。构建情景模拟不同的公共政策下城市居住区、粮食基地、国家公园等社区群体的决策方式,及其所对应的栖息地变化概率与程度,从而对未来生物多样性整体水平做出预判,作为城市绿地、基本农田、自然保护区等不同对象的生态管理政策调整的依据。第三层是明晰跨区域政策对生态系统服务流的影响路径,以及生态系统服务流对人类福祉的区域和区际反馈路径,分别识别区域生物多样性变化下的生态系统服务流向、流量、流速,区域政策与生态系统服务流特征的影响路径,以及生态系统服务流变化的局地效应和外溢效应。构建情景模拟城市群、农产品主产区、禁止开发区等不同区域政策下生物多样性改变造成的生态系统服务亏损或盈余,以及多种公共资源分配方案或生态补偿标准,预判城乡群体或个体与之相对应的决策方式,遴选面向区域共赢、多主体共赢的社会生态系统适应途径。
4 结语
生物多样性和生态系统服务情景模拟是制定生物多样性保护措施的重要科学依据,然而当前模拟结果的较高不确定性影响了区域生态系统保护方案对未来环境变化的应对能力。本研究通过梳理生物多样性与生态系统服务情景分析常用模型,综述国际上具有充分代表性的生物多样性与生态系统服务预测结果,为我国生物多样性与生态系统服务情景研究提供引导。在建模分析角度,生物多样性与生态系统服务参数关联机制的明晰是提升模型输出情景准确性的重要条件和前沿议题。在管理应用角度,生态区划与区域尺度情景模拟、景观尺度下的生态安全格局构建、基于社会生态网络的社区适应三点重要研究方向将对基于情景模拟的我国生态系统保护决策提供重要的理论和实践支持。
猜你喜欢
疯狂英语·初中天地(2022年2期)2022-07-07
内蒙古民族大学学报(社会科学版)(2020年2期)2020-11-06
科学大众(中学)(2019年3期)2019-05-17
劳动保护(2019年3期)2019-05-16
汽车观察(2018年10期)2018-11-06
小天使·一年级语数英综合(2017年3期)2017-04-25
科技知识动漫(2017年1期)2017-02-06
太空探索(2016年5期)2016-07-12
少儿科学周刊·少年版(2015年1期)2015-07-07
小天使·一年级语数英综合(2015年8期)2015-07-06
