
黄河流域干旱区湖泊冰封期浮游植物群落结构特征及影响因子研究
2020-11-12 09:20王司阳张笑欣田世民曹永涛陈融旭
水利学报 2020年9期
王司阳,张笑欣,田世民,曹永涛,陈融旭
(1.黄河水利科学研究院,河南 郑州 450003;2.北京师范大学 环境学院,北京 100875)
1 研究背景
湖泊是全球水资源的重要组成部分,是维护水域生态系统健康发展的关键因子。其中,沉积物-水界面的生物地球化学循环是驱动湖泊水环境演变的重要动力[1]。因此,研究营养元素的变化及其生物作用,对于维护湖泊生态系统平衡具有重要意义。由于我国北方干旱地区生态系统的脆弱性,近些年随着人类活动加剧,干旱区湖泊生态系统受到了不同程度影响,富营养化程度逐渐加剧[2]。湖泊作为一类独特的生态系统,一般存在清水草型或浊水藻型两种稳定的状态。当某些关键营养元素过量输入(如氮、磷)会导致清水草型向浊水藻型转换,即水体富营养化。浮游植物作为湖泊生态系统中的主要初级生产者,能够进行光合作用,当浮游植物生长时会按比例吸收水体中营养物质,当浮游植物死亡降解时,它们亦会按相同的摩尔比向水体释放营养物质,并调节水体中的营养物质比例[3]。由于不同浮游植物类群对于化学元素的需求呈现分异,长期或者短期外界营养状态的变化会引起浮游植物群落的改变。例如1998—2012年波罗的海研究结果表明,氮和磷的浓度在调节蓝藻群落组成方面具有指示意义[4];氮浓度可以显著增如微囊藻和其他非固氮蓝藻相对丰度,固氮蓝藻在高磷等环境下具有更高丰度[5];采石场湖泊原位和室内研究结果显示,只增加磷的浓度可以提高浮游植物生物量,物种群落结构也发生改变;单独提高温度和增加硝酸盐浓度则对群落结构无显著影响[6]。因此,浮游植物物不仅可以响应外界环境的营养状态变化,而且亦能对其发生反馈效应。
相较于研究较为广泛的东南部平原以及云贵高原湖泊,黄河流域上游干旱地区湖泊具有独特区域特征和气候环境,干旱区湖泊存在着冰封期与非冰封期的交替变化。由于冰盖的阻隔,冰封期水体水质特征、流速结构等会发生改变,冰体中大量的营养物质迁移到水体中,加剧了水体的富营养化,且可能对浮游植物群落结构特征产生影响[7]。西南格陵兰岛研究表明,在冰盖和无冰环境下,水体分别表现为硝酸盐限制与磷限制[8]。换言之,冰盖的存在可能影响湖泊氮磷等营养循环状态,进而对浮游植物生长产生重要影响。然而,干旱区湖泊冰封期条件下营养循环状态模式仍未得到充分研究,其与浮游植物群落结构的响应关系仍不甚明晰。因此,本文以黄河流域典型干旱区湖泊乌梁素海、奈伦湖为研究对象,通过冰封期野外调查与监测分析,研究干旱区湖泊冰封期浮游植物群落结构变化特征及其与环境因子的响应关系,为流域干旱区湖泊治理与保护提供科学依据。
2 材料与方法
2.1 采样点分布由于特殊水文情势、地理环境和气候特征,黄河上游内蒙古河段冬春季存在封河阶段(冰封期),并且易出现“凌汛”现象。为了防止凌汛灾害,常启用分凌区进行分凌调度确保黄河凌汛期安全[9],其中河套灌区及乌梁素海分凌区(乌梁素海)、乌兰布和分凌区(奈伦湖)为黄河上游最主要的两个分凌区。本研究选取位于内蒙古自治区巴彦淖尔市的乌梁素海、奈伦湖作为黄河流域上游干旱区的典型湖泊,分析冰封期湖泊浮游植物群落结构及影响因子。本研究共设9个采样点,采样点设置考虑湖泊地理环境和空间分布情况,包括退水渠、入湖口、湖区及重要影响区域,采样点设置便于样品采集与监测。其中,河套灌区及乌梁素海分凌区的乌梁素海为W1—W6(6个采样点),W1为退水渠,W2为新安生态沟扬水站区域,W3为红圪卜扬水站区域,W4为隔堤桥,W5为二分滩,W6为旅游码头区域;乌兰布和分凌区的奈伦湖为N1—N3(3个采样点),N1为引水渠入湖口,N2为东部湖区,N3分别为西部湖区。采样点位置分布见图1所示。
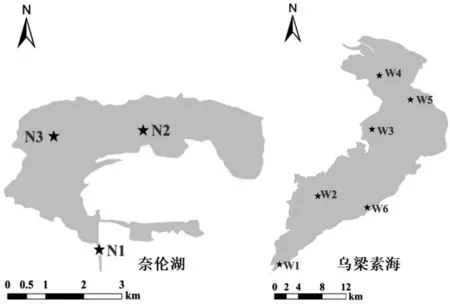
图1 研究区域采样点分布
2.2 样品测试分析方法本研究于2019年3月湖泊冰封期进行样品采集,用冰钻采集器破冰后,现场通过YSI EXO2水质分析仪测量水体水温、溶解氧、TDS、盐度、氧化还原电位、电导率、pH等理化参数;运用赛氏盘(黑白盘)法现场测定湖泊水体透明度;通过有机玻璃采水器采集水面下0.5 m处水样,并及时运回实验室参照《水和废水监测分析方法》[10]分析总磷、总氮、磷酸盐、氨氮、硝酸盐、高锰酸盐指数、叶绿素a等水质参数;浮游植物定量样品是通过采水器采集1 L水样后,现场加鲁哥试剂固定带回实验室处理,静置沉降48 h,用虹吸管缓慢吸去上清液,将样品浓缩定容至30 mL,摇匀吸取0.1 mL浓缩样品置于计数框内,在光学显微镜下进行鉴定计数;浮游植物定性样品是在每个采样点选取3处位置通过采水器各采集1 L水样后,用25号浮游生物网(网孔为0.064 mm)浓缩过滤至100 mL 标本瓶,现场用5%的甲醛溶液固定保存。浮游植物定性和定量样品按照《中国淡水藻类——系统、分类及生态》[11]、《中国淡水生物图谱》[12]等方法和描述用显微镜进行分类鉴定和定量计数;浮游植物生物量(湿重)参照章宗涉等[13]方法,按照测量体积进行换算。每个样品测试设置三组平行,平行误差均小于10%。
2.3 数据处理分析方法
2.3.1 综合营养状态指数 湖泊富营养化程度评价采用综合营养状态指数法TLI进行评价[14],选取了反映乌梁素海营养程度的叶绿素a、总磷、总氮、透明度和高锰酸盐指数5 项主要指标。采用0~100的一系列连续数字对湖泊营养状态进行分级:TLI(∑)<30为贫营养;30≤TLI(∑)≤50为中营养;TLI(∑)>50为富营养,其中50<TLI(∑)≤60为轻度富营养,60<TLI(∑)≤70为中度富营养,TLI(∑)>70为重度富营养。具体计算公式为:

式中:TLI(∑)为综合营养状态指数;Wj为第j种参数的营养状态指数的相关权重;TLI(j)为第j种参数的营养状态指数。
2.3.2 优势度 浮游植物优势种是根据物种的出现频率及个体数量采用优势度y来判定[15],当优势度y≥0.02即定为优势种。优势度y计算公式如下:

式中:ni为第i种的个体数;fi为i物种在不同采样点出现的频率;N为所有物种的个体总数。
2.3.3 多样性指数 浮游植物多样性特征采用Shannon-Wiener 多样性指数H′来测定[16-18]。其中,H′值在0~1.0为重度污染,1.0~2.0为中度污染,2.0~3.0为轻度污染,大于3.0为清洁环境。具体计算公式如下:

式中:ni为第i种的个体数;N为所有物种的个体总数;S为物种数。
2.3.4 群落相似性指数 浮游植物群落结构相似性采用Jaccard种类相似性指数J[19]计算。具体计算公式如下:

式中:a为采样点1中出现的浮游植物种类数;b为采样点2中出现的浮游植物种类数;c为采样点1和采样点2中共同出现的浮游植物种类数。由式(4)可知,浮游植物群落相似性仅与物种种类相关,与物种数量没有关系。群落相似性等级一般划分为6级[20],具体分级标准如表1所示。

表1 相似性分级评级标准
2.3.5 群落结构与环境因子影响分析 应用R语言研究藻类群落与环境因子的关系,物种数据矩阵和环境数据矩阵都经lg(x+1)转换。本研究用于排序的物种在各样点的出现频率大于12.5%,在至少一个采样点的相对丰度大于1%,同时满足该物种优势度不小于0.02。通过对物种数据进行降趋势对应分析(DCA)判断是选择基于单峰模型的典范对应分析(CCA)或基于线性模型的冗余分析(RDA),如DCA 最长轴的长度小于3,选择RDA模型;介于3到4 之间,选择CCA和RDA模型均可;大于4,选择CCA模型。为了筛选有效的环境因子,本研究首先试探性将所有环境因子作为解释变量纳入排序分析,用函数permutest()进行蒙特卡罗置换检验,分析环境因子对物种分布的解释量是否具有显著性。为了简化模型,运用函数envfit()检验每个环境因子的显著性从而筛选确定最终环境因子,最后通过方差分解分析(Variation Partitioning Analysis)关键环境因子对优势物种群落分布的贡献[21]。
3 结果分析
3.1 冰封期浮游植物组成与分布乌梁素海和奈伦湖冰封期不同采样点TLI结果显示,乌梁素海各采样点(W1—W6)TLI指数变化范围为61.0~69.9(平均值为65.7),按照湖泊营养状态分级均属于中度富营养;奈伦湖各采样点(N1—N3)TLI指数变化范围为52.3~65.0(平均值为59.3),按照湖泊营养状态分级均属于轻度富营养—中度富营养(图2)。总体而言,乌梁素海和奈伦湖冰封期主要水质参数含量较高,水体均处于富营养化状态。
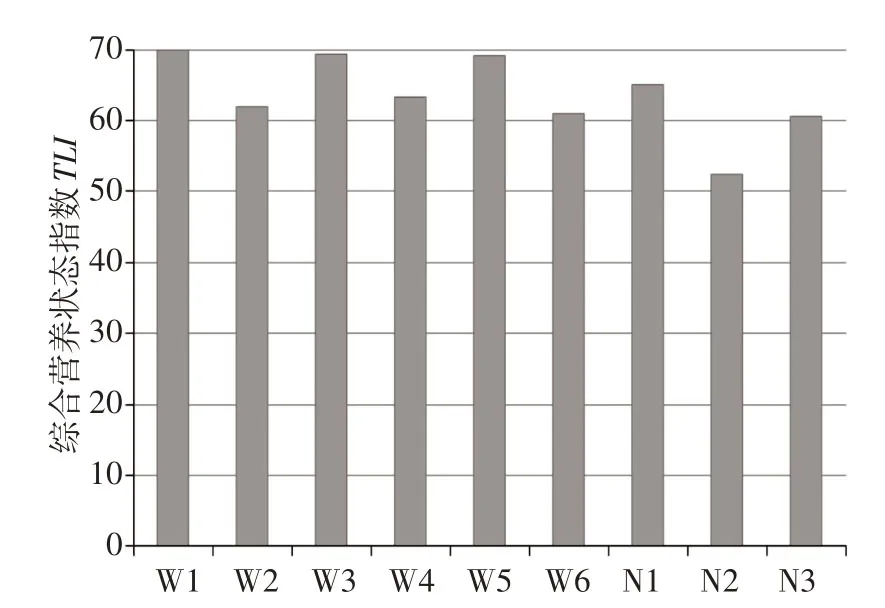
图2 不同采样点综合营养指数分布

图3 不同采样点浮游植物组成
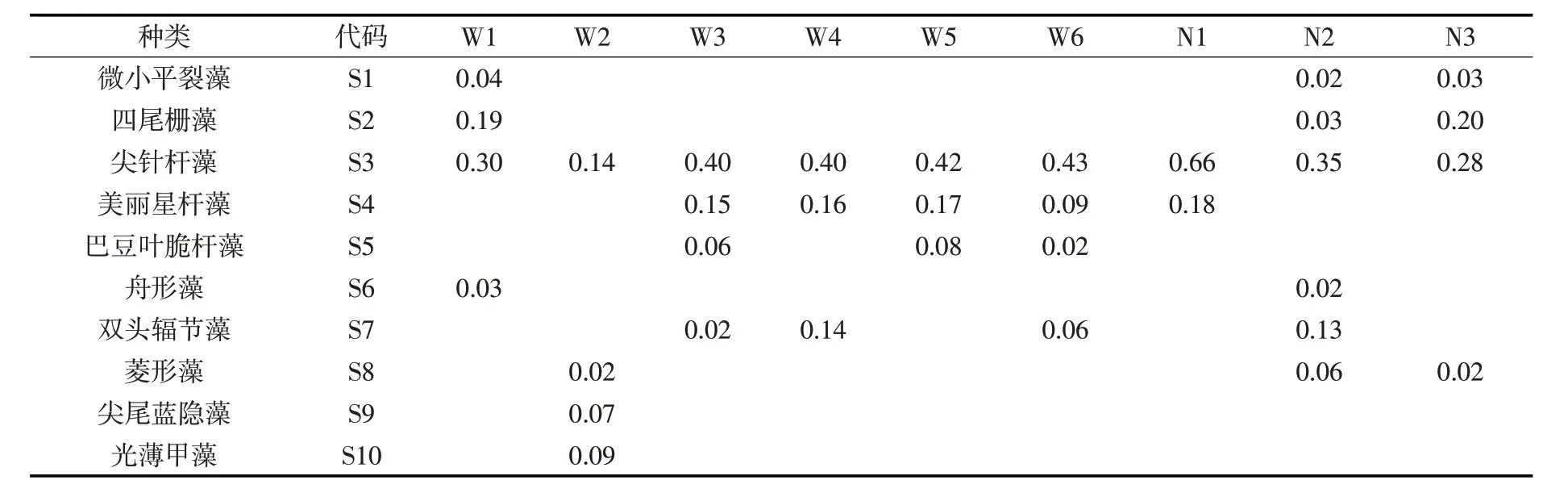
表2 不同采样点浮游植物优势种类及优势度
本研究湖泊鉴定浮游植物共计6门21属29种,其中蓝藻2属2种,绿藻4属7种,硅藻12属16种,裸藻1属1种,隐藻1属2种,甲藻1属1种。硅藻物种数占据全部物种的55.2%,在所有采样点中藻类组成中占据明显优势(图3)。根据不同采样点浮游植物优势度统计结果,本研究湖泊优势种包括微小平裂藻、四尾栅藻、尖针杆藻、美丽星杆藻等10种,其中尖针杆藻、美丽星杆藻、双头辐节藻等优势度较为明显(表2)。如图4所示,研究湖泊冰封期不同采样点浮游植物数量和生物量均处于较低水平。分湖区来看,乌梁素海浮游植物数量范围为1.1×104~1.4×105cells/L(平均数量为8.5×104cells/L),生物量范围是0.015~0.328 mg/L(平均生物量为0.143 mg/L);奈伦湖浮游植物数量范围为3.2×104~8.3×105cells/L(平均数量为4.2×104cells/L),生物量范围是0.047~0.138 mg/L(平均生物量为0.101 mg/L)。乌梁素海浮游植物数量/生物量均高于奈伦湖,但是无统计上显著差异。
3.2 冰封期浮游植物多样性与群落相似性多样性指数为浮游植物群落特征的重要参数,是衡量群落稳定特性的重要指标。如图5所示,浮游植物Shannon-Wiener 多样性指数H′为0.70~1.74,平均值为1.49,介于中度污染,即乌梁素海和奈伦湖浮游植物多样性较差。群落相似性指数是基于物种种类数量变化,分析两个群落之间的相似程度的重要指标。如表3所示,不同采样点位之间浮游植物群落相似性主要处于Ⅱ、Ⅲ等级,即为极不相似和轻度相似,部分采样点(如N2与N3)群落结构为中度相似(Ⅳ等级)。由于群落相似性分析基于物种种类数量的变化,本研究调查监测发现不同采样点物种种类数均较低,且不同点位的环境因子差异亦可能影响物种分布,故可能导致群落间相似性较低。
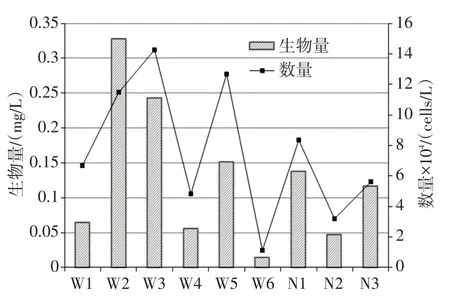
图4 不同采样点浮游植物数量/生物量分布
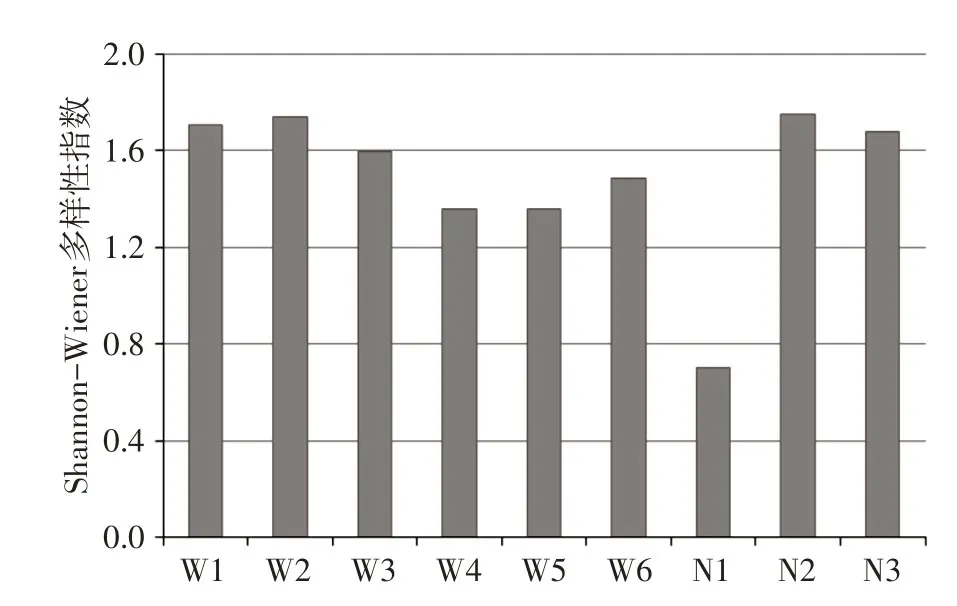
图5 不同采样点浮游植物多样性变化

表3 不同采样点浮游植物群落相似性指数
3.3 浮游植物群落结构与环境因子关系根据浮游植物相对丰度以及优势度,选择10个优势种参与RDA排序(S1—S10,表2),利用前向选择(forward selection)对透明度、总氮、总磷、叶绿素a、高锰酸盐指数、硝酸盐、氨氮、磷酸盐、水温、溶解氧、TDS、盐度、氧化还原电位、电导率、pH等15个环境因子进行逐步筛选,通过蒙特卡罗置换检(Monte-Carlo permutation test)进行显著性分析,结果显示浮游植物群落结构显著环境因子为水温、总氮和硝酸盐(P<0.05)。如图6所示,水温、总氮和硝酸盐共同解释浮游植物优势种分布的67.49%,其中第一排序轴和第二排序轴累计解释物种数据的63.11%。浮游植物与环境因子分布关系显示,尖尾蓝隐藻、光薄甲藻数量随着总氮含量增加而显著降低;微小平裂藻、四尾栅藻、舟形藻、菱形藻等数量随着水温的增加而升高,同时受硝酸盐浓度抑制;尖针杆藻、美丽星杆藻、巴豆叶脆杆藻、双头辐节藻等数量随着硝酸盐的增加而升高,同时受水温升高抑制。方差分解分析进一步显示(图7),营养因子(总氮和硝酸盐)单独对优势种物种分布解释量达到33.66%,理化因子(水温)单独对优势种物种分布解释量达到13.24%,营养因子与理化因子共同解释量为20.59%。
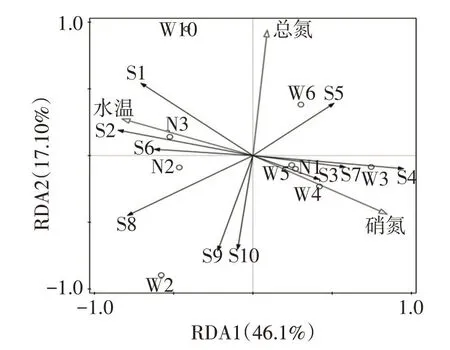
图6 浮游植物群落结构与环境因子RDA分析
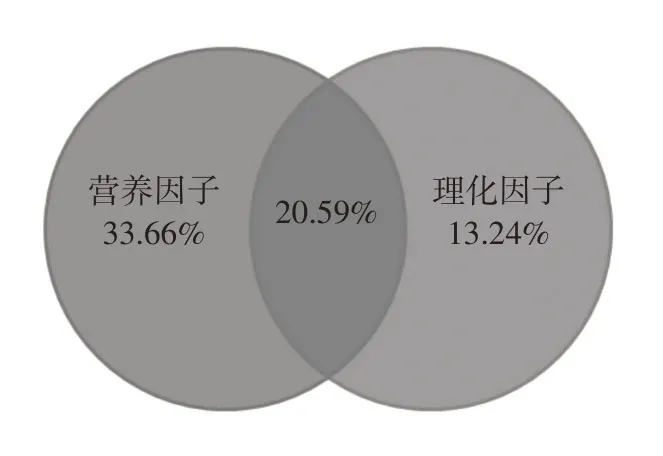
图7 基于偏RDA的方差分解分析韦恩图
4 讨论
4.1 浮游植物群落结构特征分析本研究中共鉴定浮游植物6门21属29种,其中硅藻12属16种,占据全部物种的55.2%,在不同湖区(乌梁素海、奈伦湖)冰封期的采样点中硅藻均优势明显(图2)。硅藻是一类细胞壁高度硅质化的单细胞植物,耐受力极强、生境十分广泛,可以生长在极地、寒带、高山、高原、湖泊、水库等不同生境[22]。由于硅藻门多为狭冷性物种[23],能够在低温条件下通过调节生理代谢适应外部环境,从而取得竞争优势[24]。黄河青海河段段30 多年研究结果对比显示,其浮游植物的群落结构比较稳定,仍以河流性硅藻为主[25],本研究湖泊均位于同流域寒冷干旱地区,故表现出相应的群落结构特征。牡丹江流域浮游植物调查结果表明,冰封期水体浮游植物以硅藻(占总物种数的39.29%)和绿藻(占总物种数30.36%)为主[26],乌梁素海早期研究结果亦显示,冰封期水体以绿藻、硅藻种类数量居多,分别占总物种数的33.3%和26.7%[27]。本研究湖泊冰封期浮游植物群落组成具有一致规律,即以硅藻和绿藻为主,占据浮游植物全部物种数的79.3%,浮游植物物种组成呈明显硅藻-绿藻型;与其它水体相异的是,研究湖泊硅藻占比明显高于其它水体,这可能是由于气候变化等因素造成研究区域温度呈下降趋势[28],而低温环境更加有利于硅藻竞争优势。浮游植物优势种种类数及其数量对群落结构的稳定性亦有重要影响,优势种种类数越多且优势度越小,则群落结构越复杂、稳定[29],本研究中浮游植物优势物种数为10种,主要优势物种为尖针杆藻、美丽星杆藻、双头辐节藻(表2),表明研究湖泊的群落结构相对较为简单,针杆藻等由于生态位较宽,更适合冬季气候特征,从而具有群落竞争优势[30]。由于水文地质和营养结构等因素影响,不同采样点浮游植物数量以及生物量仍有一定差异,但冰封期浮游植物数量和生物量均处于较低水平。与本研究较为一致的是,冰封条件下扎龙湿地藻类表现出较低水平[31],且冰盖的存在亦会影响浮游植物群落结构[32]。
不同水体的浮游植物群落具有不同的结构特征[33-34],本研究采用生物多样性指数和群落相似性指数分析干旱区湖泊浮游植物群落结构特征变化。生物多样性是反映生物群落组成特征的重要参数,物种的生物多样性指数越高,通常表明群落中的生物种类越多,群落结构越稳定[35],常用于反映群落结构的复杂程度[36];Jaccard群落相似性指数一般用来表示不同采样点之间物种分布的相似程度[37]。本研究中,浮游植物物种多样性指数与丰富度指数值均较低,表明研究湖泊浮游植物群落结构不稳定,物种组成较为单一。一般来说,多样性指数越高,水质越好[38];在本研究中,依据Shannon-Wiener多样性指数值判断,研究湖泊介于中度污染。牡丹江流域冰封期浮游植物多样性评价结果表明,42%断面处于中度污染[26]。冰封期水体气候条件形成了特殊的生存环境,对浮游植物耐受能力要求较高,硅藻和绿藻作为重要浮游植物类群,适应低温和高营养盐浓度环境,较易形成优势类群[39]。浮游植物生物多样性特征与群落结构易受季节等因素影响,生物多样性指数常表现为冬春季低于夏秋季[25],本研究中不同采样点浮游植物群落相似性处于Ⅱ、Ⅲ等级,即为极不相似和轻度相似,奈伦湖部分采样点群落结构为中度相似(Ⅳ等级),可能原因是不同点位浮游植物群物种种类数均较低,从而基于物种种类数量的群落结构相似性较低,由于本研究调查时间范围为冰封期,受采样时间、频次和范围等影响,浮游植物监测结果亦可能存在一定差异;另一方面不同点位的环境因子差异亦可能影响物种分布,导致浮游植物群落间相似性较低。
4.2 浮游植物群落结构特征分析不同营养状态湖泊浮游植物响应方式不同,其影响因子亦存在差异。水体中影响浮游植物生长的主要环境因子包括光照、温度、总磷、总氮、COD等,并且在不同类型的水体中,影响浮游植物群落结构的环境因子各有差别[40-42]。黄河流域上游干旱地区湖泊由于特殊的地理环境和气候特征,存在着冰封期与非冰封期变化,冰封期的冰体会影响水体与大气复氧、污染物迁移分布及太阳净辐射等[43-44],从而使冰封期水体营养元素分布以及浮游植物群落结构特征均表现出一定典型性和特殊性。在冰封条件下,达里诺尔湖水体中总氮和总磷的含量是其上覆冰体中的含量的9.1倍和3.6倍,略高于非冰封期水体中总氮和总磷的相应值[45]。冰封期与非冰封期相异的水文气候特征也会导致不同营养利用模式[8],冰盖的阻隔会影响水体流动和加速污染物迁移,冰体中大量的营养物质迁移到水体中,加剧了水体的富营养化,且可能对藻类群落结构分布产生影响,本研究湖泊在冰封条件下表现出明显富营养化湖泊特征,浮游植物群落组成呈以耐低温环境和高营养盐浓度的硅藻和绿藻为主。一般而言,影响浮游植物群落结构的因素较多,目前广泛认同的是浮游植物群落结构受生物、物理、化学等影响因素共同作用。Padisák等[46]对欧洲8个具长期观测数据的湖泊中浮游植物群落演替进行分析,表明物理因素(光的可利用性、温度、电导率、沉积物扰动))与生物因素(基本营养元素可利用性、牧食作用和细菌寄生)对浮游植物群落演替具有重要作用。黄河流域干旱区湖泊由于气候变化等因素造成生物群落结构及其功能影响仍知之甚少,尤其是冰封条件下浮游植物群落变化特征及其影响因子的研究研究仍值得重点关注。
冬季冰层覆盖会对湖泊生态系统的食物网结构产生显著影响[32],海冰消退时间、光照有效性、海面温度、营养和盐度显著影响冰封期浮游植物群落组成[47]。冰盖的变化伴随着浮游植物群落结构变化,在冰层覆盖条件下,丝状硅藻大量繁殖;而无冰条件则促进小尺寸浮游植物细胞的生长[31]。本研究湖泊采样时间为冰封期阶段,温度较低,浮游植物主要优势物种为尖针杆藻、美丽星杆藻、双头辐节藻等硅藻,适应于低温环境生长。不同的水体环境存在不同的浮游植物群落结构组成,如长江口北支邻近海域水温为18.7~21.7 ℃,盐度介于16.8~28.7之间,适宜骨条藻的生长,使其成为绝对优势种[48];同时浮游植物群落结构的差异会影响水体环境因子的变化,北极浮冰下的金藻水华导致水体表面硝酸盐存量几乎耗尽[49]。由于气候变化等因素影响,研究区域近几十年冬春季温度呈下降趋势[28],即表现出冰封期水体温度更低;另一方面,湖泊冰封期阶段由于冰体排斥作用,导致冰体污染物迁移至水体,加剧水体富营养化程度[50],从而表现出冰封期水体氮磷等营养盐浓度高于非冰封期。较低的温度环境和较高的营养盐浓度有利于部分硅藻和绿藻竞争优势,从而表现出明显的硅藻-绿藻型。RDA研究结果显示,冰封期浮游植物关键影响因子为总氮、硝酸盐和水温,其中,总氮和硝酸盐作为关键影响因子与扎龙湿地研究结果较为一致[51]。方差分解分析进一步显示,营养因子(总氮和硝酸盐)对浮游植物分布的影响更为关键,可能原因是占据优势的硅藻和绿藻对硝酸盐等氮源需求更高,同时研究区域位于河套灌区,存在大量农业非点源氮排放导致[52]。需要说明的是,本研究湖泊采样时间处于分凌调度前的冰封期,且此阶段农田作业影响相对较小,即湖泊处于相对自然状态,考虑到影响湖泊浮游植物的因素较多,本研究主要通过分析冰封期浮游植物群落特征及其影响因子,以期为开展黄河流域干旱区湖泊健康评价及生态修复提供重要支撑。
5 结论
(1)在冰体形成和生长过程中,营养盐等主要污染物从冰层迁移至水体中,从而导致冰封期水体营养盐含量较高,表现出明显富营养化特征。本研究湖泊乌梁素海、奈伦湖综合营养状态指数分别为65.7和59.3,属于中度富营养和轻度富营养。研究湖泊共鉴定浮游植物6门21属29种,乌梁素海和奈伦湖浮游植物组成较为一致,均以硅藻和绿藻为主,占据浮游植物全部物种数的79.3%,浮游植物种类组成呈硅藻-绿藻型。由于气候变化等因素的影响,研究区域气温呈下降趋势,从而加剧硅藻竞争优势。(2)研究湖泊冰封期不同采样点浮游植物数量和生物量均处于较低水平,乌梁素海浮游植物数量/生物量均高于奈伦湖,但无显著差异。从主要优势种来看,乌梁素海主要为尖针杆藻、美丽星杆藻和巴豆叶脆杆藻,奈伦湖为尖针杆藻、微小平裂藻和四尾栅藻。乌梁素海和奈伦湖浮游植物多样性均较差,按照多样性指数评价标准介于中度污染,且不同采样点之间浮游植物群落相似性较低,而不同采样点环境因子的差异可能是导致群落结构差异重要因素。(3)维持河湖生态系统健康是幸福河的基本要求,也是推进黄河流域生态保护和高质量发展的重要保障。本研究结果显示,影响湖泊冰封期浮游植物物种分布的主要环境因子水温、总氮和硝酸盐,且不同采样点之间存在差异。方差分解分析结果进一步表明,营养因子(总氮和硝酸盐)对浮游植物物种分布的影响程度高于理化因子(水温),这可能是与湖泊冰封期浮游植物以硅藻-绿藻为主,且部分硅藻、绿藻对硝酸盐需求较大,本研究为开展黄河流域河湖健康评价及生态修复提供基础支撑。
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
疯狂英语·新读写(2021年8期)2021-11-05
奇闻怪事(2021年2期)2021-04-20
潍坊学院学报(2020年2期)2021-01-18
海洋通报(2020年5期)2021-01-14
黑龙江水利科技(2020年8期)2020-01-12
流行色(2019年10期)2019-12-06
少儿美术(快乐历史地理)(2019年11期)2019-04-20
小朋友·聪明学堂(2015年12期)2016-01-07
