
不同春季追氮模式对小麦茎秆抗倒性能及木质素积累的影响
2020-11-09 05:17董荷荷骆永丽李文倩王元元张秋霞陈金金敏李勇王振林
中国农业科学 2020年21期
董荷荷,骆永丽,李文倩,王元元,张秋霞,陈金,金敏,李勇,王振林
不同春季追氮模式对小麦茎秆抗倒性能及木质素积累的影响
董荷荷,骆永丽,李文倩,王元元,张秋霞,陈金,金敏,李勇,王振林
(山东农业大学农学院/作物生物学国家重点实验室,山东泰安 271018)
【】探讨春季不同追氮模式对小麦各节间茎秆抗倒伏能力、木质素积累及籽粒产量的影响,明确高施氮量条件下适宜的春季追氮模式,为小麦高产稳产抗逆应变栽培提供技术支撑。于2017—2018和2018—2019年2个小麦生长季,以倒伏敏感型品种山农16和抗倒伏品种济麦22为供试材料,在高施氮量300 kg·hm-2基施1/3条件下设置4种春季追肥模式,分别为等量二次性追氮和剩余一次性追氮,即起身期﹕孕穗期1/3﹕1/3(T1),拔节期﹕开花期1/3﹕1/3(T2),孕穗期一次性追施剩余2/3氮(T3)和拔节期一次性追施剩余2/3氮(CK)。深入研究春季不同追氮模式对冬小麦植株茎秆抗折力、木质素积累、木质素合成关键酶基因的表达丰度及籽粒产量的调控效应。抗倒伏品种济麦22的各节间茎秆抗折力、木质素积累量以及单体含量均高于倒伏敏感型品种山农16,2种类型品种开花期T1、CK处理的抗折力高于T2和T3处理,木质素积累量、单体的含量表现为T1>T3>CK>T2,灌浆期和成熟期各处理间抗折力、木质素积累量以及单体的含量表现为T1>T3>T2>CK。灌浆期山农16和济麦22在T1处理下抗折力较CK、T2、T3处理分别增加24.69%、19.97%、13.15%和26.92%、15.36%、5.87%;山农16和济麦22在T1处理下的各生育阶段木质素积累量平均值分别较CK、T2、T3处理提高了21.71%、15.45%、8.85%和25.19%、21.75%、15.83%;成熟期2个品种T1处理的木质素S型单体含量平均值分别较CK、T2、T3处理高18.82%、18.48%、8.39%。不同追氮模式处理的木质素合成相关酶基因(苯丙氨酸解氨酶:PAL、咖啡酸3氧甲基转移酶:COMT、香豆酸-3-羟基氧化酶:C3H、肉桂酰辅酶 A 还原酶:CCR、肉桂酸4羟化酶:C4H等)表达均随生育进程呈下降趋势,其表达量高低依次为T1>T3>T2>CK。孕穗期追氮处理模式的千粒重高于其他处理,因T1处理可提高穗粒数以及群体有效穗数,其最终籽粒产量较高。同一时期相同处理各节间茎秆木质素积累量、单体含量均呈现为I1>I2>I3>I4>I5的趋势。高施氮量300 kg·hm-2基施1/3条件下起身与孕穗期等量二次性追氮模式较其他春季追氮模式处理显著提高了小麦开花后茎秆各节间抗折能力、木质素积累量、木质素合成途径相关酶基因的表达以及籽粒产量。因此,起身与孕穗期等量二次性追氮模式可作为黄淮海麦区高施氮量300 kg·hm-2基施1/3条件下的春季适宜追氮模式。
追氮模式;抗倒性能;木质素积累;籽粒产量;冬小麦
0 引言
【研究意义】倒伏是小麦高产的限制因子,黄淮海麦区每年约有10%的面积发生倒伏,严重的年份高达20%以上,甚至出现绝收[1-2]。合理的氮肥调控可以有效缓解个体与群体的矛盾,改善群体结构,不合理追肥易造成茎秆质量降低,群体结构变差[3-5]。氮肥合理后移能够增加基部节间壁厚、充实度及木质素积累量,从而提升茎秆的抗倒性能[4,6]。目前在生产上仍存在高产农田过量施用氮肥的问题[7-8]。因此,试验在高施氮量300 kg·hm-2基施1/3条件下设置不同的追肥时期,探究高施氮量条件下不同春季追氮模式对茎秆各节间生育后期抗倒伏能力的影响,以期为小麦高产稳产抗逆应变栽培提供理论依据。【前人研究进展】氮肥调控主要包括合理的氮肥施用量和合适的基追比两方面,是作物增产的重要途径[9]。前人研究表明,在小麦和玉米上,施足底肥,分次施肥有助于提高作物籽粒产量及茎秆强度,孕穗期追肥可以提高小麦茎秆中木质素的积累量,提高茎秆的抗折力,进而提高茎秆的抗倒伏能力[10-12]。茎秆的抗倒伏性能与植株高度、茎粗、壁厚、抗折力等生理指标密切相关,还与茎秆的解剖结构以及木质素代谢有关[13-21]。合理的氮肥运筹可以提高茎秆基部第二节间木质素积累量以及木质素合成途径关键酶的活性,且合理的氮肥施用量可以提高茎秆各节间中木质素单体的含量[4,15]。过量施氮会导致水稻茎秆基部节间中木质素、纤维素、半纤维素的含量降低,使茎秆抗折力下降,茎秆质量劣化。水稻中穗肥过量降低了木质素合成基因PAL、COMT、4CL3、CCR、CAD2、CAD7在茎秆发育早期的表达量[22-23]。木质素是植物体中重要的苯酚类代谢产物,和纤维素、半纤维素约占植物体干重的15%—40%,存在于细胞壁的多聚糖框架中以增强植物的机械强度[24-26]。木质素是由松柏醇、芥子醇和对香豆醇聚合形成的物质,在相关酶的催化作用下对应生成愈创木基木质素(G型木质素)、紫丁香基木质素(S型木质素)和对羟基苯基木质素(H型木质素)[27-29],木质素单体的含量及比例与茎秆的机械强度有密切关系,其中起主要机械支撑作用的是S型木质素[13,21,30-32]。关于木质素合成途径的分子调控前人已经进行了较多的研究,木质素合成途径呈网络状,表现在中间产物及木质素的侧基修饰类型丰富,许多产物的生成不止一条合成途径,且这些木质素合成途径相关基因均表现为多基因家族[32]。木质素中3种单体含量和比例受物种、品种、生育时期、环境因素、木质素合成相关酶基因上调、下调以及相互作用的影响[33-38]。PAL是木质素合成途径的关键起始酶,其表达丰度影响着整个木质素代谢过程,PAL基因下调时,植物体内木质素含量降低[39]。F5H或COMT基因调控均可严重影响S型木质素的生物合成[40-41],水稻中过量表达F5H增加了S型单体的含量,减少了G型单体的含量,影响水稻的抗倒伏能力[34]。【本研究切入点】关于氮肥对小麦茎秆抗倒伏能力的研究多集中于氮肥施用量,对不同抗倒伏品种小麦茎秆性能影响的研究也多集中于基部节间。生育后期受外界天气及茎秆自身变化的影响,上部节间也容易发生倒伏[42-43],不同春季追氮模式对茎秆基部第二节间茎秆抗倒伏能力的调控已有报道,但对茎秆各节间抗折力、木质素单体积累以及木质素合成相关酶基因的表达量尚无深入研究。【拟解决的关键问题】本试验以抗倒伏性能不同的2个小麦品种为材料,在总施氮量300 kg·hm-2的条件下,研究不同春季追氮模式对茎秆各节间木质素积累规律的影响及抗折力的影响,为提高小麦各节间抗倒伏能力及小麦高产稳产抗逆应变栽培提供理论依据。
1 材料与方法
1.1 试验设计
2017—2018和2018—2019年在山东农业大学农学实验站进行大田试验,试验选用倒伏敏感型品种山农16和抗倒伏品种济麦22。试验田土壤为棕壤土,前茬作物为玉米,秸秆全部还田。小麦全生育期施用纯N 300 kg·hm-2(尿素),P2O590 kg·hm-2(过磷酸钙),K2O 100 kg·hm-2(氯化钾),种植密度300万株/hm2。钾肥、磷肥和1/3的氮肥作为基肥均匀施入田中,翻耕入土。剩余氮肥后期开沟追施,设置4个追肥时期,分别为等量二次性追氮和剩余一次性追氮,即起身期﹕孕穗期1/3﹕1/3(T1)、拔节期﹕开花期1/3﹕1/3(T2)、孕穗期一次性追施2/3(T3)、拔节期一次性追施2/3(CK)。小区面积9 m2(3 m×3 m),每个处理设置3次重复,共24个小区,人工开沟播种,行距25 cm,其他管理同一般大田。
1.2 测定项目和方法
1.2.1 茎秆抗折力的测定 于小麦开花期、灌浆期、成熟期取15株生长均匀一致的植株,分别测量各节间的抗折力,使用的仪器为浙江托普仪器有限公司生产的茎秆强度测定仪(YYD-1),测量参照Peng等[18]和Zheng等[13]的方法。将小麦从顶部向下依次分为I5、I4、I3、I2、I1,把每一节间中间部分置于茎秆强度测定仪的凹槽中,支撑长度5 cm,此时茎秆与压力器探头垂直,手持压力传感器手柄匀速下压,茎秆被折断时屏幕显示的数值(单位为N)表示为该节间的抗折力。
1.2.2 木质素含量的测定 木质素含量的测定参照Syros等[27]的方法略有改动。取鲜样置于研钵中,加液氮研磨成粉状。称取0.1 g于10 ml离心管中,加入8 ml 95%乙醇过夜浸提后离心去上清。沉淀中加入8 ml正己烷﹕乙醇=2﹕1的混合溶液浸提,离心弃上清,重复2次,50℃烘干至恒重。干燥物用2.5 ml 25%乙溴酰冰醋酸(体积比)溶解,70℃水浴保温30 min,加热期间摇晃2次混匀,冷水中快速冷却至室温,然后加入0.9 mL 2 mol·l-1NaOH溶液终止反应,加0.1 ml 7.5 mol·l-1盐酸羟胺和4 ml冰乙酸,混匀后离心5 min,吸取0.1 ml上清液,加7.9 ml冰乙酸(前2次取样含量较少,可取上清液0.2 ml,加3.8 ml冰乙酸稀释20倍)稀释后测定A280nm。以单位质量鲜样在A280nm处的吸光值表示木质素含量。
1.2.3 木质素单体含量的测定 参照Zheng等[17]的方法,茎秆样品研磨过筛,经NaCl溶液、无水乙醇、95%乙醇、丙酮、色谱氯仿﹕色谱甲醇1﹕1萃取后50℃烘干备用。准确称取0.02 g上述烘干备用的样品,置于50 ml白色消解管中(微波消解仪配套白色50 ml消解管),加入3 ml 2 mol·l-1的NaOH和0.5 ml的硝基苯,混匀后放入微波消解仪中(MULTIWAVE 3000),150℃保温1 h,然后将样品转移到10 ml离心管中并加入0.5 ml 2 mol·l-1NaOH冲洗白管,冲洗后的液体转移到10 ml离心管中,5 000×离心5 min。吸取2.7 ml上清液转移到新的10 ml离心管中,加入4 ml乙酸乙酯(CNW),用振荡器漩涡混匀5 000×离心后,吸取3 ml上清转移到新的离心管中。重复加入4 ml乙酸乙酯,漩涡混匀吸取4 ml上清液,与上一步骤得到的3 ml乙酸乙酯合并。将7 ml有机相利用真空离心浓缩仪(Thermo Fisher)蒸干,蒸干后加6 ml 50%的乙腈水复溶样品,用振荡器涡旋并于超声仪中进行超声,使样品充分溶解混匀。将样品用50%的乙腈水稀释后过0.22 μm有机滤器,利用超高效液相-三重四级杆质谱仪(Xevo TQ-S,Waters,Milford,MA,USA)进行样品定性及定量的测定。
1.2.4 小麦茎秆木质素合成关键酶基因表达丰度的测定 将按照节间分好的鲜样用液氮研磨至粉末状,然后迅速转移到无RNA酶、无DNA酶、无菌的2 ml离心管中,利用全式金试剂盒提取RNA,然后用全式金反转录试剂盒反转录为cDNA。配制96孔板所需体系,每孔配制20 μl,引物序列如表1所示。将20 μl体系分别加入96孔板相应孔内(操作要缓慢,避免产生气泡),将96孔板放入ABI 7500型实时荧光定量PCR仪(赛默飞)中反应,进行目的基因的表达分析。

表1 木质素合成途径中关键基因引物及探针的序列
ACTIN:内参基因;PAL:苯丙氨酸解氨酶;C4H:肉桂酸4羟化酶;HCT:羟基肉桂酰辅酶A转移酶;C3H:香豆酸-3-羟基氧化酶;CCR:肉桂酰辅酶 A 还原酶;CAD:肉桂醇脱氢酶;4CL:香豆酸辅酶A连接酶;F5H:阿魏酸 5 羟化酶;COMT:咖啡酸3氧甲基转移酶;CSE:咖啡酰莽草酸酯酶
ACTIN: Reference gene; PAL: phenylalanine ammonia-lyase; C4H: cinnamate 4-hydroxylase; HCT: p-hydroxycinnamoyl-CoA-shikimate/quinate p-hydroxycinnamoyl transferase; C3H: Coumarate -3- hydroxyl oxidase; CCR: Cinnamoyl CoA reductase; CAD: cinnamyl alcohol dehydrogenase; 4CL: 4-coumarate CoA ligase; F5H: Ferulic acid 5-hydroxylase; COMT: caffeic acid3-o-methytransferase; CSE: Caffeoyl shikimate esterase
1.3 数据分析和处理
用DPS(Data Processing System)软件分析数据,采用最小显著极差法(LSD 0.05)进行平均数显著性检验,用Microsoft Excel软件对数据进行绘图。2年试验结果趋势一致,本文采用2018—2019基因表达丰度的数据,其他指标均为2年数据。
2 结果
2.1 不同春季追氮模式对小麦各节间抗折力的影响
从开花期到成熟期,抗倒伏品种济麦22的抗折力高于倒伏敏感型品种山农16,2个品种的抗折力呈现出先增加后降低的趋势,在灌浆期达到最大值(表2)。在高施氮量300 kg·hm-2基施1/3条件下,2种类型品种均表现出T1处理(起身期和孕穗期1﹕1等量二次性追氮模式追施氮肥)显著提高了小麦茎秆各节间的抗折力。开花期2种类型品种T1、CK处理抗折力高于T2、T3处理,在灌浆期和成熟期呈现T1>T3>T2>CK的规律,不同春季追氮模式对小麦茎秆抗折力的影响在2个生长季表现一致。以2017—2018年为例,灌浆期山农16和济麦22第二节间T1处理分别较CK、T2、T3增加了24.69%、19.97%、13.15%和26.92%、15.36%、5.87%,其他节间抗折力表现趋势与第二节间一致。说明将追肥时期调整为起身期与孕穗期1﹕1等量二次性追氮模式有利于提高小麦茎秆各节间的抗折力。
比较2种类型品种不同节间的抗折力,从I1—I5呈现逐渐下降的趋势。以2017—2018年灌浆期T1处理的5个节间为例,与I1相比,山农16 I2、I3、I4、I5抗折力分别下降了11.19%、35.49%、45.95%、73.38%,济麦22抗折力分别降低了15.66%、42.12%、56.86%、103.91%。不同节间之间抗折力差异显著,上部节间抗折力显著小于下部节间,在生育后期受恶劣天气的影响更容易发生倒伏,因此上部节发生倒伏的可能性同样不可忽视。

表2 不同春季追氮模式对不同抗倒伏型小麦品种各节间抗折力的影响
SN16表示山农16,JM22表示济麦22。CK表示拔节期一次性追施2/3,T1表示起身期﹕孕穗期等量二次性追施2/3,T2表示拔节期﹕开花期等量二次性追施2/3,T3表示孕穗期一次性追施2/3。茎秆从基部向上依次进行标记为I1、I2、I3、I4、I5。同一年份同列数据后不同字母表示差异达5%显著水平。下同
SN16 and JM22 represent Shannong16 and Jimai22, respectively. CK represents the jointing stage 2/3 which is remaining one-time nitrogen topdressing. TI represents the rising stage﹕booting stage 1/3﹕1/3, which is equal amount of secondary nitrogen topdressing. T2 represents the jointing stage﹕flowering stage 1/3﹕1/3, which is equal amount of secondary nitrogen topdressing. T3 represents booting stage 2/3, which is remaining one-time nitrogen topdressing. The stems are labeled I1, I2, I3, I4 and I5 from the base to top. Different letters of a column in the same year are significantly different at<0.05 level. the same as below
2.2 不同春季追氮模式对小麦茎秆木质素积累量的影响
不同春季追氮模式对小麦茎秆木质素的积累具有调控作用。抗倒伏品种济麦22各时期木质素积累量高于山农16,在不同生育时期,木质素的积累量呈现显著差异。随着生育进程推进,木质素总积累量增加,各生育阶段木质素积累量呈逐渐减小的趋势(图1)。不同处理下,拔节期至开花期木质素积累量表现为T1>T3>CK>T2,开花期到灌浆期及灌浆期到成熟期木质素积累量表现为T1>T3>T2>CK,2年结果趋于一致。以2018—2019年第二节间为例,拔节期至开花期山农16和济麦22在T1处理下木质素积累量分别比CK、T2、T3处理高出13.43%、30.69%、12.88%和30.79%、38.33%、29.47%,开花期至灌浆期山农16和济麦22在T1处理下木质素积累量分别比CK、T2、T3处理高出28.58%、0.27%、4.71%和22.11%、15.97%、16.80%,灌浆期至成熟期山农16和济麦22在T1处理下木质素积累量分别比CK、T2、T3处理高出23.11%、15.38%、8.96%和22.66%、10.97%、1.23%。说明起身期﹕孕穗期等量二次性追肥提高了茎秆后期各生育阶段木质素积累量。
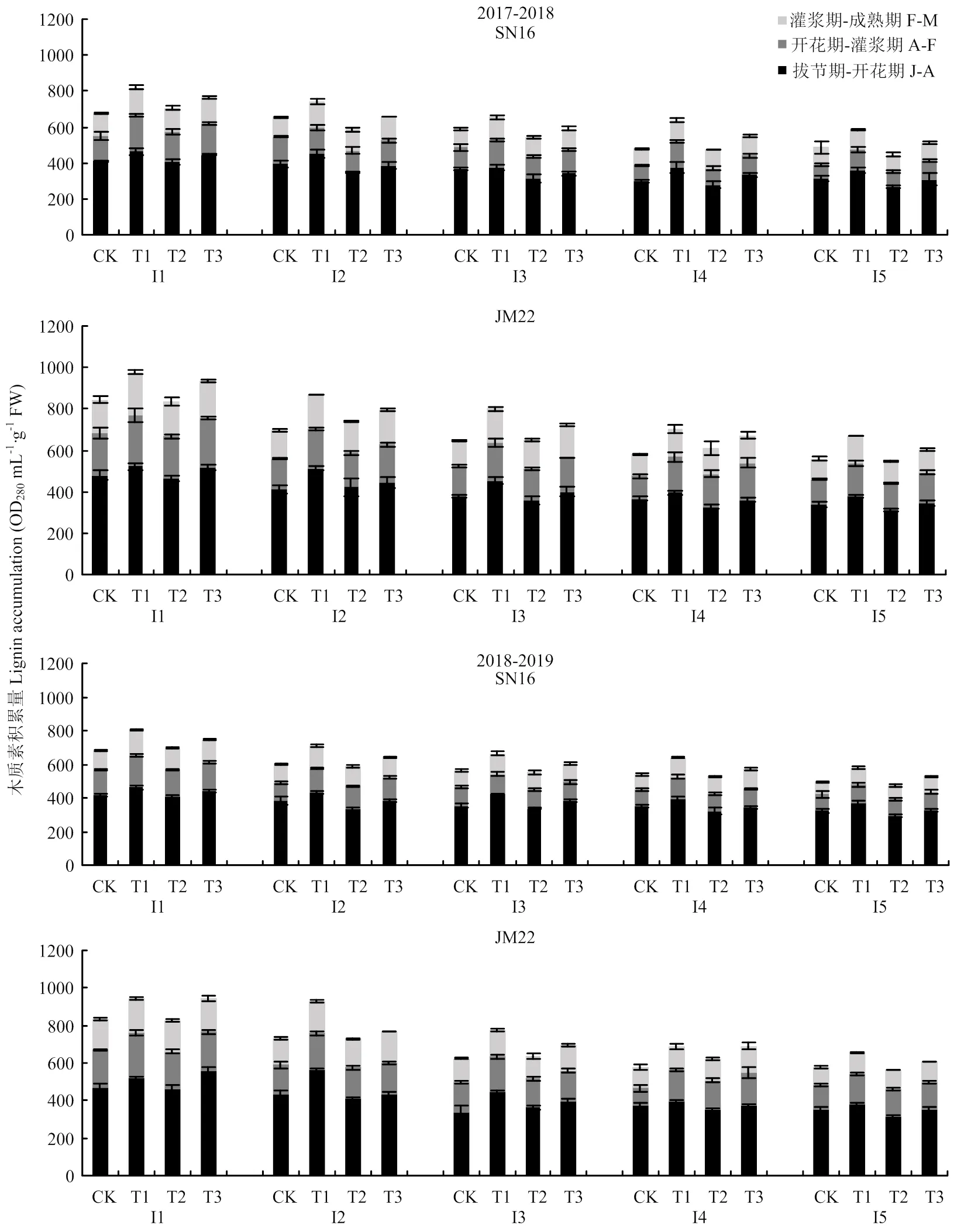
SN16表示山农16,JM22表示济麦22。J表示拔节期,A表示开花期,F表示灌浆期,M表示成熟期。下同
相同处理下,不同节间之间存在差异,基部节间木质素积累量要高于上部节间,在2018—2019年T1处理中,山农16第一节间3个生育时期均值较第二节间、第三节间、第四节间、第五节间分别高出18.46%、30.63%、32.10%、46.53%。JM22第一节间3个生育时期均值较其他节间高出7.86%、24.01%、42.14%、49.51%。不同处理之间节间变化趋势与T1处理变化规律一致。3个生育阶段各节间木质素的积累量随生育期呈现降低的趋势。
2.3 不同春季追氮模式对小麦茎秆木质素单体含量及积累规律的影响
木质素是小麦茎秆的重要组成成分,由愈创木基木质素(G型)、紫丁香基木质素(S型)和对羟基苯基木质素(H型)3种单体组成。由图2可知,在小麦中G型、S型单体的含量显著高于H型,抗倒伏品种济麦22 3种单体的总含量显著高于倒伏敏感型品种山农16。开花期2种类型品种不同处理间S型单体含量的变化趋势表现为T1>T3>CK>T2,灌浆期和成熟期表现为T1>T3>T2>CK。随着生育进程推进,3种单体含量的总和逐渐增加,S型/G型的比例逐渐减少,S型单体含量占3种单体含量总和的31.05%—49.80%,G型单体含量占3种单体含量总和的41.94%—53.21%,G型+S型单体含量占3种单体含量总和的78.05%—93.31%(以2018—2019年数据为例)。不同春季追氮模式对小麦茎秆木质素含量及其积累规律有一定影响,以2018—2019年成熟期第二节间为例,山农16 T1处理S型单体含量分别比CK、T2、T3处理增加了13.01%、25.62%、5.55%,G型单体含量分别比CK、T2、T3处理增加了17.18%、18.01%、1.24%,JM22 T1处理S型单体含量分别比CK、T2、T3处理增加了24.62%、11.34%、11.22%,G型单体含量分别比CK、T2、T3处理增加了14.38%、9.67%、3.03%。追肥模式以起身期﹕孕穗期1﹕1等量二次性追氮模式显著增加了茎秆生育后期各节间G型、S型、H型单体的含量,拔节期全部追施不利于生育后期木质素单体的积累。
各节间变化规律趋于一致,从开花期到成熟期G型、S型、H型单体的含量逐渐增大。从I1至I5 S型、G型单体的含量逐渐下降,3种单体的总含量也逐渐下降。2018—2019年山农16成熟期第一节间S型单体含量较第二节间、第三节间、第四节间、第五节间分别高出13.71%、39.56%、45.72%、61.26%,G型单体含量较其他节间分别高出2.73%、10.95%、28.67%、54.14%。不同处理间各节间变化趋势一致,说明起身期﹕孕穗期1﹕1等量二次性追施氮肥有利于提高高氮肥处理下小麦生育后期茎秆各节间木质素单体的积累量,孕穗期一次性追氮也有利于生育后期茎秆各节间木质素3种类型单体的积累。
茎秆抗折力与木质素积累量、S型单体含量以及S型+G型单体含量呈极显著正相关(表3)。2018—2019年成熟期抗折力与木质素积累量以及S型单体含量、S型+G型单体含量相关系数分别为0.85、0.70和0.66,结果表明抗折力高的品种茎秆中木质素积累量以及S型单体含量、S型+G型单体含量较高。本试验中,T1处理茎秆抗折力优于其他处理,说明在高施氮量300 kg·hm-2基施1/3条件下可以通过起身期﹕孕穗期1﹕1等量二次性追施提高茎秆中木质素积累量、S型单体含量及G型+S型单体的含量进而提高茎秆的抗倒伏能力。
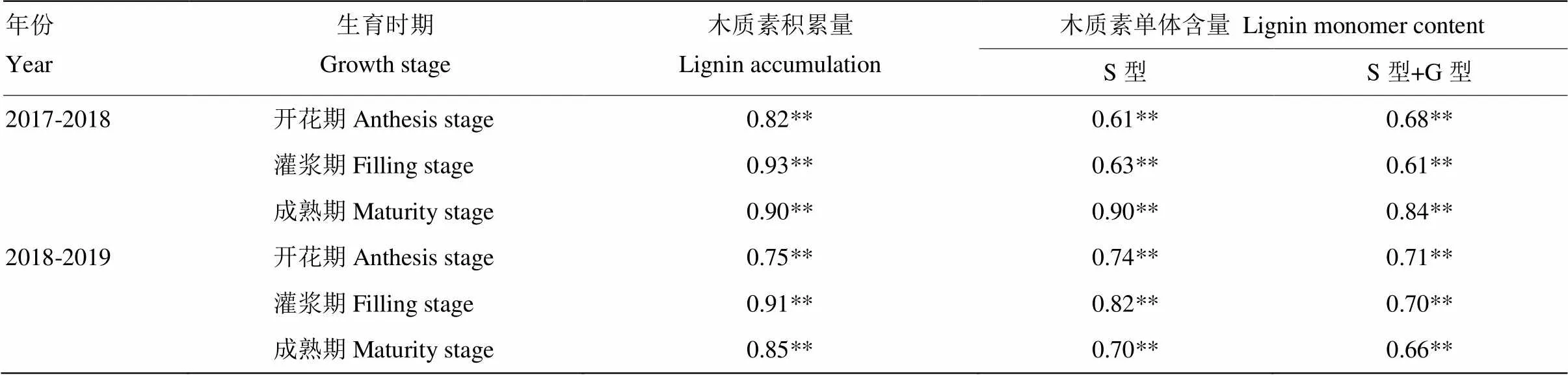
表3 抗折力与木质素积累的相关分析
*表示<0.05的显著水平,**表示<0.01的显著水平。下同 *and **represent significant at 0.05 and 0.01 probability levels, respectively. The same as below

2.4 不同春季追氮模式对小麦茎秆木质素合成相关酶基因表达丰度的影响
苯丙氨酸解氨酶PAL是木质素合成途径的关键起始酶。由图3可知,木质素合成相关酶基因苯丙氨酸解氨酶(PAL)、肉桂酸4羟化酶(C4H)、羟基肉桂酰辅酶A转移酶(HCT)、香豆酸-3-羟基氧化酶(C3H)、肉桂酰辅酶 A 还原酶(CCR)、肉桂醇脱氢酶(CAD)、香豆酸辅酶A连接酶(4CL)、阿魏酸 5 羟化酶(F5H)、咖啡酸3氧甲基转移酶(COMT)、咖啡酰莽草酸酯酶(CSE)在抗倒伏品种济麦22中基因表达丰度高于倒伏敏感型品种山农16。随着生育进程的推进,PAL、C3H、C4H、COMT、F5H等木质素合成相关酶基因的表达丰度呈逐渐降低的趋势。开花期第二节间不同处理间表现为T1>CK>T3>T2,开花后7 d及之后的时期,不同处理间表现为T1>T3>T2>CK。说明在本试验追肥条件下,将追肥时期改为起身肥﹕孕穗期1﹕1等量二次性追施可以提高开花期及之后时期茎秆木质素合成相关酶基因的表达丰度,拔节期一次性追施以及过晚施肥均不利于茎秆中木质素合成途径相关酶基因的表达,进而影响茎秆中木质素单体的含量和抗折力,最终影响茎秆质量。
不同节间木质素合成途径相关酶基因的表达丰度呈现I5>I4>I3>I2>I1的趋势。相同生育时期其余节间木质素合成途径相关酶基因的表达丰度与第二节间开花期后木质素合成途径相关酶的基因表达丰度变化趋势基本一致。结果表明,不同春季追氮模式对茎秆各节间木质素合成相关酶基因的表达有调控作用,起身期﹕孕穗期1﹕1等量二次性追施可以提高各节间开花期及之后生育时期木质素合成相关酶基因的表达。
2.5 不同春季追氮模式对小麦籽粒产量和产量构成因素的影响
山农16籽粒产量呈现T1>T2>T3>CK的趋势,济麦22籽粒产量呈现出T1>T2>CK>T3的趋势,2种类型品种均以T1处理籽粒产量最高,山农16在T1处理下2年籽粒产量平均比CK、T2、T3高7.47%、4.37%、5.49%。济麦22在T1处理下2年籽粒产量平均比其他处理高9.97%、6.58%、12.59%(表4)。CK、T3处理产量差异不显著,说明过晚追肥、一次性追肥均不利于籽粒产量的提高。不同处理之间,以T3处理千粒重最高,说明孕穗期追肥可以显著提高籽粒的产量。虽然T3处理千粒重较高,但由于追肥时期过晚,植株成穗数少,导致T3籽粒产量低于T1。拔节期一次性追氮模式虽可保证足够穗数,但不利于提高茎秆抗折能力。

表4 不同春季追氮模式对不同抗倒伏能力小麦籽粒产量的影响

5月3日标为开花后0 d,7 DAA、14 DAA、21 DAA分别表示开花后7 d,14 d,21 d
由表5可知,对2018—2019年开花期、灌浆期、成熟期茎秆基部第二节间茎秆抗折力、木质素积累量等抗倒伏指标与籽粒产量进行相关分析,籽粒产量与木质素积累量以及S型单体含量呈极显著正相关(籽粒产量与开花期S型单体含量呈显著正相关),籽粒产量与成熟期茎秆抗折力、木质素积累量和S型单体的含量呈极显著正相关,相关指数分别为0.69、0.91和0.81,与G型单体含量、S型+G型单体的含量呈显著正相关,相关系数为0.75和0.77。说明合理的栽培措施不仅可以提高茎秆的质量,籽粒产量也会增加。

表5 茎秆抗倒伏指标与籽粒产量指标的相关分析(2018—2019)
3 讨论
3.1 不同春季追氮模式对小麦茎秆抗倒性能的影响
优化的氮肥施用模式(基肥﹕拔节肥﹕孕穗肥4﹕4﹕2)可以提高茎秆中木质素合成关键酶PAL、TAL、POD的活性以及木质素积累量,提高茎秆抗折力,最终提高茎秆抗倒伏能力[4]。施氮量过高导致茎秆壁厚、充实度降低,大小维管束数目及木质素积累量减少,茎秆抗折力降低,在生长发育后期易引发倒伏[15,23]。本试验探讨了高施氮量300 kg·hm-2且基施1/3条件下不同春季追氮模式对茎秆各节间抗折力的影响,不同处理间茎秆抗折力以起身肥﹕孕穗肥等量二次性追施茎秆抗折力优于其他处理,且2种类型品种抗折力随生育时期均呈现先增高后降低的趋势,在灌浆期达到最大值。S型单体主要为茎秆提供机械强度支撑[13,15]。在本试验条件下,S型单体的含量在2年生长季的不同生育时期均表现为起身期﹕孕穗期1﹕1等量二次性追施显著高于其他处理。开花期木质素各生育阶段积累量、单体的含量呈现T1>T3>CK>T2的规律,灌浆期和成熟期呈现T1>T3>T2>CK的趋势。起身期﹕孕穗期等量二次性追施(T1)提高了高氮肥种植条件下小麦茎秆各节间木质素合成相关酶基因的表达量、各生育阶段木质素积累量及单体的含量,说明起身肥﹕孕穗肥等量二次性追施通过提高各节间木质素合成途径关键酶的表达丰度,进而提高了茎秆各节间木质素的积累量,最终提高了各节间茎秆抗折力。
茎秆质量和木质素代谢研究表明,木质素的含量与茎秆的抗倒伏能力密切相关,抗倒伏强的品种木质素含量较高[4,17]。在本研究中,相同处理间抗倒伏品种济麦22抗折力、木质素的积累量、单体的含量较倒伏敏感型品种山农16高。倒伏一般发生在小麦生长发育的中后期[5]。本研究探究了从开花期到成熟期,茎秆各节间木质素单体的含量、木质素总量等指标随生育时期的变化。木质素合成相关酶基因的表达量随着生育时期呈现降低趋势,木质素总积累量随生育时期增加,但积累速率逐渐降低。3种木质素单体S型、G型、H型的含量随生育时期逐渐增加,S型/G型单体的值减小,这与抗折力随生育时期进程的变化规律不一致。本研究还发现,2个生长季茎秆各节间抗折力与木质素积累量、S型单体含量以及S型+G型单体含量均呈显著正相关。结合前人研究结果,高施氮量300 kg·hm-2且基施1/3条件下,在小麦不同生育时期可以通过合理追肥定向调控茎秆的机械强度,将追肥时期改为起身期﹕孕穗期1﹕1等量二次性追施,提高茎秆各节间木质素合成关键酶基因的表达以及木质素单体的积累,以达到提高茎秆抗倒伏能力的目的。不同节间之间,茎秆基部节间短而充实,上部节间中空细长,且上部节间拔节时间晚,木质素积累量小于下部节间。从I1—I5茎秆抗折力、木质素积累量及单体的含量逐渐减少,木质素合成相关酶各节间表达规律与上述指标相反,从I1—I5相关酶基因的表达量逐渐增加,推测是由于上部节间拔节较晚,茎秆组织幼嫩,相关酶基因的表达量较高。
3.2 不同春季追氮模式对冬小麦籽粒产量的影响
在实际大田生产中,兼顾籽粒产量和茎秆质量一直是研究的重点[14-15,18]。小麦籽粒产量及构成因素受多种条件的影响,包括氮肥施入量、肥料利用率、施用时期以及氮肥基追比例[4,6,9-10]。合适的栽培措施可以保证茎秆质量,增加籽粒产量,且适宜的追肥时期可显著增加茎秆中的单糖、多糖的含量,提高茎秆充实度,促使纤维素及木质素的形成,提高抗折力,改善其抗倒伏性能[44-46]。本研究表明,起身期、拔节期追肥有助于提高小麦分蘖成穗率,孕穗肥有助于显著提高千粒重,减缓后期木质素积累速率的降低,增加木质素单体的含量。本研究还发现,从开花期到成熟期,籽粒产量与木质素积累量以及S型单体含量呈极显著正相关(籽粒产量与开花期S型单体含量呈显著正相关),说明在增强茎秆抗倒伏能力的同时,籽粒产量也能得到提高。结合前人分析可得,在高施氮量300 kg·hm-2基施1/3情况下,在小麦生育后期以起身期﹕孕穗期1﹕1等量二次性追氮可以提高茎秆中木质素总积累量及木质素单体的积累,有助于提高茎秆抗折力,增加籽粒产量。
4 结论
本试验条件下,基肥﹕起身肥﹕孕穗肥为1﹕1﹕1施用时较其他处理显著提高了小麦生育后期茎秆各节间的抗折力、木质素积累量、单体含量、木质素合成途径相关酶基因的表达以及产量。开花期、灌浆期、成熟期茎秆抗折力与木质素积累量、S型单体含量、S型+G型单体含量存在极显著正相关关系,说明抗倒伏品种具有更高的木质素合成相关基因的表达量、木质素积累量和木质素单体含量。起身期:孕穗期1﹕1等量二次性追施既可以保证单位面积成穗数,又能提高粒重,进而获得高产。拔节期一次性追肥虽然可以保证足够的穗数,但不利于茎秆各节间抗折力和产量的提高。因此,在高施氮量300 kg·hm-2且基施1/3条件下,起身期与孕穗期1﹕1等量二次性追氮模式可作为黄淮海麦区高施氮量300 kg·hm-2基施1/3条件下既能保证高产又能提高生育后期茎秆质量的春季适宜追氮模式。
[1] 李金才, 尹钧, 魏凤珍. 播种密度对冬小麦茎秆形态特征和抗倒指数的影响. 作物学报, 2005, 31(5): 662-666.
LI J C, YIN J, WEI F Z. Effects of planting density on characters of culm and culm lodging resistant index in winter wheat.,2005, 31(5): 662-666. (in Chinese)
[2] 魏凤珍, 李金才, 王成雨, 屈会娟, 沈学善. 氮肥运筹模式对小麦茎秆抗倒性能的影响. 作物学报, 2008, 34(6): 1080-1085.
WEI F Z, LI J C, WANG C Y, QU H J, SHEN X S. Effects of nitrogenous fertilizer application model on culm lodging resistance in winter wheat., 2008, 34(6): 1080-1085. (in Chinese)
[3] 边大红, 刘梦星, 牛海峰, 魏钟博, 杜雄, 崔彦宏. 施氮时期对黄淮海平原夏玉米茎秆发育及倒伏的影响. 中国农业科学, 2017, 50(12): 2294-2304.
BIAN D H, LIU M X, NIU H F, WEI Z B, DU X, CUI Y H. Effects of nitrogen application times on stem traits and lodging of summer maize (L.) in the Huang-Huai-Hai plain.,2017, 50(12): 2294-2304. (in Chinese)
[4] CHEN X G, WANG J, WANG Z L, LI W Q, WNAG C Y, YAN S H, LI H M, ZHANG A J, TANG Z H, WEI M. Optimized nitrogen fertilizer application mode increased culms lignin accumulation and lodging resistance in culms of winter wheat., 2018, 228: 31-38.
[5] 安志超, 黄玉芳, 赵亚南, 汪洋, 刘小宁, 叶优良. 植株氮营养状况与冬小麦倒伏的关系. 植物营养与肥料学报, 2018, 24(3): 751-757.
AN Z C, HUANG Y F, ZHAO Y N, WANG Y, LIU X N, YE Y L. Relationship between plant nitrogen nutrition and lodging of winter wheat.,2018, 24(3): 751-757. (in Chinese)
[6] 张明伟, 马泉, 丁锦峰, 李春燕, 朱新开, 封超年, 郭文善. 密度与肥料运筹对迟播小麦产量和茎秆抗倒能力的影响. 麦类作物学报, 2018, 38(5): 584-592.
ZHANG M W, MA Q, DING J F, LI C Y, ZHU X K, FENG C N, GUO W S. Effect of density and nitrogen applicationon clum lodging resistance and yield of late sowing wheat., 2018, 38(5): 584-592. (in Chinese)
[7] 张福锁, 王激清, 张卫峰, 崔振岭, 马文奇, 陈新平, 江荣风. 中国主要粮食作物肥料利用率现状与提高途径. 土壤学报, 2008, 45(5): 915-924.
ZHANG F S, WANG J Q, ZHANG W F, CUI Z L, MA W Q, CHEN X P, JIANG R F. Nutrient use efficiencies of major cereal crops in China and measures for improvement., 2008, 45(5): 915-924. (in Chinese)
[8] 赵风华, 马军花, 欧阳竹. 过量施氮对冬小麦生产力的影响.植物生态学报, 2012, 36(10): 1075-1081.
ZHAO F H, MA J H, OUYANG Z. Effects of excessive nitrogen supply on productivity of winter wheat., 2012, 36(10): 1075-1081. (in Chinese)
[9] 周洁, 王旭, 朱玉磊, 刘惠惠, 陈翔, 魏凤珍, 孙建强, 宋有洪, 李金才. 氮肥运筹模式对小麦茎秆抗倒性能与产量的影响. 麦类作物学报, 2019, 39(8): 979-987.
ZHOU J, WANG X, ZHU Y L, LIU H H, CHEN X, WEI F Z, SUN J Q, SONG Y H, LI J C. Effects of nitrogen fertilizer management on stem lodging resistance and yield of wheat., 2019, 39(8): 979-987. (in Chinese)
[10] 卢昆丽, 尹燕枰, 王振林, 李勇, 彭佃亮, 杨卫兵, 崔正勇, 杨东清, 江文文. 施氮期对小麦茎秆木质素合成的影响及其抗倒伏生理机制. 作物学报, 2014, 40(9): 1686-1694.
LU K L, YIN Y P, WANG Z L, LI Y, PENG D L, YANG W B, CUI Z Y, YANG D Q, JIANG W W. Effect of nitrogen fertilization timing on lignin synthesis of stem and physiological mechanism of lodging resistance in wheat., 2014, 40(9): 1686-1694. (in Chinese)
[11] 张明伟, 易媛, 董召娣, 柯裴蓓, 朱新开, 封超年, 郭文善, 彭永欣. 氮肥对扬麦20茎秆性状和抗倒性能的影响. 麦类作物学报, 2014, 34(9): 1260-1266.
ZHANG M W, YI Y, DONG Z D, KE P P, ZHU X K, FENG C N, GUO W S, PENG Y X. Effects of nitrogen on internode traits and lodging resistance of wheat variety Yangmai 20.,2014, 34(9): 1260-1266. (in Chinese)
[12] 周羊梅, 顾正中, 王安邦, 杨子博, 冷苏凤. 播期、密度和不同施氮时期对高产品种‘淮麦33’产量和品质的调控. 中国农学通报, 2019, 35(19): 1-5.
ZHOU Y M, GU Z Z, WANG A B, YANG Z B, LENG S F. Effect of sowing date, density and nitrogen management on grain yield and quality of high yield ‘Huaimai 33’., 2019, 35(19): 1-5. (in Chinese)
[13] Zheng M J, Chen J, Shi Y H, Li Y X, Yin Y P, Yang D Q, Luo Y L, Pang D W, XU X, Li W Q, Ni J, Wang Y Y, Wang Z L, Li Y. Manipulation of lignin metabolism by plant densities and its relationship with lodging resistance in wheat., 2017, 7(1): 41805.
[14] Xu C L, GAO Y B, TIAN B J, REN J H, MENG Q F, WANG P. Effects of EDAH, a novel plant growth regulator, on mechanical strength, stalk vascular bundles and grain yield of summer maize at high densities.,2017, 200: 71-79.
[15] Luo Y L, NI J, PANG D W, JIN M, CHEN J, KONG X, LI W Q, CHANG Y L, LI Y, WANG Z L.Regulation of lignin composition by nitrogen rate and density and its relationship with stem mechanical strength of wheat., 2019 , 241: 107572.
[16] KAMRAN M, AHMAD I, WANG H Q, WU X R, XU J, LIU T N, DING R X, HAN Q F. Mepiquat chloride application increases lodging resistance of maize by enhancing stem physical strength and lignin biosynthesis.,2018, 224: 148-159.
[17] ZHENG M J, GU S B, CHEN J, LUO Y L, LI W Q, NI J, LI Y,WANG Z L. Development and validation of a sensitive UPLC-MS/MS instrumentation and alkaline nitrobenzene oxidation method for the determination of lignin monomers in wheat straw., 2017, 1055-1056: 178-184.
[18] Peng D L, Chen X G, Yin Y P, Lu K L, Yang W B, Tang Y H, Wang Z L. Lodging resistance of winter wheat (L.): Lignin accumulation and its related enzymes activities due to the application of paclobutrazol or gibberellin acid., 2014, 157: 1-7.
[19] 陈晓光, 史春余, 尹燕枰, 王振林, 石玉华, 彭佃亮, 倪英丽, 蔡铁. 小麦茎秆木质素代谢及其与抗倒性的关系.作物学报, 2011, 37(9): 1616-1622 .
Chen X G, Shi C Y, Yin Y P, Wang Z L, Shi Y H, Peng D L, Ni Y L, Cai T. Relationship between lignin metabolism and lodging resistance in wheat., 2011, 37(9): 1616-1622. (in Chinese)
[20] 董琦, 王爱萍, 梁素明. 小麦基部茎节形态结构特征与抗倒性的研究. 山西农业大学学报, 2003, 23(3): 188-191.
DONG Q, WANG A P, LIANG S M. Study on the architectural characteristics of wheat stalks., 2003, 23(3): 188-191. (in Chinese)
[21] RAGAUSKAS A J, BECKHAM G T, BIDDY M J, CHANDRA R, Chen F, DAVIS M F, DAVISON B H, DIXON R A, GILNA P, KELLER M, LANGAN P, NASKAR A K, SADDLER J N, TSCHAPLINSKI T J, TUSKAN G A, WYMAN C E. Lignin valorization: improving lignin processing in the biorefinery., 2014, 344(6185): 1246843.
[22] 蒋明金, 王海月, 何艳, 王春雨, 李娜, 杨志远, 孙永健, 马均. 氮肥管理对直播杂交水稻抗倒伏能力的影响. 核农学报, 2020, 34(1): 157-168.
JIANG M J, WANG H Y, HE Y, WANG C Y, LI N, YANG Z Y, SUN Y J, MA J. Effect of nitrogen fertilizer management on lodging resistance of direct-seeding hybrid rice., 2020, 34(1): 157-168. (in Chinese)
[23] ZHANG W J, WU L M, WU X R, DING Y F, LI G H, LI J Y, WENG F, LIU Z H, TANG S, DING C Q, WANG S H. Lodging resistance of japonica rice (L.): Morphological and anatomical traits due to top-dressing nitrogen application rates., 2016, 9(1): 31.
[24] MIAO Y C, LIU C J. ATP-binding cassette-like transporters are involved in the transport of lignin percursors across plasma and vacuolar membranes., 2010, 107(52): 22728-22733.
[25] ALEJANDRO S, LEE Y, TOHGE T, SUDRE D, OSORIO S, PARK J, BOVET L, LEE Y, GELDNER N, FERNIE A R, MARTINOIA E. AtABCG29 is a monolignol transporter involved in lignin biosynthesis., 2012, 22(13): 1207-1212.
[26] 任佰朝, 李利利, 董树亭, 刘鹏, 赵斌, 杨今胜, 王丁波, 张吉旺. 种植密度对不同株高夏玉米品种茎秆性状与抗倒伏能力的影响. 作物学报, 2016, 42(12): 1864-1872.
REN B Z, LI L L, DONG S T, LIU P, ZHAO B, YANG J S, WANG D B, ZHANG J W. Effects of plant density on stem traits and lodging resistance of summer maize hybrids with different plant heights.,2016, 42(12): 1864-1872. (in Chinese)
[27] SYROS T, YUPSANIS T, ZAFIRIADIS H, ECONOMOU A. Activity and isoforms of peroxidases, lignin and anatomy, during adventitious rooting in cuttings ofL..,2004, 161(1): 69-77.
[28] BOERJAN W, RALPH J, BAUCHER M. Lignin biosynthesis.2003, 54(1): 519-546.
[29] VANHOLME R, DEMEDTS B, MORREEL K, RALPH J, BOERJAN W. Lignin biosynthesis and structure., 2010, 153(3): 895-905.
[30] 章霄云, 郭安平, 贺立卡, 孔祥. 木质素生物合成及其基因调控的研究进展. 分子植物种, 2006, 4(3): 431-437.
ZHANG X Y, GUO A P, HE L K, KONG X. Advances in study of lignin biosynthesis and its genetic manipulation.,2006, 4(3): 431-437. (in Chinese)
[31] ANDERSON N A, TOBIMATSU Y, CIESIELSKI P N, XIMENES E, RALPH J, DONORHOE B S, LADISCH M, CHAPPLE C. Manipulation of guaiacyl and syring monomer biosynthesis in ancinnamyl alcohol dehydrogenase mutant results in atypical lignin biosynthesis and modifed cell wall structure., 2015, 27(8): 2195-2209.
[32] 刘希强, 张涵, 龚攀, 宫文龙, 王赞. 紫花苜蓿不同发育时期次生壁合成调控的转录组分析. 中国农业科学, 2018, 51(11): 2049-2059.
LI X Q, ZHANG H, GONG P, GONG W L, WANG Z. Transcriptome analysis of secondary cell wall synthesis regulation at different developmental stages in alfalfa (L.)., 2018, 51(11): 2049-2059. (in Chinese)
[33] VOELKER S L, LACHENBRUCH B, MEINZER F C, STRAUSS H S. Reduced wood stiffness and strength, and altered stem form, in young antisense 4CL transgenic poplars with reduced lignin contents., 2011, 189(4): 1096-1109.
[34] TAKEDA Y, KOSHIBA T, TOBIMATSU Y, SUZUKI S, MURAKAMI S, YAMAMURA M, RAHMAN M M, TAKANO T, HATTORI T, SAKAMOTOO M, UMEZAWA T. Regulation ofexpression to modulate cell wall lignin structure in rice., 2017, 246(2): 337-349.
[35] Wu Z Y, Wang N F, Cao Y P, Liu W W , Bao Y, Fu C X. Simultaneous regulation of F5H in COMT-RNAi transgenic switchgrass alters effects of COMT suppression on syringyl lignin biosynthesis., 2019, 17(4): 836-845.
[36] SHAFRIN F, DAS S S, SANAN M N, KHAN H. Artificial miRNA-mediated down-regulation of two monolignoid biosynthetic genes (C3H and F5H) cause reduction in lignin content in jute., 2015, 89(4-5): 511-527.
[37] SYKES R W, GJERSING E L, FOUTZ K, ROTTMANN W H. KUHN S A, FOSTER C E, ZIEBELL A, TURNER G B. DECKER S R, HINCHEE M A W, DAVIS M F. Down-regulation of-coumaroyl quinate/shikimate 3′-hydroxylase (C3’H) and cinnamate 4-hydroxylase (C4H) genes in the lignin biosynthetic pathway ofleads to improved sugar release., 2015, 8(1): 128-137.
[38] 黄杰恒, 李威, 曲存民, 刘列钊, 徐新福, 王瑞, 李加纳. 甘蓝型油菜不同抗倒性材料中木质素代谢途径关键基因表达特点. 作物学报, 2013, 39(8): 1339-1344.
HUANG J H, LI W, QU C M, LIU L Z, XU X F, WANG R, LI J N. Expression characteristics of key genes in lignin pathway among different lodging resistance lines ofL., 2013, 39(8): 1339-1344. (in Chinese)
[39] SHI R, YANG C M, LU S F, SEDEROFF R, CHIANG V L. Specific downregulation of PAL genes by artificial micro RNAs in Populustrichocarpa., 2010,232 (6): 1281-1288.
[40] BAXTER H L, MAZAREI M, LABBE N, KLINE L M, CHENG Q, WINDHAM M T, MANN D G, FU C X, ZIEBELL A, SYKES R W, RODRIGUEZ M, DAVIS M F, MIELENZ J R, DIXON R A, WANG Z Y, STEWART C N. Two-year field analysis of reduced recalcitrance transgenic switchgrass., 2015, 12(7): 914-924.
[41] CHEN F, DIXON R A. Lignin modification improves fermentable sugar yields for biofuel production., 2007, 25(7): 759-761.
[42] ISHIMARU K, TOGAWA E, OOKAWA T, KASHIWAGI T, MADOKA Y, HIROTSU N. New target for rice lodging resistant and its effect in a typhoon., 2008, 227(3): 601-609.
[43] BERRY P M, SPINK J. Predicting yield losses caused by lodging in wheat., 2012, 137(3):19-26.
[44] 陈晓光, 石玉华, 王成雨, 尹燕枰, 宁堂原, 史春余, 李勇, 王振林. 氮肥和多效唑对小麦茎秆木质素合成的影响及其与抗倒伏性的关系. 中国农业科学, 2011, 44(17): 3529-3536.
CHEN X G, SHI Y H, WANG C Y, YIN Y P, NING T Y, SHI C Y, LI Y, WANG Z L. Effects of nitrogen and PP333 application on the lignin synthesis of stem in relation to lodging resistance of wheat., 2011, 44(17): 3529-3536. (in Chinese)
[45] 杨世民, 谢力, 郑顺林, 李静, 袁继超. 氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响. 作物学报, 2009, 35(1): 93-103.
YANG S M, XIE L, ZHENG S L, LI J, YUAN J C. Effects of nitrogen rate and transplanting density on physical and chemical characteristics and lodging resistance of culms in hybrid rice., 2009, 35(1): 93-103. (in Chinese)
[46] AHMAD I, MENG X P, KAMRAN M, ALI S, AHMAD S, LIU T N, CAI T, HAN Q F. Effects of uniconazole with or without micronutrient on the lignin biosynthesis, lodging resistance, and winter wheat production in semiarid regions., 2020, 19(1): 62-77.
Effects of Different Spring Nitrogen Topdressing Modes on Lodging Resistance and Lignin Accumulation of Winter Wheat
DONG HeHe, LUO YongLi, LI WenQian, WANG YuanYuan, ZHANG QiuXia, CHEN Jin, JIN Min, LI Yong, WANG ZhenLin
(College of Agronomy, Shandong Agricultural University/State Key Laboratory of Crop Biology, Tai’an 271018, Shandong)
【】The purpose of this experiment was to explore the effects of different spring nitrogen topdressing modes on stem lodging resistance, lignin accumulation, grain yield of winter wheat, and to identify the appropriate spring nitrogen topdressing modes under the condition of high nitrogen application, so as to provide technical support for high and stable yield and stress-resistant cultivation of winter wheat.【】In the two wheat growing seasons of 2017-2018 and 2018-2019, the lodging sensitive variety Shannong 16 and the lodging resistant variety Jimai 22 were used as test materials, and the application rate was 1/3 under high nitrogen application rate of 300 kg·hm-2. There were four types of spring topdressing modes, which were equal amount of secondary nitrogen topdressing and remaining one-time nitrogen topdressing, namely the rising stage﹕booting stage 1/3﹕1/3 (T1), jointing stage: flowering stage 1/3﹕1/3 (T2), the remaining 2/3 nitrogen was applied at the booting stage (T3) and the remaining 2/3 nitrogen was applied at the jointing stage (CK). The effects of different nitrogen topdressing modes on stem resistance, lignin accumulation, expressive abundance of the key genes involving in lignin biosynthesis pathway and grain yield of winter wheat were studied.【】The total lignin accumulation and lignin monomers content of the lodging resistance wheat were both higher than those of the lodging sensitive wheat. The breaking strength under T1, CK was higher than that under T2 and T3, the lignin accumulation and monomer content were T1>T3>CK>T2 in two types cultivar, and the breaking strength, lignin accumulation, monomer content under all treatments at grain filling stage and maturity stage were T1>T3>T2>CK in two types cultivar. The breaking strength of Shannong 16 and Jimai 22 under T1 treatment were increased by 24.69%, 19.97%, 13.15% and 26.92%, 15.36%, 5.87%, respectively, compared with CK, T2, T3 at grain filling stage. The average lignin accumulation of Shannong 16 and Jimai22 under T1 at each growth stage was 21.71%, 15.45% , 8.85% and 25.19%, 21.75%, 15.83% higher than CK, T2, and T3, respectively. The average content of S monomer was 18.82%, 18.48%, and 8.39% higher than CK, T2 and T3 at maturity stage, respectively. The expressive abundance of key genes involved in lignin biosynthesis pathway (phenylalanine ammonia-lyase: PAL, caffeic acid3-o-methytransferase: COMT, coumarate-3-hydroxyl oxidase: C3H, innamoyl Co A reductase: CCR, cinnamate 4-hydroxylase: C4H etc.) decreased with the growth process, that tended to T1>T3>T2>CK under different stage of growth. The 1000-grain weight of the nitrogen topdressing remaining one-time at booting stage was higher than other treatments. T1 treatment could increase the spike number, grain number and yield. The lignin accumulation and monomer content of stem in different internodes during the same stage were I1>I2>I3>I4>I5.【】Under the condition of high nitrogen application rate of 300 kg·hm-2and basal application rate of 1/3, the same amount of secondary nitrogen topdressing modes treatment at the rising stage and booting stages significantly improved the breaking strength, lignin accumulation, lignin monomer content, the expressive abundance of key genes involved in lignin biosynthesis pathway and yield after anthesis stage, compared with other spring nitrogen topdressing modes. Therefore, the same amount of secondary nitrogen topdressing mode at rising stage and booting stage could be used as an appropriate spring nitrogen topdressing mode under the condition of high nitrogen application rate of 300 kg·hm-2and basal application rate of 1/3 in Huang-Huai-Hai plain.
nitrogen topdressing modes; lodging resistance; lignin accumulation; grain yield; winter wheat

10.3864/j.issn.0578-1752.2020.21.009
2020-05-14;
2020-07-29
国家重点研发计划(2017YFD0301001,2016YFD0300403)
董荷荷,E-mail:dong15650092156@163.com。通信作者李勇,E-mail:wooowo@126.com
(责任编辑 杨鑫浩)
猜你喜欢
中国农学通报(2022年29期)2022-11-25
造纸信息(2022年8期)2022-11-10
江西农业大学学报(2022年3期)2022-07-07
大连工业大学学报(2022年1期)2022-03-14
中国生态农业学报(中英文)(2021年9期)2021-09-10
新能源进展(2020年1期)2020-03-09
中国造纸(2019年6期)2019-09-10
现代农业科技(2018年22期)2018-01-15
绿色科技(2017年7期)2017-05-12
江苏农业科学(2016年12期)2017-04-05
