
全缘叶绿绒蒿的花内热量来源和温度调节功能
2020-11-02 02:27钟涛段旭宇姜银银刘光立
广西植物 2020年9期
钟涛 段旭宇 姜银银 刘光立

摘 要:為探究全缘叶绿绒蒿( Meconopsis integrifolia )的花内热量来源和温度调节功能,该研究选择在全缘叶绿绒蒿的巴朗山居群,对其进行遮阴及去瓣处理,并采用红外热像仪监测全缘叶绿绒蒿的花内微环境温度日变化及花器官温度,用环境温度计监测环境温度。结果表明:(1)太阳照射显著提高全缘叶绿绒蒿花内微环境温度和花器官温度,全缘叶绿绒蒿的热量主要来源于太阳辐射。花内微环境昼夜温差显著低于环境昼夜温差,全缘叶绿绒蒿的花具有温度调节功能。(2)白天环境温度较高时,太阳照射显著提高全缘叶绿绒蒿花内微环境温度,花瓣会降低花内微环境温度;夜间环境温度较低时,花瓣闭合会提高花内微环境温度;花瓣闭合运动降低了花内微环境昼夜温差,产生了保温效果。(3)在太阳照射下,花器官温度差异显著,雌雄蕊温度显著高于花瓣温度,且花器官温度由雌蕊柱头中心点向外递减,全缘叶绿绒蒿能有效调控花器官各部位的温度。综上认为,全缘叶绿绒蒿的花内热量来源于太阳辐射,主要通过花瓣闭合运动降低花内微环境昼夜温差并能在太阳照射下调节各花器官的温度实现温度调节功能。
关键词:全缘叶绿绒蒿,热量来源,温度调节功能,花内微环境温度,花器官温度
中图分类号:Q948.112
文献标识码:A
文章编号:1000-3142(2020)09-1315-08
Abstract:In order to explore the floral heat source and temperature regulation function of Meconopsis integrifolia in the late July of 2017 and 2018, we conducted two treatments,the shaded treatment and the petal removal treatment in the Balang Mountain population. A infrared temperature imager was used to monitor the daily dynamics of within-flower air temperature and the temperature of different floral organs of M. integrifolia; the ambient temperature was monitored by an environmental thermometer. The results were as follows:(1) Solar radiation significantly increased the within-flower air temperature and the flower temperature of M. integrifolia,and the heat of M. integrifolia flowers resulted mainly from solar radiation. The within-flower air temperature showed significantly lower difference between day and night than the ambient temperature,so M. integrifolia has a kind of floral temperature regulation function. (2) When the ambient temperature was high during the daytime,the solar radiation significantly increased the within-flower air temperature of M. integrifolia,and the petals reduced the within-flower air temperature; When the nighttime ambient temperature was low,the petals closure increased the within-flower air temperature,and the closed motion of the petals effectively reduce the within-flower air temperature difference between the day and night and produced a certain thermal insulation effect. (3) In the sun,the temperature difference of the floral organs was significant,the pistil and stamen temperatures were significantly higher than the petal temperature,and the temperature of the floral organs decreased from the center point of the pistil stigma to the outside. M. integrifolia could effectively regulate the temperature of various floral organs. M. integrifolia has a thermal regulation function,mainly through the petal closure to reduce the within-flower air temperature difference between the day and night and regulating temperatures of various floral organs in the sun. However,the temperature regulation mechanism is still unclear,and the adaptative significance needs further study.
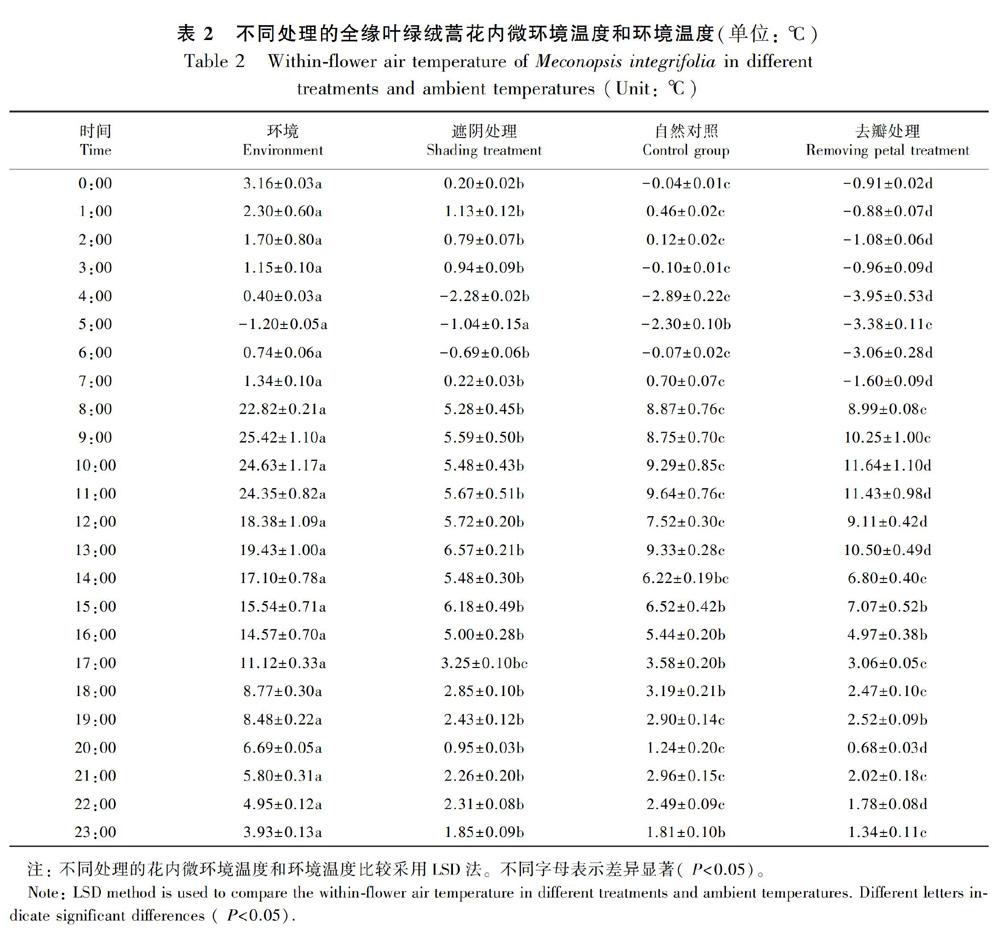
2.2 花瓣闭合运动和太阳照射对花内微环境温度日变化的影响
如图1所示,不同处理的全缘叶绿绒蒿花内微环境温度日变化趋势大致相同,去瓣处理的花内微环境温度日变化幅度最大,遮阴处理的花内微环境温度日变化幅度最小。白天环境温度较高时,去瓣处理的全缘叶绿绒蒿花内微环境温度高于自然对照组的花内微环境温度,遮阴处理的花内微环境温度最低。如表1所示,不同处理的全缘叶绿绒蒿花内微环境温度和环境温度相关性显著,去瓣处理的相关性高于自然对照组,遮阴处理的相关性相对最低。如表2所示,10:00—13:00,去瓣处理的花内微环境温度显著高于自然对照组,白天其他时刻差异性不显著,夜晚的所有时刻,去瓣处理的花内微环境温度均显著低于自然对照组。7:00—13:00,环境温度较高时,遮阴处理的花内微环境温度显著低于自然对照组,14:00—18:00遮阴处理的花内微环境温度仍然低于自然对照组,但差异性不显著。这表明,白天环境温度较高时,太阳照射会显著提高全缘叶绿绒蒿花内微环境温度,花瓣会降低花内微环境温度,夜间环境温度较低时,花瓣闭合会提高花内微环境温度,花瓣闭合运动有效降低了花内微环境昼夜温差,产生了保温效果。
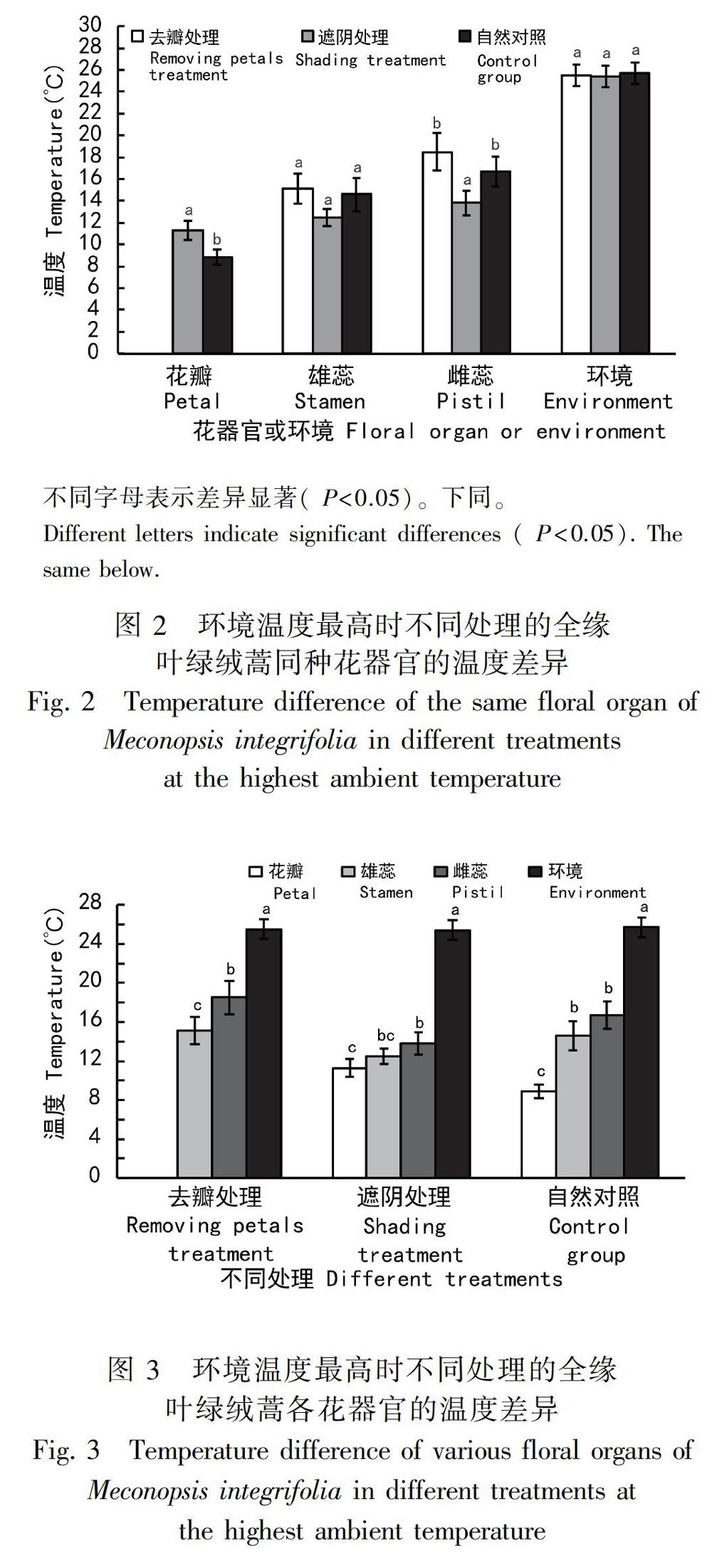
2.3 花瓣和太阳照射对花器官温度的影响
如图2所示,环境温度最高时,去瓣处理和自然对照的全缘叶绿绒蒿雌蕊温度显著高于遮阴处理的雌蕊温度,去瓣处理的雌蕊溫度最高,去瓣处理和自然对照的雌蕊温度差异性不显著。去瓣处理和自然对照的全缘叶绿绒蒿雄蕊温度高于遮阴处理的雄蕊温度,去瓣处理的全缘叶绿绒蒿雄蕊温度最高,不同处理的雄蕊温度差异不显著。遮阴处理的花瓣温度显著高于自然对照组。总的说来,遮阴处理的全缘叶绿绒蒿花器官温度低于自然对照的花器官温度。
2.4 不同花器官的温度差异
如图3和图4所示,环境温度最高时,不同处理的全缘叶绿绒蒿各花器官温度均有所差异,雌蕊温度最高,并且花器官温度由雌蕊柱头中心点向外递减。自然对照组全缘叶绿绒蒿雌雄蕊温度显著高于花瓣温度,且雌蕊温度最高;遮阴处理的全缘叶绿绒蒿雌蕊温度最高,花瓣温度显著低于雌蕊温度,但遮阴处理的雄蕊与雌蕊,雄蕊与花瓣的温度差异并不显著;去瓣处理的全缘叶绿绒蒿雌蕊温度显著高于雄蕊温度。这表明,在太阳照射下,全缘叶绿绒蒿能有效调节各花器官的温度。
3 讨论与结论
3.1 全缘叶绿绒蒿的花内热量来源
产热植物是开花期间能主动产热以调节花器官温度的植物,即使环境温度变化,产热植物的花器官温度能在相对较长的一段时间内保持比环境温度更高的恒定温度,其花器官温度和花器官呼吸不同字母表示差异显著( P<0.05)。下同。
耗氧量呈正相关(Nagy et al.,1972; Knutson,1974; Seymour & Schultze-Motel,1996,1998;Seymour et al.,1998)。王若涵(2010)定义开花生热效应为一些原始被子植物在开花时,花部各器官能始终保持在较高的温度范围内,且明显高于环境温度,直到传粉授精过程完成。本研究发现,全缘叶绿绒蒿的花内微环境温度变化和环境温度变化一致,花内微环境温度受环境温度影响显著且始终低于环境温度,说明全缘叶绿绒蒿不具有开花生热作用,不属于产热植物。植物热量在高海拔环境中主要来源于太阳辐射(Rejskova et al.,2010),花冠对部分太阳辐射能产生反射作用(张舒等,2008; Koski & Ashman,2015)。本研究发现,太阳照射显著提高了全缘叶绿绒蒿的花内微环境温度和花器官的温度,说明太阳辐射是全缘叶绿绒蒿花内热量的主要来源。吴云(2015)发现,7:00到9:00以及18:00到19:00时间段,全缘叶绿绒蒿的花内温度高于环境温度,且二者变化趋势呈负相关,而7:00到17:00内的其他时间段花内温度并不高于环境温度。这与本研究结果不完全一致,可能是由于测定方法不同。本研究发现,全缘叶绿绒蒿花内微环环境昼夜温差显著低于环境昼夜温差,说明全缘叶绿绒蒿的花具有温度调节功能。
3.2 花瓣闭合运动对花内微环境温度的影响
高山植物对温度的适应机制各不相同,相关报道集中于垫状植物、莲座叶丛植物、温室植物、棉毛植物和花具有向日性运动的植物(杨扬和孙航,2006)。垫状植物的球形或半球形结构使得其表面积大于体积从而提高了热量捕获效率,莲座叶丛植物盖尼千里光(Senecio keniodendron)和特尼克半边莲(Lobelia telekii)的莲座叶丛在夜晚相互折叠,其形状和卷心菜相似,使得卷叶内部的花芽免受夜晚低温伤害(Hedberg,1964)。Omori & Ohba(1996)的研究表明,温室植物塔黄苞片对其总状花序能起到类似温室的保温作用并对处于小孢子形成期的繁殖器官起到重要保护作用。艾沙江·阿不都沙拉木(2013)的研究表明,在伊犁郁金香花暂时性闭合的夜间里,花内微环境温度比周围环境温度高了3.18 ℃左右。与上述研究结果一致,花瓣闭合运动降低了花内微环境昼夜温差,产生了保温效果,使全缘叶绿绒蒿具有温度调节功能。全缘叶绿绒蒿花内微环境温度始终低于环境温度,可能是由于实验时间处于全缘叶绿绒蒿开花末期。本研究结果表明,白天全缘叶绿绒蒿花瓣张开,其花内微环境温度仍然比去瓣处理花内微环境温度低。有研究指出,辐照度的大小和棉毛植物花序内部温度的高低密切相关(杨扬和孙航,2006)。因此推测,可能是由于本研究以花丝基部温度作为花内微环境温度,花瓣的遮挡降低了部分花丝基部受到的辐照度。
3.3 花瓣对其它花器官温度的影响
棉毛植物的花序没有自升温系统,并推测水母雪兔子的头状花序的颜色和植株的形状能促进热量的吸收(Tsukaya,2002)。山罂粟和全叶仙女木的花瓣具有聚光作用,可以将太阳辐射聚集到繁殖器官上,且其花瓣决定了植物的向日运动,带动茎随着太阳入射方向的变化而弯曲转动,使繁殖器官获得最大热量(Kevan,1975)。本研究发现,太阳照射下,全缘叶绿绒蒿花瓣呈展开状态,去瓣处理后的全缘叶绿绒蒿雌雄蕊温度没有降低,反而略微升高,但并不显著。因此推测,太阳照射下,全缘叶绿绒蒿雌雄蕊温度显著偏高不是由花瓣导致,可能是由于雌雄蕊具有吸热作用。
3.4 不同花器官的溫度差异
本研究发现,自然对照组和去瓣处理的全缘叶绿绒蒿的雌蕊温度均最高,且花器官温度由雌蕊柱头中心点向外递减。毛茛科植物(Ranunculus adoneus)的花梗顶部是光的接收中心,且其向日性由花梗决定,受花瓣的影响很小(Sherry & Galen,1998)。高山植物的繁殖器官比营养器官积累更A-C. 环境温度最高时自然对照、遮阴处理和去瓣处理的全缘叶绿绒蒿热像图; D-F. 相应的可见光照片。
A-C. Thermal image of Meconopsis integrifolia in natural control,shading treatment and removing petals treatment at the highest ambient temperature; D-F. Corresponding visible light photo.
大的热量,有利于其在较短的生长期内完成繁殖(Dietrich & Krner,2014)。因此推测,全缘叶绿绒蒿可能存在向日性,且其向日性由花梗决定,使得雌蕊温度最高,从而有利于其生殖发育的顺利进行。本研究发现,太阳照射下,全缘叶绿绒蒿雌雄蕊温度显著高于花瓣温度,且花瓣温度最低,遮阴条件下,花器官温度差异不显著。塔黄苞片不含叶绿体,其对于紫外线的透射率显著低于红外线辐射(IR)光合作用激活光(PA),进一步明确了苞片的保温作用(Omori et al.,2000)。毡毛雪莲苞片具有显著的聚热增温作用,主要由于其特殊的解剖结构和对入射辐照的选择透过性(杨扬,2009)。因此推测,全缘叶绿绒蒿花瓣温度低可能与其解剖结构和光谱特性有关。
综上所述,环境温度显著影响全缘叶绿绒蒿花内微环境温度,太阳辐射显著影响花内微环境温度和花器官温度,太阳辐射是花内热量的主要来源。全缘叶绿绒蒿不属于产热植物,但其花内微环境昼夜温差显著低于环境昼夜温差,全缘叶绿绒蒿的花具有温度调节功能。全缘叶绿绒蒿主要通过花瓣闭合运动降低花内微环境昼夜温差并能在太阳照射下调节各花器官的温度实现温度调节功能,但其温度调节机制尚不明确,其适应意义有待进一步研究。
参考文献:
AYSAJAN A,2013. Reproductive biology of Tulipa iliensis and its adaptive strategies to the early spring environment [D]. Urumqi:Xinjiang Agricultural University:31-32. [艾沙江·阿不都沙木,2013. 伊犁郁金香的繁殖生物学特性及其生态适应对策 [D]. 乌鲁木齐:新疆农业大学:31-32.]
DIETRICH L,KRNERC,2014. Thermal imaging reveals massive heat accumulation in flowers across a broad spectrum of alpine taxa [J]. Alp Bot,124(1):27-35.
GRANT NM,MILLER R,WATLING JR,et al.,2010. Distribution of thermogenic activity in floral tissues of Nelumbo nucifera [J]. Funct Plant Biol,37(11):1085-1095.
HEDBERG O,1964. Features of Afroalpine plant ecology [J]. Acta Phytogeogr Suec,49:1-144.
KNUTSON RM,1974. Heat production and temperature regulation in eastern skunk cabbage [J]. Science,186(4165):746-747.
KEVAN PG,1975. Sun-tracking solar furnaces in high arctic flowers:significance for pollination and insects [J]. Science,189(4204):723-726.
KOSKI MH,ASHMAN TL,2015. Floral pigmentation patterns provide an example of Glogers rule in plants [J]. Nat Plants,1(1):14007.
NAGY KA,ODELL DK,SEYMOUR RS,1972. Temperature regulation by the inflorescence of Philodendron [J]. Science,178(4066):1195-1197.
OMORI Y,OHBA H,1996. Pollen development of Rheum nobile Hook. f. & Thomson(Polygonaceae),with reference to its sterility induced by bract removal [J]. Bot J Linn Soc,122(3):269-278.
OMORI Y,TAKAYAMA H,FLS HO,2000. Selective light transmittance of translucent bracts in the Himalayan giant glasshouse plant Rheum nobile Hook. f. & Thomson (Polygonaceae) [J]. Bot J Linn Soc,132(1):19-27.
REJSKOVA A,BROM J,POKORNY J,et al.,2010. Temperature distribution in light-coloured owers and inorescences of early spring temperate species measured by infrared camera [J]. Flora,205:282-289.
SEYMOUR RS,SCHULTZE-MOTEL P,1996. Thermo regulating lotus flowers [J]. Nature,383(6598):305-305.
SEYMOUR RS,SCHULTZE-MOTEL P,1998. Physiological temperature regulation by flowers of the sacred lotus [J]. Philos Trans R Soc B-Biol Sci,353(1371):935-943.
SEYMOUR RS,SCHULTZE-MOTEL P,LAMPRECHT I,1998. Heat production by sacred lotus flowers depends on ambient temperature,not light cycle [J]. J Exp Bot,49(324): 1213-1217.
SEYMOUR RS,SCHULTZE-MOTEL P,1997. Heat-producing flowers [J]. Endeavour,21(3):125-129.
SHERRYR A,GALEN C,1998. The mechanism of floral heliotropism in the snow buttercup [J]. Philosophy,21(10): 983-993(11).
TSUKAYA H,2002. Optical and anatomical characteristics of bracts from the Chinese “glasshouse” plant,Rheum alexandrae Batalin (Polygonaceae),in Yunnan,China [J]. J Plant Res,115(1117):59-63.
WU Y,LIU YR,PENG H,et al.,2015. Pollination ecology of alpine Meconopsis integrifolia in different altitudes [J]. Chin J Plant Ecol,39(1):1-13. [吳云,刘玉蓉,彭瀚,等,2015. 高山植物全缘叶绿绒蒿在不同海拔地区的传粉生态学研究 [J]. 植物生态学报,39(1):1-13.]
WANG RH,2010. A study on reproductive biology in Magnolia and its systematic characterization [D]. Beijing:Beijing Forestry University:76-77. [王若涵,2010. 木兰属生殖生物学研究及系统演化表征探析 [D]. 北京:北京林业大学:76-77.]
YANG Y,SUN H,2006. Advances in the functional ecology of alpine and arctic plants [J]. Acta Bot Yunnan,28(1):43-53. [杨扬,孙航,2006. 高山和极地植物功能生态学研究进展 [J]. 云南植物研究,28(1):43-53.]
YANG Y,2009. Study on ecological adaptation mechanism of special plant life types in alpine flowing rock beach in the Hengduan mountains [D]. Kunming:Kunming Institute of Botany,Chinese Academy of Sciences:1-2. [杨扬, 2009. 横断山区高山流石滩特殊植物生活型的生态适应机制的研究 [D]. 昆明:中国科学院昆明植物研究所:1-2.]
ZHANG GP,YANG ML,CHENG XX,et al.,2017. Effects of flower morphological characteristics on flower temperature accumulation in alpine plants [J]. Guihaia,37(7):822-828. [张国鹏,杨明柳,程贤训,等,2017. 高山植物花形态特征对花温度积累的影响 [J]. 广西植物,37(7):822-828.]
ZHANG S,YU WB,WANG H,2008. Flower heliotropism of alpine plants and its adaptive significance [J]. Plant Sci J,26:197-202. [张舒,郁文彬,王红,2008. 高山植物花的向日运动及其适应意义 [J]. 植物科学学报,26:197-202.]
(责任编辑 何永艳)
