
中国北方草原植物光谱beta多样性年内变化驱动因子分析
2020-10-30 05:35白岚田佳玲彭羽
生态环境学报 2020年8期
白岚,田佳玲,彭羽
中央民族大学生命与环境科学学院,北京 100081
植物多样性作为生态系统的重要组成部分,有助于维持草原所提供的生态系统多功能性,并对系统的生产力、稳定性及其抗干扰能力至关重要(Yann et al.,2017;Prieto et al.,2015)。近年来,将遥感技术应用于植物多样性监测,已成为生物多样性研究的最新趋势。目前,使用遥感数据提取物种丰富度或多样性信息总体上分为两种方法(Nagendra,2001):(1)直接法,从遥感数据中直接识别物种和物种分布,这对遥感数据的光谱和空间分辨率提出很高的要求;(2)间接法,由遥感数据衍生的环境参数或者生物学模型,结合一定的实地调研数据对植物多样性进行间接评估(Turner et al.,2003;Levin et al.,2007;Rocchini et al.,2007)。
光谱多样性(Ustin et al.,2010)是指通过光学遥感检测光谱反射率变化。植物物种之间的光谱差异反映出植物在化学、解剖和形态特征等方面的分类差异。因此,光谱多样性可以作为物种多样性的指示指标(Schweiger et al.,2018)。许多遥感指数被用于估算植物多样性(Wang et al.,2018)。例如,植被反射率指数的空间变异性(Carlson et al.,2007;Gould,2000;Wang et al.,2016;Wang et al.,2018),归一化植被指数NDVI的均值和标准差被用来研究物种丰富度或其他多样性指标(胡海德等,2012)。beta多样性是生物多样性不可或缺的重要组成部分,对于揭示植物多样性维持机制具有重要意义。beta多样性为理解物种与其环境之间的关系提供了关键环节(Christian et al.,2016),能够反映物种多样性变化的空间异质性。通过植被光学性质的空间变化评估beta多样性,已有在森林、草原等有较多应用。我们前期开发了164个高光谱指数用于估算浑善达克沙地草原植物物种beta多样性指数,结果验证了高光谱遥感对于估算植物物种beta多样性的潜力(彭羽等,2020)。本研究中我们利用遥感数据计算植物光谱beta多样性指数,从而估测研究区植物beta多样性的时空变化。
在草原生态系统中,植物多样性受地理地形、土壤、放牧以及气候变化等环境因素影响(赵峰侠等,2019)。其中,气候因素成为解释物种多样性变化的主要机制(张树斌等,2018;沈芳芳等,2019)。气候对植物多样性的影响会随着植被类型以及生态系统的不同而表现出空间和时间上的异质性(马明国等,2006)。研究发现,平均温度的变化会直接降低物种丰富度,而总降水量的变化直接破坏了群落稳定性(Boulangeat et al.,2014)。不同类型草原的物种丰富度、多样性的驱动因素存在差异。对吉林灌木群落物种多样性研究发现,物种丰富度与降水呈负相关关系,而与冬季低温没有显著相关性(张树斌等,2018)。在南非的克鲁格国家公园和美国堪萨斯州东北部的高草草原,年降水量的减少造成了物种丰富度的丧失(Hoover et al.,2014)。白永飞等(2000)对锡林河流域草原群落植物多样性沿水热梯度变化的研究发现,物种多样性与年降水量呈正相关,而与年平均温度呈负相关。孙小丽等(2015)发现,水热因子与内蒙古荒漠草原区物种多样性的相关性最高。穆少杰等(2012)发现在年际尺度上,内蒙古草原植被区受降水的影响;而荒漠植被区在年际、月际尺度上均与降水有较强的相关性。
现有的关于植物物种多样性与气候因子之间的关系研究,更多的集中在年际尺度,很少对草原植物beta多样性年内变化驱动因子的研究。不同月度的草原植物数量特征和多样性变化趋势与同时间段的温度和降水等关键气候因子的不同配置密切相关。为了进一步阐明中国北方不同草原植物多样性年内变化的主要驱动因子,本研究提取了1982—2017年卫星数据集的植物光谱beta多样性指数,结合月度和年度气候数据,对中国北方4种不同类型草原beta多样性年内变化特征与水热条件之间的相互关系进行了探讨,并确定影响草原植物多样性的关键气候因子。本研究为揭示中国北方草原植物多样性年内变化趋势及其与气候变化的关系、植物多样性维持机制,以及草原植物多样性的可持续利用提供参考。
1 研究区概况
沿着气候梯度,选取中国北方4个草原自然保护区:安西、西鄂尔多斯、锡林郭勒和图牧吉自然保护区,分别代表4种不同类型的草原生态系统:草原化荒漠、荒漠草原、典型草原和草甸草原。安西自然保护区(94°45′—97°00′E,39°52′—41°53′N)位于甘肃省瓜州县境内,是中国唯一以保护极旱荒漠生态系统及其生物多样性为主的多功能综合性自然保护区,地形特征南高北低,海拔1 300 m以上;保护区气候属温带大陆性干旱气候,降水极少,蒸发量大;植被类型主要为砾石荒漠植被、沙质荒漠植被等。西鄂尔多斯自然保护区(106°40′—107°44′E,39°13′—40°11′N)位于内蒙古自治区西部,以保护古老孑遗濒危植物及草原向荒漠过渡的植被带和多样的生态系统为主要对象,属典型的暖温带大陆性气候,有野生植物335种,其中国家级珍稀濒危保护植物7种。锡林郭勒草原自然保护区(115°32′—117°12′E,43°26′—44°33′N)位于内蒙古自治区锡林郭盟境内,主要保护对象为典型草原生态系统及各生态系统中繁衍生息的生物多样性,属中温带半干旱、干旱大陆性季风气候。图牧吉自然保护区(122°44′13″—123°10′24″E,46°04′12″—46°25′47″N)地处松嫩平原西侧,内蒙古自治区与黑龙江省、吉林省三省区的交汇地带,生物多样性十分丰富,属于温带大陆性季风气候,生境类型丰富,构成了草原、湿地、灌丛相间分布的多样的生态系统。
2 研究方法
2.1 植物多样性指数
在地理空间数据云中(http://www.gscloud.cn/),下载1982—2017年的Landsat系列产品的遥感影像数据。剔除云量超过10%的遥感影像,时间为每年的4、7、10月。遥感影像需要进行包括几何校正、辐射纠正和地形影响去除等一系列的预处理。结合当年土地利用类型,在保护区中进一步筛选草原化荒漠、荒漠草原、典型草原和草甸草原进行植物多样性分析。计算遥感影像1—7波段的变异系数之和,作为区域植物光谱beta多样性的指标(Wang et al.,2016;Gholizadeh et al.,2018):
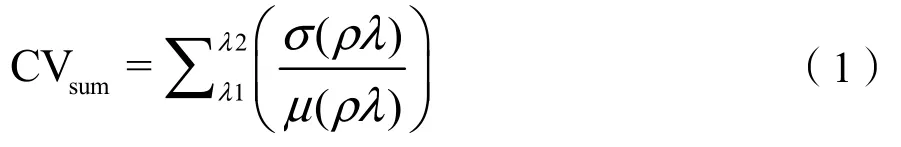
σ(ρλ)和μ(ρλ)表示波段λ的标准差和平均值。在ERDAS IMAGINE 2015和ArcGIS 10.4软件中进行计算。
2.2 气候因子来源及方法
本研究从中国气象数据服务中心获取全国2 400多个气象观测站点的月值温度和降水数据,时间跨度与Landsat遥感影像数据完全一致。基于研究区及其周边气象站点的气候数据,利用ArcGIS 10.4中克里金插值法处理得到各研究区气候数据。该方法在气象、气候、生态和环境领域得到广泛应用。最终将气候数据归类为7个气候变量:年高温(ATmax)、年低温(ATmin)、月平均温度(MMT)、月最低温度(MTmin)、月最高温度(MTmax)、年降水(AP)以及季降水等共计186种变量(详见表1)。

表1 本文所选气候因子及其含义表1 Alternative climate factors and their implications
2.3 数据分析
采用多元逐步线性回归(MSLR)方法分析186个气候变量和植物光谱beta多样性之间的关系,通过比较R2、P值和方程冗余度值筛选出关键气候因子。采用SSR方法比较年内4、7、10月beta多样性变化差异。采用1982—2017年间植物光谱beta多样性的变异系数来表示变化程度,变异系数CV表示植物beta多样性变异程度,可以用来衡量植被多样性变化的稳定性。公式如下:

续表1 本文所选气候因子及其含义Continued table 1 Alternative climate factors and their implications

S和分别表示1982—2017年beta多样性变化的标准差和平均值。
3 研究结果与分析
3.1 北方草原植物beta多样性变化规律

图1 北方草原植物光谱beta多样性变化Fig.1 Variation of spectral beta diversity of grassland plants in northern grasslands
由图1可知,各保护区草原植物多样性年内变化趋势存在较大差异。在安西,植物多样性在10月达到最高,其次是7月,最后是4月。10月与其他2个月份之间存在显著性差异(P<0.05)。在西鄂尔多斯,总体上7月植物多样性略低于4、10月植物多样性,3个月之间多样性差异不显著(P>0.05)。在锡林郭勒和图牧吉保护区内,总体上植物多样性年内变化趋势较为一致,均为4月植物多样性最高,其次是10月,而7月植物多样性最低。7月与其他2个月份之间存在显著性差异(P<0.05)。
3.2 草原化荒漠植物beta多样性变化驱动因子
多元逐步回归分析表明(表2),安西自然保护区4月草原beta多样性回归方程引入的气候因子是当年年低温(X4)、前1年7月月低温(X26)(AdjustedR2=0.030);7月植物beta多样性回归方程引入的气候变量是前1年10月月低温(X′32)(AdjustedR2=0.017);10月多样性回归方程引入的是当年8月月低温(X″45),当年6月月均温(X″22),前1年8月月低温(X″33),前1年12月月高温(X″58)(AdjustedR2=0.728)。
3个方程均引入了月度最低温因子作为重要的气候变量,且与植物多样性呈负相关关系,即月最低温越高,植物beta多样性越低。由此可以看出,安西不同季节的植物beta多样性主要受与低温有关的气候变量的影响。相比于10月,4、7月的植物多样性与气候因子回归方程中调整后R2都较低,说明除了气候变量,其他社会、环境因素对该地区产生一定的影响。但本研究我们只关注气候变量对植物beta多样性的影响,结果会有一定的偏差。我们将各月份的植物beta多样性与相对应的重要气候变量进行回归分析(图2)。当年年低温X4作为4月植物多样性的第一驱动力,在一定温度范围内年低温越低,植物beta多样性越高。前1年10月月低温X′32,温度保持在-7— -4范围内,7月的植物多样性较高。而超过或低于这一温度范围,植物多样性都会降低。而随着当年8月月低温X″45温度升高,10月植物多样性降低。
3.3 荒漠草原植物beta多样性变化驱动因子
在西鄂尔多斯,4月草原植物beta多样性回归方程(表3)引入的气候因子是前1年12月月高温(X46),前1年6月月低温(X25),前1年2月月高温(X36)以及前1年4月月低温(X23)(AdjustedR2=0.868);7月多样性回归方程引入的是前1年3月月高温(X′43),当年2月月高温(X′54),前1年1月月高温(X′41),前1年2月月高温(X′42)(AdjustedR2=0.986);10月多样性回归方程引入的气候变量为前1年1月月高温(X″47),前1年3月月高温(X″49)(AdjustedR2=0.931)。

图2 安西4、7和10月植物多样性与温度变量之间的关系Fig.2 Relationship between plant beta diversity and temperature variables in Anxi in April, July and October
相比于其他3个地区,西鄂尔多斯荒漠草原各月植物beta多样性与气候因子回归方程的调整后R2最高,均超过0.8,表明气候因素,尤其是温度能解释该地区各月植物beta多样性85%以上的变化。3个方程均引入了前1年月度最高温因子作为第一驱动力,其中4月beta多样性与前1年12月月高温呈正相关,而7、10月beta多样性分别与前1年3月月高温、前1年1月月高温呈负相关关系。特别的是,7、10月回归方程引入的气候变量均为月度最高温因子。由此看出,西鄂尔多斯荒漠草原不同季节的植物beta多样性主要受月度高温因子的影响。

表2 安西保护区草原植物beta多样性与气候因子回归方程Table 2 Regression equation of beta diversity of grassland plants and climate factors in Anxi Reserve

表3 西鄂尔多斯保护区草原植物beta多样性与气候因子回归方程Table 3 Regression equation of beta diversity of grassland plants and climate factors grassland in West Ordos reserve
3.4 典型草原植物beta多样性变化驱动因子
在锡林郭勒,4月草原beta植物多样性回归方程(表4)引入的气候因子是前1年8月月低温(X27)(AdjustedR2=0.010);7月引入的是当年年高温(X′2)(AdjustedR2=0.017);10月引入的是前1年8月月均温(X″12),当年6月月高温(X″64)以及前1年11月月高温(X″57)(AdjustedR2=0.018)。
根据回归模型,我们发现温度并不是该地区植物beta多样性变化的主控因素,可能与当地开展的生态保护和恢复工程使得草地覆盖得到明显改善有关(苏日古格,2016)。我们将各月份的植物beta多样性与相对应的关键气候变量进行回归分析(图3),发现4和7月份植物beta多样性方程分别引入了前1年8月月低温X27以及当年年高温X′2作为第一驱动力,且均呈负相关关系,温度升高会降低植物多样性。10月植物beta多样性与其第一驱动因子,前1年8月月均温X″12,并无相关性。10月多样性的影响因子比较复杂,受多种气候因子的综合影响。
3.5 草甸草原植物beta多样性变化驱动因子分析
在图牧吉保护区,4月草原植物beta多样性回归方程(表5)引入的气候因子是当年年高温(X2)以及前1年10月月低温(X29)(AdjustedR2=0.049);7月引入的是当年4月月高温(X′56)(AdjustedR2=0.083);10月引入的是当年5月月高温(X″63)(AdjustedR2=0.052)。

图3 锡林郭勒4、7和10月植物多样性与温度变量之间的关系Fig.3 Relationship between plant beta diversity and temperature variables in Xilingol in April, July and October
3个方程均引入了最高温因子作为第一驱动力。我们将各月份的植物beta多样性与相对应的重要气候变量进行回归分析(图4)。4月植物beta多样性受当年年高温X2的负面影响,温度越高,多样性越低。7月植物多样性与当年4月月高温X′56并无显著关系,温度变化较为稳定。而10月与当年5月月高温X″63呈负相关,温度升高,多样性降低。由此看出,图牧吉草甸草原不同季节的植物多样性主要受与高温相关的气候变量的影响。

表4 锡林郭勒保护区草原植物beta多样性与气候因子回归方程Table 4 Regression equation of beta diversity of grassland plants and climate factors grassland in Xilingol reserve

表5 图牧吉保护区草原植物beta多样性与气候因子回归方程Table 5 Regression equation of beta diversity of grassland plants and climate factors grassland in Tumuji reserve

图4 图牧吉4、7和10月植物多样性与温度变量之间的关系Fig.4 Relationship between plant beta diversity and temperature variables in Tumuji in April, July and October
3.6 植物光谱beta多样性空间变化特征
研究通过用1982—2017年植物光谱beta多样性变化的变异系数来衡量各研究区植被多样性空间变化的稳定性。变异系数越大,多样性的离散趋势越大,多样性的稳定性越差。反之,稳定性越好。图5显示4类草原植物光谱beta多样性在1982—2017年间发生了显著的变化。安西保护区的西部和西南部(图5A)、西鄂尔多斯保护区的西北部(图5B)、锡林郭勒保护区的西部和西北部(图5C)均发生了显著变化,而图牧吉保护区因为生态系统交错分布,beta多样性变化区域为北、中、南交错分布(图5D)。
4 讨论
不同类型草原植物多样性年内变化差异显著,说明群落类型不同,各研究区物种多样性变化也不相同。即使在同一草原,同一植被型类型,由于植物自身生物学特性以及不同生长阶段对水热等气候因子的响应不同,所以在月际尺度上的植物beta多样性变化也会有所差异。物候是影响植物物种多样性格局的主要时间因子。草原植物物候对气候变化的响应也会影响植物多样性。此外,不同类型草原植物多样性变化的气候驱动因子也存在差异。然而,相比于年际尺度,月际尺度上的气候因子对草原植物多样性影响更大。在引入的7个气候变量里,4个保护区均未引入降水变量(包括年降水和季降水);温度变量成为植物beta多样性年内变化的主要驱动因子。
西鄂尔多斯荒漠草原4、7和10月的植物beta多样性差异不显著,比较稳定,表明年内不同月份的群落具有较高的相似性,物种替换速率较少,即植物beta多样性较低。已有研究表明,在西鄂尔多斯荒漠草原,相邻群落间或者相似生境的不同群落间物种均表现出较高的相似性和较低的beta多样性(王乐等,2015)。在过去35年间西鄂尔多斯保护区所有月份的beta多样性呈增加趋势,最大值出现在4月,最小值出现在10月,表明研究区生长季春季提前、秋季推迟趋势较明显(额尔敦格日乐等,2016)。而安西草原化荒漠4月植物beta多样性较低,尚未有明显植物返青。对这两个地区4月植物beta多样性气候驱动力进行分析发现,安西4月beta多样性与年低温呈负相关,从1982—2017年年低温变化较稳定,呈缓慢的升高趋势(图6),因而植物beta多样性总体上也会有所降低。而在西鄂尔多斯,12月月高温是影响4月植物beta多样性的关键气候因子,两者之间呈现显著正相关。但35年来,12月月高温呈降低趋势(图7),进而植物beta多样性减少。

图5 4个草原保护区植物光谱beta多样性在1982—2017年间的变化Fig.5 Plant beta diversity changes in four different types of grasslands from 1982 to 2017

图6 安西保护区1982—2017年月最低温度和年低温变化Fig.6 Monthly minimum temperature (MTmin) change in Anxi reserve from 1982 to 2017
安西和西鄂尔多斯7月和10月植物beta多样性变化具有相同的趋势,即温度升高造成植物beta多样性降低。随着全球变暖,最高温和最低温的增加,适应较高温度的植物物种逐渐占优势,适应性趋向一致,物种分布均质化,beta多样性降低。在西鄂尔多斯,月度高温因子是影响7月和10月植物beta多样性变化的第一驱动力。由图7可知,从1982—2017年,西鄂尔多斯3月月高温和1月月高温均逐步升高。而温度的升高造成了7月和10月群落物种具有较高的相似性,植物beta多样性的降低。同样地,在安西,10月月低温和8月月低温分别与7月和10月植物beta多样性呈显著负相关,随着8月和10月最低温度不断升高,植物beta多样性逐渐减少。Marcilio-Silva et al.(2017)也强调了温度对巴西大西洋森林木本植物多样性的重要性,发现其多样性与最冷月份的最低温度呈负相关。夏季和秋季较高的温度会对植被生长起到限制作用(杭玉玲等,2014),尤其是夏季较高的温度可能会引起干旱加剧,草原植物可利用水减少。

图7 西鄂尔多斯保护区1982—2017年月最高温度变化Fig.7 Monthly maximum temperature (MTmax) change in West Ordos reserve from 1982 to 2017
锡林郭勒典型草原和图牧吉草甸草原植物beta多样性年内变化趋势基本一致:beta多样性以气候变化剧烈的4月较高,10月次之,7月最低(图1)。对这两种类型的草原来说,植被生长季春季提前和秋季推迟趋势十分明显。苗百岭等(2016)发现内蒙古不同类型草原物候变化趋势基本一致,即生长季长度均有不同程度的延长。典型草原和草甸草原植物返青期提前以及黄枯期推后趋势最大;而荒漠草原植物返青期提前以及黄枯期推后趋势较小。因此,图牧吉和锡林郭勒保护区植物beta多样性显著高于物候期变化较小的安西和鄂尔多斯保护区。
吴佳佳等(2020)研究发现,锡林浩特典型草原的一二年生植物功能群对气温、地表温度变化相对敏感,易受环境温度的干扰。植物种对温度变化的响应差异会进一步引起在月际尺度上原有群落的物种多样性改变。锡林郭勒与图牧吉7月植物beta多样性显著低于4、10月。7月水热条件充足,植物长势相似,草原物种组成相接近,造成beta多样性较低(图1)。有研究发现锡林郭勒典型草原7月及夏季的蒸散发(ET)达到最大值,是水分流失的主要途径。水分流失较多会进一步加剧草原干旱,对植被正常生长产生不利影响(张巧凤等,2017)。对7月植物beta多样性主要气候驱动因素分析,发现锡林郭勒7月主要受当年年高温(图8)的影响,年高温越高,植物beta多样性越低。这与其他自然保护区植物物种响应气候变暖而产生的物种组成趋同效应的趋势是一致的。而图牧吉7月植物beta多样性主要受4月月高温(图8)的影响,温度越高,植物beta多样性越高。但温度有可能超出植被生长的阈值,反而限制植被的生长,造成植物beta多样性降低。
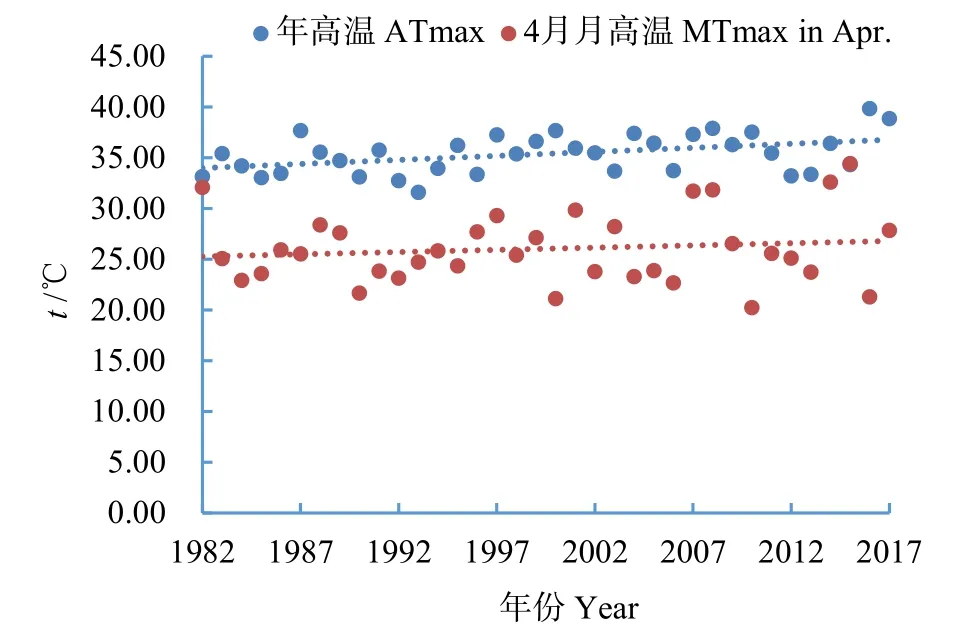
图8 1982—2017年锡林郭勒年高温变化以及图牧吉4月月高温变化Fig.8 Annual maximum temperature change in Xilingol and monthly maximum temperature in April change in Tumuji from 1982 to 2017
草原植物多样性不仅受气候因素的影响,还受地形、土壤、空间异质性等诸多因素的影响(沈芳芳等,2019)。本研究优先考虑了气候对草原植物多样性的影响,因为同一研究地点的土壤理化性质、土壤类型等一般较为稳定,不会有太大的变化。另外,植物多样性还受人为干扰。本文研究区均位于自然保护区内,所以未考虑放牧、耕地、人口等人为干扰问题。以往的研究大多是从气候年际尺度出发,探究植物多样性对气候变化的响应。但由于水热条件时空分配不均匀,使用气候月际尺度变量能更好地了解植被在不同生长条件下多样性的变化(Miao et al.,2015;丁勇等,2014)。研究各月份草原植物beta多样性变化趋势,对明确影响各月份植被多样性变化的主要气候驱动因子,以及对春季和秋季生长季提前或延长与否等具有重要意义。
5 结论
本研究利用1982—2017年遥感和气象数据,系统分析了中国北方4种不同类型草原植物beta多样性的年内变化趋势,并探讨了其对月际、年际温度和降水的响应特征。西鄂尔多斯荒漠草原4、7、10月植物beta多样性变化差异不显著,说明在月际尺度上不同月份的群落具有较高的相似性。安西草原化荒漠4月植物beta多样性较低,10月最高,表明尚未有明显的植物返青,且秋季生长季有推迟。锡林郭勒典型草原和图牧吉草甸草原4月和10月beta多样性较高,植被生长季春季提前和秋季推迟趋势十分明显。这种表现多与植物物候期表现相一致。中国北方草原年内植物beta多样性对环境温度变化十分敏感。温度对研究区植物beta多样性的影响因地区和季节而异。总体上,温度升高对植物beta多样性的增加产生了限制作用。
利用遥感数据可快速有效地估算植物物种beta多样性,但两者间关系模型的建立方法仍存在一些问题,例如,物候期-植物多样性对气候变化的协同效应、植物多样性响应气候变化的空间尺度依赖性、植物多样性alpha、beta及gamma成分之间的关系及其对气候变化的响应,这些都是未来待揭示的问题。
猜你喜欢
科学与财富(2021年36期)2021-05-10
中学生数理化·高一版(2021年2期)2021-03-19
中学生数理化·高一版(2021年2期)2021-03-19
作文小学中年级(2020年10期)2020-12-29
城市道桥与防洪(2019年5期)2019-06-26
中学生数理化(高中版.高二数学)(2019年6期)2019-06-24
江苏农业科学(2017年5期)2017-04-15
中国卫生(2016年10期)2016-11-13
科技与创新(2016年18期)2016-11-04
西夏学(2016年2期)2016-10-26
