
贡嘎山不同海拔峨眉冷杉叶片生态解剖结构特性研究
2020-10-27 06:12陈艳艳黄晓霞程小毛
西南林业大学学报 2020年6期
陈艳艳 黄 轩 黄晓霞,2 程小毛,2
( 1. 西南林业大学园林园艺学院,云南 昆明 650233;2. 西南林业大学国家林业与草原局
西南风景园林工程技术研究中心,云南 昆明 650233)
海拔作为自然稳定型变量,在多个领域的研究中都能起到非常重要的作用。环境因素受不同海拔梯度的影响而发生改变[1],不同的海拔环境下植物形态的特征以及生理会随着环境的改变而改变,由此为探究植被的生长对不同环境的响应提供了有利的条件。与植物光合作用和呼吸作用密切相关的叶片对环境因素的改变较为敏感,在植被的进化发展过程中,叶片结构特征对不同环境因素的响应较为灵敏。通过研究位于不同海拔梯度上的植物叶片的形态结构,分析其叶片的差异性,可以帮助进一步对此探明针叶林植物生长与环境因子之间的关系。针叶植物叶型与其他植物不同,具有独特的结构与形态,其叶片解剖结构特性的研究对于掌握针叶树种与环境因子间的关系有着重大的意义,因此对于探明针叶树种对长期环境变化的适应机制具有重要意义[2]。
峨眉冷杉(Abies fabri),又名泡杉、塔杉,是松科冷杉属植物。成熟峨眉冷杉一般高18~35 m,且粗壮,生长于垂直海拔的方向上,主要分布在海拔2 800~3 800 m,在不到海拔2 800 m 时,峨眉冷杉常与油麦吊杉(Picea brachytyla)形成混交林,在海拔2 800 m 以上则多为峨眉冷杉纯林[3]。峨眉冷杉是青藏高原东缘亚高山暗针叶山林区的优势树种,作为当地重要的水资源林,在调节气候类型、改善水质资源、保持水土环境以及维护生态系统平衡等方面均发挥着极为重要的作用[4]。其木材颜色为浅红褐色或黄褐带红,材质具有量轻质软,加工容易,平展开阔,可用于房屋、板料、家具及木纤维工业等原料,树胶可制成冷杉胶,用于做光学仪器粘合剂,树皮可用作增加尿醛树脂胶合性的增量剂[5]。现阶段关于峨眉冷杉的探究主要集中在群落的动态和结构、生境研究调查、生理机制及凋落物特征等方面,而关于海拔梯度对峨眉冷杉叶片解剖结构特征的影响的相关研究尚少,限制了其大规模应用。因此,通过对峨眉冷杉的叶片解剖结构进行分析,从叶片结构对贡嘎山峨眉冷杉在不同海拔下其叶片对生态的适应情况进行研究,从而增加对高海拔脆弱生态系统的认识,能够给保护高海拔脆弱生态系统提供一定的借鉴,且能够揭示高山植物与环境相互作用的复杂关系及给人类提供科学保护和持续利用高山植物资源的借鉴[6]。
1 研究区概况
贡嘎山地处青藏高原和四川盆地的交界处,地处东经101°03′50″~102°05′49″,北纬29°33′12″~29°36′30″,在青藏高原东侧,是横断山系下大雪山脉的主峰,是川西地区非常具有代表性的高山生态系统[7],其最高峰海拔7 566 m。由于受亚热带季风气候和高山气候的影响,海拔梯度上的气候差异明显。该地区为山地寒温带气候,气温4 ℃,年均降水量1 900 mm[8]。该区人为干扰因素较少,因生物多样性、地质结构、生态系统等较为复杂,所以较好的保留了原始森林的生态系统。因相对高差达6 km,所以完整的植被和土壤带谱在急剧变化的环境要素下形成[9]。在30 km的水平距离下相对高差约为6 400 m,林线和雪线位置分别是3 700 m 与4 900 m[10]。沿海拔的变化梯度依次是1 600~2 400 m 的常绿阔叶林带,最主要的是栎类(Quercusspp.)和包果柯(Lithocarpus cleistocarpus);2 400~2 900 m 的针阔叶混交林带,主要为麦吊云杉、云南铁杉(Tsuga dumosa)和扇叶槭(Acer flabellaatum);2 900~3 650 m 的暗针叶林带,树种以冷杉为主;3 700~4 100 m 的灌丛草甸带,高山杜鹃(Rhododendron lapponicum)为优势树种;4 100 m 至雪线为草甸带。针叶林是最重要的植被类型之一,范围最广分布在贡嘎山地区,东坡海拔2 600~3 200 m 是峨眉冷杉林带[11]。
2 材料与方法
2.1 采样方法
在四川省贡嘎山东坡海拔区域2 800~3 800 m处峨眉冷杉林进行取样,研究设置2 800、3 000、3 200、3 500、3 800 m 共 5个海拔梯度,每个海拔梯度取样5 棵,在每棵树各个方向采集当年生枝条中部叶片,每棵树采集叶片鲜质量为10 g。
2.2 石蜡切片及指标测定
制作切片和叶片解剖结构参数的测定切片制作方法参照李兰芳等[12]的方法。利用LEICA DM 2000 光学显微镜(尼康株式会社,日本)观测叶片角质层厚度、韧皮部厚度、上表皮厚度、维管束直径、木质部厚度、转输组织厚度、内皮层厚度、叶片长短径,每项指标均重复测量5次,取其平均值。
2.3 数据分析
采用SPSS22 统计分析软件对所有数据计算进行一元方差分析,分析的叶片特征参数分别位于不同的海拔,平均数之间的多重比较采用Ducan′s 的检验方法。
3 结果及分析
3.1 峨眉冷杉叶片位于不同海拔梯度下的解剖形态比较
由图1 可知,峨眉冷杉叶片结构主要包括表皮组织、海绵组织和栅栏组织,随海拔的升高,峨眉冷杉叶片表皮角质层的厚度呈现出先上升后下降随后小幅上升的趋势,并且在海拔3 000 m左右达到最大值,上表皮由2 层细胞组成且排列紧密。栅栏组织和海绵组织的厚度在海拔3 000 m左右时均达到最大值。海拔3 500 m 附近,叶片上表皮厚度达到最大值。由此可以看出,峨眉冷杉的叶片形态结构能够在不同海拔梯度的复杂环境条件下形成差异,需要对在不同海拔下的叶片解剖结构具体参数进行更深入的比较,分析其环境对于叶片结构的影响。
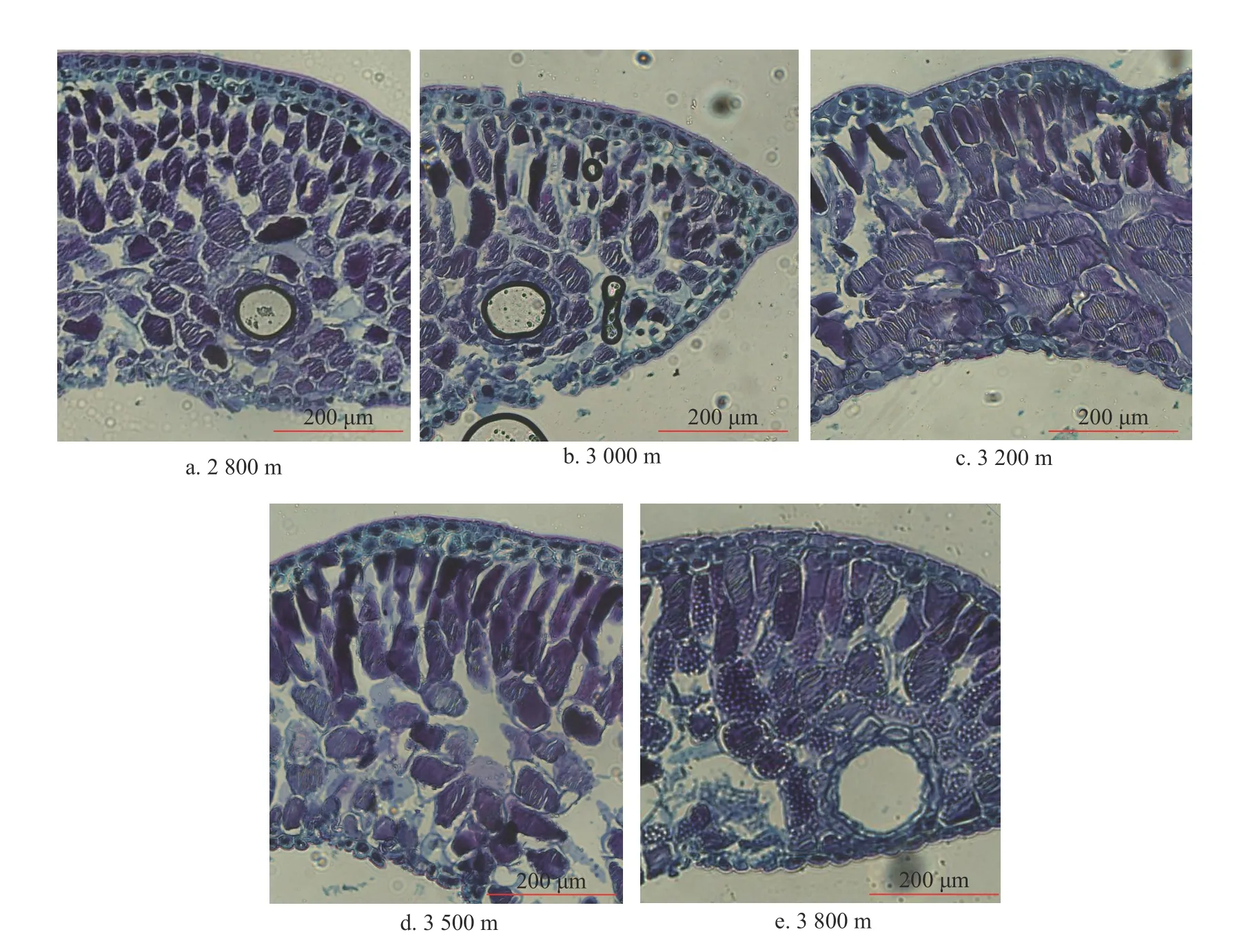
图1 峨眉冷杉叶片位于不同海拔下的解剖形态比较Fig. 1 A comparative study on anatomic form at gradient elevation of A. fabiri leaves
3.2 峨眉冷杉叶片解剖结构对海拔梯度的响应
由图2 可知,随着海拔的上升,峨眉冷杉叶片厚度呈波动性变化趋势。在海拔为3 000 m处,叶片总厚度达到最大值,与海拔2 800 m 处的最小值相比,上升了53.95%,差异显著(P<0.05),其余各海拔梯度间峨眉冷杉叶片厚度变化均不显著。不同海拔梯度下峨眉冷杉叶片的表皮组织结构参数随海拔的上升,叶片的表皮角质层厚度呈现出上升下降随后小幅上升的变化,且在海拔3 000 m 左右达到最大值,但其位于各个海拔梯度上的差异并不显著。在海拔3 500 m 附近,叶片上表皮厚度达到最大值,3 800 m 附近的最小值与之相比,降低32.49%,差异显著(P<0.05)。而随海拔的升高,峨眉冷杉叶片下表皮的厚度仅在海拔3 500 m 处有轻微回落,整体呈现上升趋势,并且在海拔3 800 m 左右达到最大值,与海拔2 800 m 左右的最小值相比,上升了75.31%,但该项指标位于各海拔梯度间差异不明显。
峨眉冷杉叶片叶肉组织结构参数位于不同海拔梯度下的不相同叶肉组织中的海绵组织与栅栏组织的厚度为(127.47±19.86)~(183.27±20.41)μm 与(91.81±10.60)~(127.27±14.64)μm,并且在海拔升高的情况下,海绵组织与栅栏组织都有相同的厚度变化趋势。当海拔3 000 m 左右时,海绵组织和栅栏组织的厚度均达到最大值。在各个海拔梯度下,栅栏组织厚度在海拔3 000 m与海拔3 200 m 处差异显著(P<0.05),相差38.62%,其余各海拔梯度间均无显著差异;但随着海拔的上升海绵组织厚度表现出先上升再下降的趋势,在海拔3 000 m 处达到最大值,并与其他海拔梯度间表现出显著差异(P<0.05)。
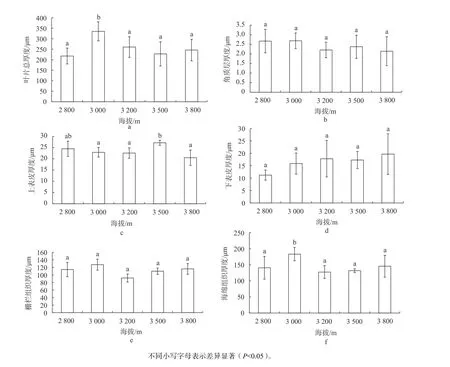
图2 不同海拔峨眉冷杉叶片解剖结构参数的变化Fig. 2 Changes in anatomical characteristics of A. fabri leaves at different elevations
3.3 不同海拔峨眉冷杉群体叶片各解剖结构间的相关性分析
由表1 可知,不同海拔峨眉冷杉群体叶片各项解剖结构间均存在一定的相关性。其中,在峨眉冷杉叶片总厚度与叶片表皮组织厚度之间,叶片总厚度与叶片上表皮间具有显著正相关关系(P<0.05),叶片总厚度与角质层厚度具有极显著正相关关系(P<0.01);当叶片总厚度与叶片叶肉组织厚度比较时,叶片总厚度与栅栏组织厚度及海绵组织厚度间均为极显著正相关关系(P<0.01)。在峨眉冷杉叶片表皮组织厚度之间均存在显著的相关性(P<0.05),其中叶片上表皮厚度与下表皮厚度为显著正相关(P<0.05),与角质层厚度之间存在极显著正相关关系(P<0.01);同时,叶片下表皮厚度与角质层厚度之间也存在显著的正相关关系(P<0.05)。在峨眉冷杉叶片叶肉组织厚度之间,叶片栅栏组织厚度与海绵组织厚度之间为极显著正相关的关系(P<0.01)。当峨眉冷杉叶片的表皮组织厚度与叶肉组织厚度比较时,叶片的下表皮厚度与海绵组织厚度之间呈出显著的正相关关系(P<0.01),角质层厚度与栅栏组织厚度和海绵组织厚度之间均存在极显著的正相关关系(P<0.01)。
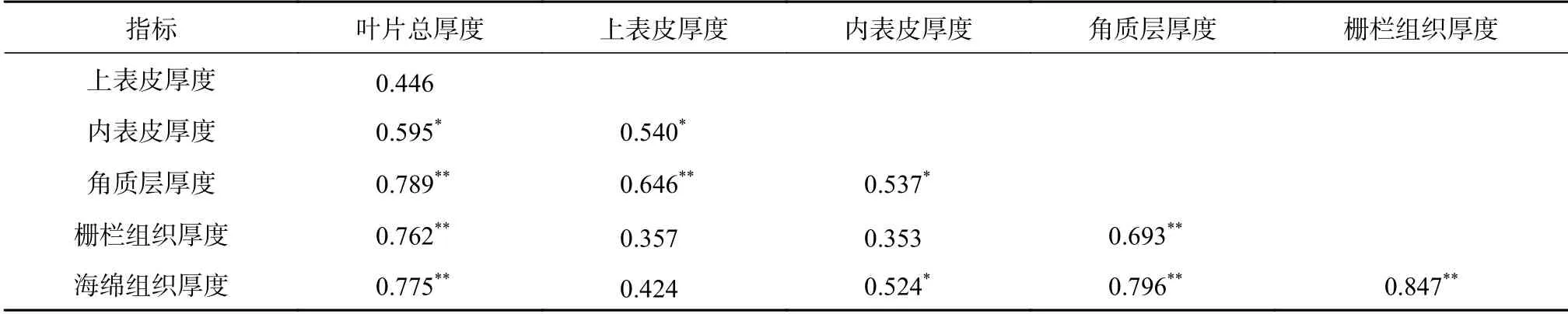
表1 峨眉冷杉叶片各解剖结构间相关性分析Table 1 The correlation analysis of anatomical characteristics of A. fabri leaves
4 结论与讨论
环境因素因受不同的海拔梯度影响而发生改变,在不同的海拔条件下,植物的生理及形态特征会随着环境的改变而改变,为了研究植物的生长对不同环境的响应提供了良好的条件[13]。与植物光合作用和呼吸作用密切相关的叶片对环境因素的改变较为敏感,在植物的进化过程中,叶片结构特征对于不同的环境因素响应较为灵敏,通过对位于不同海拔梯度上的植物叶片结构形态的研究,分析其在不同环境下的差异,可以帮助进一步探明植物生长和环境因子间的关系。海拔的变化往往会引起温度和水分条件的差异,随海拔的升高,植物叶片形态的可塑性也会发生变化。因此,在不同海拔生境中,降水和温度随着海拔的变化而成为影响植物叶片叶肉组织厚度的主要生态因子[12-16]。在本研究中,贡嘎山峨眉冷杉的植物叶片总厚度与叶肉结构中的海绵组织、栅栏组织厚度均在海拔3 000 m 附近达到最大值,造成这种状况的原因可能是由于贡嘎山东坡海拔3 000 m 附近为针阔叶混交林带,此处年降水量为1 938 mm,年均温4 ℃ 260 d,大气相对湿度大于90%[17-18],该处空气湿度大,气温低的良好的水分条件为叶片的生长以及厚度的增加提供了有利的条件,不仅具有保温的效果使得植物能够适应这种湿冷的环境,且海拔3 000 m处是阔叶针叶混交林带,植被种类相对丰富,凋落物结构及土壤养分也较其他海拔更为丰富,这样的环境条件均有利于植物的生长和发育。因此,在高海拔地区,叶片厚度增加不仅可以减少水分的蒸腾,防止高海拔地区风力对其造成的损伤,增加叶片的保水性,更有利于植株适应独特的高海拔地区严酷生态环境[13]。在本研究中,峨眉冷杉叶片上表细胞形状规则,排列紧密,而下表皮细胞排列较疏松,这样的结构特征可有效减少强辐射对叶肉细胞的损伤和细胞水分的散失[19-20]。植物进行光合作用的主要场所是叶肉细胞,栅栏组织以及海绵组织厚度对于植物光合速率的影响重大[21]。李广民等[22]发现短波辐射在海拔升高的情况下不断增强,这能够产生保护叶片栅栏组织细胞的稳定遗传结构,并由此防止叶面因温度升高而引起的组织灼伤。本研究中,贡嘎山峨眉冷杉叶片中的栅栏组织厚度与海绵组织厚度在海拔3 000 m附近达到最高值后回落,随后均随海拔的上升呈持续上升的趋势。这可能是由于随着海拔的不断升高,降水量增加与温度降低,光照增强,叶片栅栏组织和海绵组织厚度的增加可以有效减少水分的散失[13-14],并帮助植物体抵御干旱、寒冷胁迫及光照辐射带来的伤害,是峨眉冷杉在贡嘎山高海拔地区这类特定环境条件中形成的适应性机制[23]。
本研究中,贡嘎山峨眉冷杉的叶片在解剖结构之间基本都具有一定的关联性,说明随着海拔的升高,峨眉冷杉的叶片解剖机构均形成了针对不同海拔梯度的响应机制,且叶片组成部分间具有一定的联系。相关研究发现,叶片的表皮细胞以及表皮角质层具有防止植物叶片水分过度蒸腾、抵抗外界风雨及病虫侵害的作用[24]。而本研究中,尽管在不同海拔高度,峨眉冷杉群体间的叶片表皮各组织结构变化的趋势并不一致,但相关性分析结果显示,叶片上表皮、下表皮及角质层的厚度3 项解剖结构两两之间均存在显著或极显著的正相关关系,这说明叶片上表皮、下表皮及角质层的变化一致,这可能与高海拔区域,水热及光照条件的特殊性相关。
综合分析表明,由于贡嘎山海拔3 000 m 附近的降水量较大,植被种类相对丰富,因此峨眉冷杉植物叶片总厚度与叶肉结构中栅栏组织、海绵组织的厚度均在该海拔处达到最大值。叶片表皮间各组织结构的变化趋势不一致,但相关性分析结果显示,叶片上表皮、下表皮及角质层的厚度3 项解剖结构两两之间均存在着显著或极显著的正相关关系,这说明叶片上表皮、下表皮及角质层的变化是一致的。
猜你喜欢
意林·少年版(2021年7期)2021-06-15
林业与生态(2019年9期)2019-10-07
岷峨诗稿(2019年4期)2019-04-20
花火A(2019年1期)2019-03-22
科普童话·百科探秘(2018年7期)2018-09-13
当代教育(2018年4期)2018-01-23
现代养生·上半月(2017年7期)2017-07-07
发明与创新·大科技(2017年6期)2017-06-20
当代贵州(2016年36期)2017-05-08
第二课堂(小学版)(2014年4期)2014-08-02
