
TSWV 病毒粒子在系统寄主烟草K326 维管组织中的分布特征
2020-10-27 06:11温志多尚卫娜王田田吴少政苏晓霞张仲凯郑宽瑜
西南林业大学学报 2020年6期
温志多 丁 勇 尚卫娜 王田田 吴少政 苏晓霞 张 洁 张仲凯 郑宽瑜
( 1. 西南林业大学云南省高校林木生物技术重点实验室,云南 昆明 650233;2. 云南省农业科学院生物技术与种质资源研究所,云南昆明 650205;3. 浙江大学生命科学研究院,浙江 杭州 310058)
番茄斑萎病毒属(Orthotospovirus)病毒属于布尼亚病毒目(Bunyavirales)番茄斑萎病毒科(Tospoviridae),是该目中唯一可侵染植物的病毒属,属名由该属内发现的第1个病毒—番茄斑萎病毒(TSWV)命名而成的[1-2]。TSWV 寄主范围非常广泛,能侵染番茄(Solanum lycopersicum)、辣椒(Capsicum annuum)、烟草(Nicotiana tabacum)、莴苣(Lactuca sativa)、洋葱(Allium cepa)等多种作物,造成的经济损失非常严重[3]。到目前为止,该属已报道了至少30个番茄斑萎病毒属病毒种,不同种之间的寄主范围有很大的差异[4]。该属病毒为脂膜包被的球形病毒粒子,直径80~120 nm,表面有一层膜包裹。与布尼亚病毒目的其他病毒成员都很类似,番茄斑萎病毒属病毒基因组为三分体RNA,分别为L RNA、M RNA、S RNA 3 条RNA。番茄斑萎病毒代表种TSWV基因组总长约为16 600 Nt,其中L RNA 长8 897 Nt,M RNA 全长4 821 Nt,S RNA 全长2 316 Nt,核酸约占病毒粒子质量的1%~2%。L RNA 为负义链,仅编码1个开放阅读框,互补链编码大小为332 kDa 依赖于RNA 的RNA 复制酶(RdRp)。M RNA 为双义链,病毒链编码大小为33. 6 kDa移动蛋白NSm,互补链编码2个糖蛋白Gn 和Gc(G1 和G2),糖蛋白G1 的分子质量为78 kDa,G2的分子质量为58 kDa。S RNA 为双义链,病毒链编码长度52.4 kDa 的与致病性相关的蛋白NSs,互补链编码长度28.8 kDa 核壳体蛋白N,N 蛋白核苷酸序列是该属病毒分类的重要依据[5]。
植物病毒利用寄主的维管系统进行长距离运动是引起寄主植物系统性感病的关键环节。之前的研究表明大多数的植物病毒可通过筛管进行长距离运动,实现系统侵染[6]。但少数病毒也发现能利用导管进行长距离运动[7-9]。病毒在导管和筛管中的存在形式可分为3 种:病毒粒体、vRNP 复合体以及囊泡相关复制酶复合体(VRC)。南方菜豆花叶病毒(SBMV)和芜菁黄化病毒(TuYV)严格以病毒粒子的形式进行长距离运动[10-11],雀麦花叶病毒(BMV)和形影病毒属(Umbravirus)病毒是以RNP 复合体形式进行长距离运动[12-13],而芜菁花叶病毒(TuMV)在导管和筛管内存在形式则为囊泡相关VRC,由病毒膜蛋白6K2介导形成囊泡,包裹病毒RNA 及病毒RdRp 进行长距离运动[14-15]。
目前番茄斑萎病毒属在系统侵染寄主植物中的长距离运动机制仍不清楚。之前的研究利用互补实验证实TSWV N 及NSm 对运动缺陷型TMV长距离运动是必需的[16-17],并通过缺失突变发现NSm 分别由不同的结构域介导病毒细胞间运动及长距离运动,提示NSm 介导的病毒细胞间运动及长距离运动机制并不相同[18]。但是TSWV 长距离运动的输导组织和存在形式仍然未知。本研究利用透射电子显微镜观察TSWV 在系统寄主普通烟草K326 维管组织各类细胞中的分布特征,为解析TSWV 长距离运动机制提供参考。
1 材料与方法
1.1 材料来源
系统寄主植物K326 普通烟草由云南省烟草农业科学研究院提供,栽培于人工控制气候的室中,培养条件:温度25 ℃,湿度60%,光照时间16 h/d。TSWV 分离物14YV283 由云南省农业科学院生物技术与种质资源研究所作物病毒应用与创新团队实验室分离并保存于-80 ℃,经机械摩擦接毒于本氏烟上。
1.2 实验方法
1.2.1 TSWV 机械摩擦接毒及负染色检测
采取症状明显的叶片组织放置于已灭菌的研钵,并加入接种Buffer 充分研磨至匀浆;将金刚砂均匀撒在K326 烟的叶片表面,用棉签蘸取少量汁液在洒有金刚砂的叶片上轻轻摩擦2~3 次,5 min 之后用清水清洗叶面,并标记接种样品和时间。3个重复,每次接种5 株K326 烟苗,7~14 d后电镜负染色检测。病毒粒子的负染色检测参照方琦等[19]的方法,取少许K326 烟叶片置于载玻片上,滴加少许2.5%的戊二醛,用刀片尽量切碎(在切的过程中保持叶片湿润,及时滴加2.5%的戊二醛),将铜网有膜的一面倒扣在叶片浸出液表面3 min,然后用滤纸将铜网表面的液体吸干,将铜网有膜的一面扣于染色剂(钼酸铵或磷钨酸)液体表面2 min,充分染色,将铜网表面的液体用吸水纸吸干,置于红外灯下烘烤15 min,烘干后在透射电镜(FEI TECNAI G2,USA)下观察。
1.2.2 超薄切片制样及电镜观察
将经摩擦接毒TSWV 分离物14YV283 的K326 烟草以及健康对照K326 烟草样品取茎部组织切成1 mm× 3 mm 的小块,迅速放入具有2.5%戊二醛溶液的2 mL 离心管中固定;用0.1 mol/L 的PBS 漂洗3 次,15 min/次;经1%锇酸固定1.5~2 h,再用0.1 mol/L PBS 漂洗3 次,15 min/次;然后将样品分别进行30%、50%、70%、80%、90%、95%、100%乙醇脱水,每级脱水15 min,然后在100%乙醇再次脱水20 min 后用丙酮脱水20 min;脱水后的样品于纯包埋剂Epson-812 中过夜;先把纯包埋剂移至0.5 mL 离心管或包埋模具中,再放样品进去,然后放入聚合箱中聚合样品,70 ℃、12 h 或者60 ℃、24 h 两次;样品聚合完成后要对树脂包埋块做修正定位,修整其形状为梯形,每边小于0.5 mm,高度不易超过0.2 mm;修块完成后进行超薄切片,将包埋块中的生物样品切成尺寸为50~70 nm 薄片;切片完成后,用睫毛笔将切片拨开,用镊子夹住铜网,对准液面上的切片轻轻一沾,使切片覆盖于铜网上,去除铜网表面多余水分置于铜网盒;然后先用醋酸双氧铀染色5~10 min 后蒸馏水冲洗,再用柠檬酸铅染色5 min 后冲洗,烘烤灯烘烤后上透射电镜观察。每个样品做3个包埋块,每个包埋块切片后,在适当放大倍数下观察20个有效视野,最后进行统计分析。
2 结果与分析
2.1 TSWV 机械摩擦接毒及负染色观察
摩擦接毒TSWV 7 d 时,接种叶片上出现黄色斑点,后期逐步变为坏死斑,直至接种叶片整片坏死;8~9 d 时,顶部的非接种叶片上开始显示出坏死斑点,到第14 天,症状扩散到大部分的非接种叶片,顶芽坏死(图1b)。实验于第10 天取接毒后的顶部非接种叶片进行负染色电镜观察,发现有囊泡包裹的80~120 nm 直径病毒粒子聚集体(图1c)。表明TSWV 已系统侵染至整个K326 烟草植株。

图1 TSWV 症状及负染色观察结果Fig. 1 Symptoms and negative staining of TSWV
2.2 导管细胞病理切片观察结果
成熟导管是一种死亡细胞,只存在细胞壁。细胞壁分为初生细胞壁及加厚的次生细胞壁。而且上下两个细胞是贯通的,位于维管束的木质部。其功能是把从根部吸收的水和无机盐输送到植株身体各处。在透射电子显微镜下观察接毒植株样品超薄切片,并与健康K326 植株导管相比较,可观察到在系统侵染TSWV 的K326 烟草导管内有囊泡包裹的病毒粒体,大小约80 nm、球状,并发现有病毒核壳体N 聚集体的类似物(图2a,b)。未摩擦接病毒的健康对照K326 植株,其导管中基本未发现内容物,其中也没有发现病毒粒子及病毒核壳体聚集体(图2c,d)。正常情况下,导管主要运输水和无机物,但在TSWV 侵染条件下发现TSWV 病毒粒子存在于导管中,说明TSWV 病毒粒子有可能利用导管进行长距离运动,实现系统侵染。
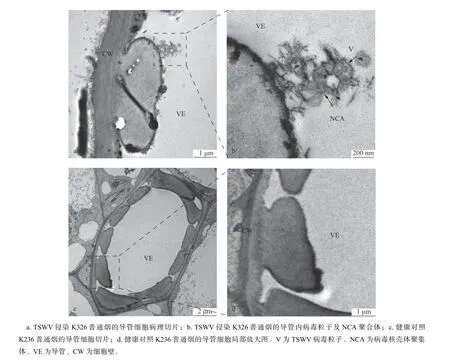
图2 导管细胞病理切片Fig. 2 Characteristics of vessel elements by TEM
2.3 筛管细胞病理切片观察结果
成熟筛管显著特征是没有细胞核或液泡,具有筛管特异性P 质体,2个筛管之间具有筛板。胞间连丝在结构和功能上分别特化为筛板上的筛孔和伴细胞界面上有较大开口的孔-胞间连丝单元(PPU)。PPU 大小介于胞间连丝和筛孔之间。它与胞间连丝相似,具有中央膜质通道,与筛孔通道具有相同的胼胝质沉积模式和潜在的病毒粒子输运能力。经超薄切片后,经透射电子显微镜下观察,与健康K326 植株相比较,在TSWV侵染的K326 烟草筛管中,并未发现该病毒粒子,但是可观察到囊泡状小体,直径80~200 nm(图3a,b),而在健康对照的K326 植株中具有较少囊泡状小体(图3c,d)。在统计的60个有效视野中,TSWV 侵染的样品中有筛管细胞53个,筛管内直径80~200 nm 的囊泡共计432个,平均1个筛管细胞内囊泡数量为8.15个;对照样品中有筛管细胞45个,筛管内直径80~200 nm 的囊泡数量189个,平均1个筛管细胞内囊泡数量为4.2个。推测TSWV 侵染导致筛管内囊泡增多,其囊泡可能为TSWV 囊泡相关VRC。
2.4 伴细胞病理切片观察结果
伴细胞是一种特殊类型的薄壁组织细胞,它在起源、位置及功能上和筛管分子紧密联系。它和筛管分子由同一个母细胞分裂而来,提供筛管必需的营养物质。健康K326 烟草的伴细胞有较大的细胞核,有丰富的细胞器和膜系统,高尔基体、线粒体、粗面内质网较多,细胞质密度也较大,这些都表明伴细胞有很高的代谢活性(图4e,f)。在透射电子显微镜下观察TSWV 侵染的K326 烟草伴细胞,发现细胞质中有成熟的病毒粒子,而且细胞质中还有大量囊泡状小体,直径在120 nm 左右(图4a,b)。同时线粒体的基质膨大,有的甚至整个线粒体崩解(图4c,d)。在统计的60个有效视野内,TSWV 侵染烟草样品中共有伴细胞43个,其中35个伴细胞具有上述病理现象。说明TSWV 的侵染影响了伴细胞线粒体的合成代谢。而大量囊泡小体有可能为TSWV囊泡相关VRC,介导TSWV 由伴细胞通过PPU向筛管运输。
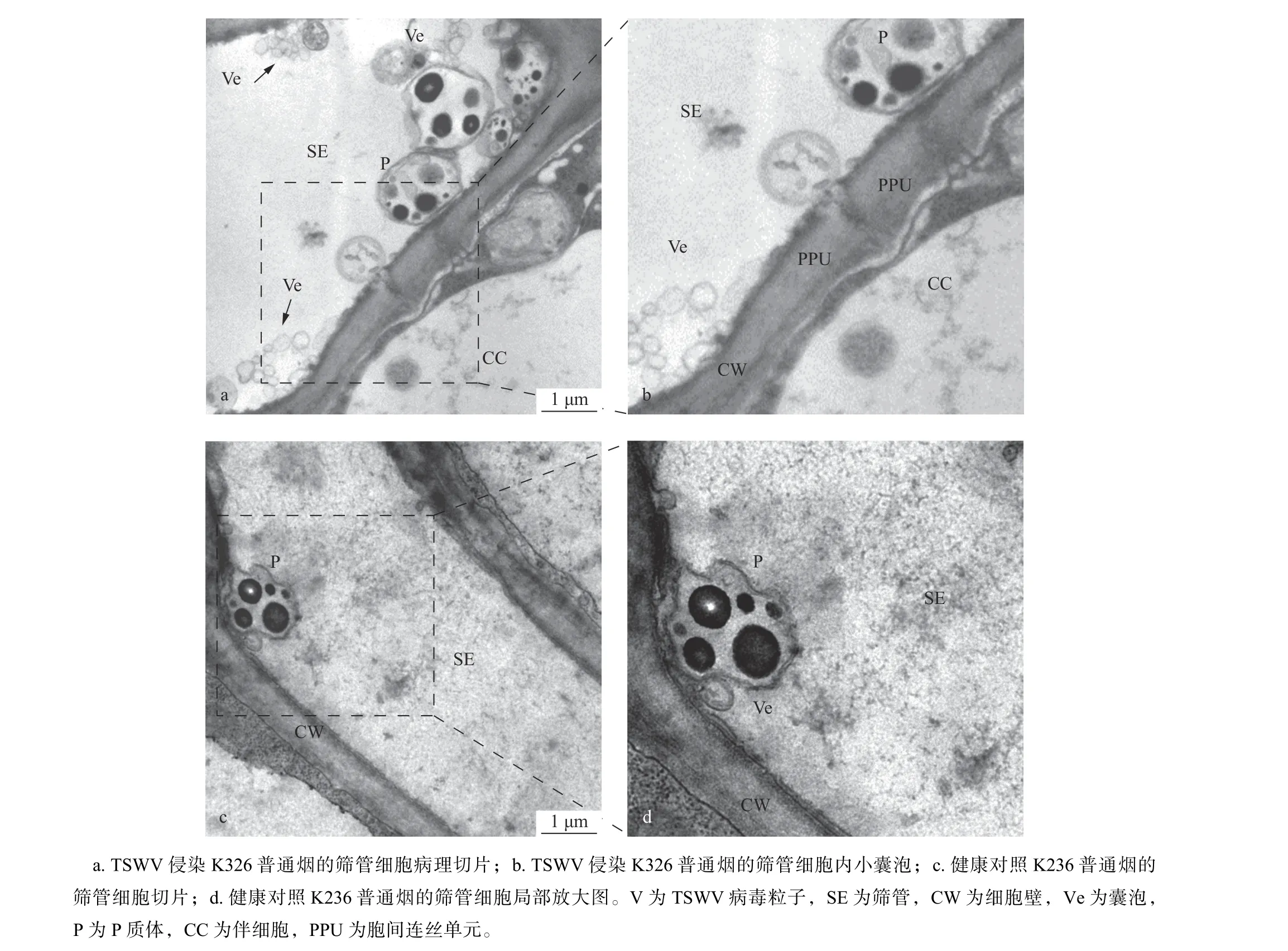
图3 筛管细胞病理切片Fig. 3 Characteristics of sieve elements by TEM
2.5 维管薄壁细胞病理切片分析
维管薄壁细胞的特征是具细胞核、细胞质、液泡、线粒体、叶绿体、高尔基体及内质网等多种细胞器。感染TSWV 的K326 普通烟草经超薄切片在透射电镜观察,发现在细胞质中存在TSWV病毒粒子聚集体,病毒粒子直径约80 nm,病毒粒子聚集体外有单层囊膜包裹,囊膜上具有黑色电子致密点状物质,推测为核糖体(图5a,b)。未接毒的健康对照烟草维管薄壁细胞,未发现有TSWV 病毒粒子,细胞质中可观察到叶绿体、线粒体、高尔基体等细胞器(图5c,d)。维管薄壁细胞虽然不具备长距离运输病毒的潜在功能,但维管薄壁细胞和叶肉细胞相似,具有较完善的、多种类型的细胞器,因此可能是病毒长距离运动过程中提供病毒复制增殖及装配的重要场所。
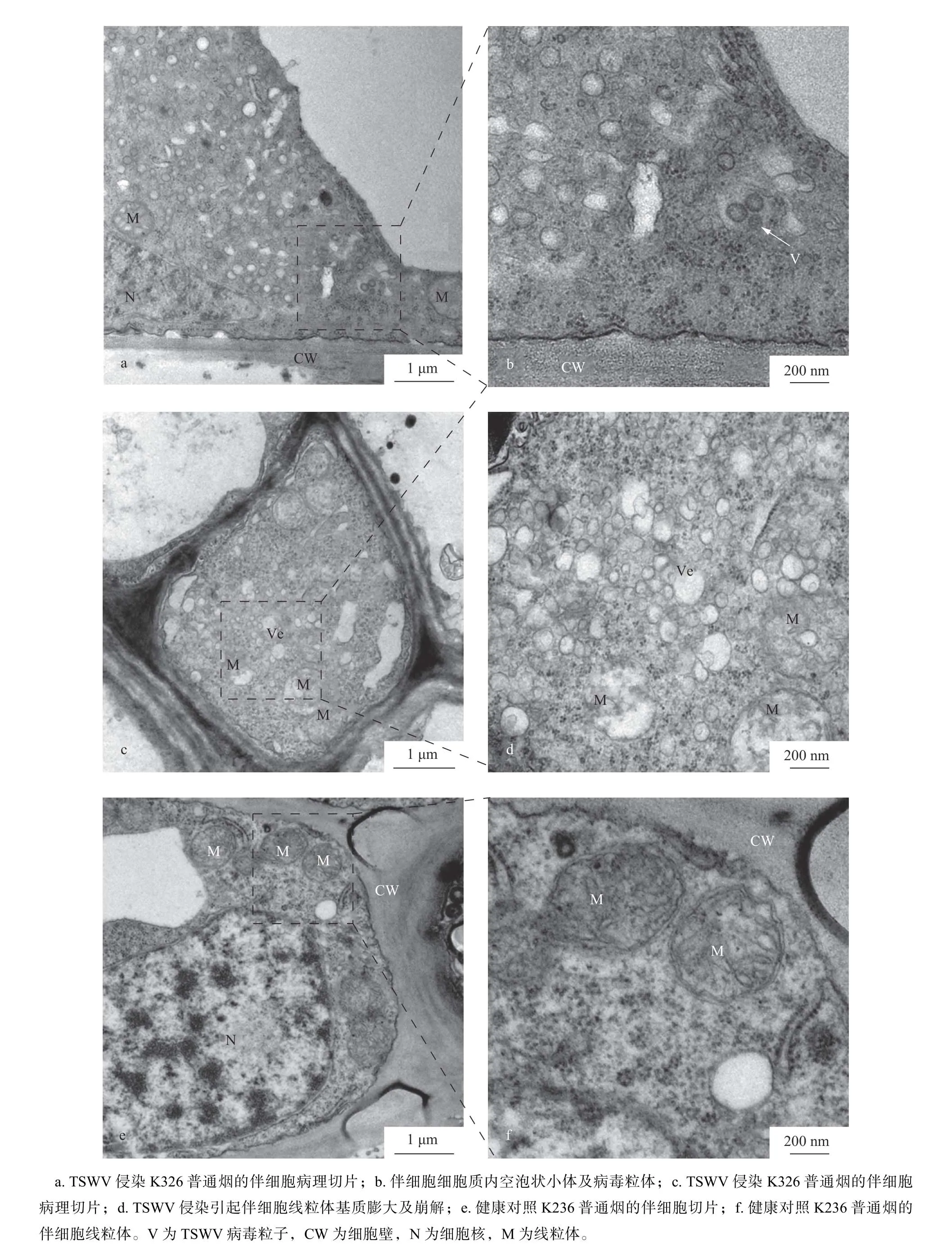
图4 伴细胞病理切片Fig. 4 Characteristics of companion cells by TEM
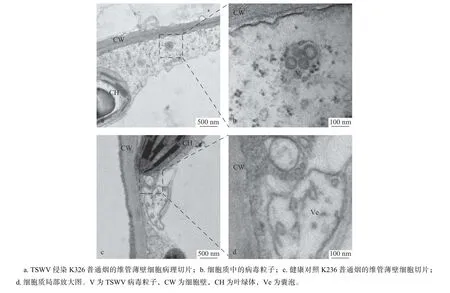
图5 维管薄壁细胞病理切片Fig. 5 Characteristics of vascular parenchyma cells by TEM
3 结论与讨论
植物的维管组织是植物重要的运输系统及支持系统。维管束是维管组织在植物器官中的束状结构,分为木质部和韧皮部。木质部的导管其功能主要运输水和无机物,韧皮部的筛管其功能主要是运输有机物质,即将叶片中形成的光合产物运输到植物体的各部分。近年的研究还发现筛管还能运输抗逆相关信号分子、小RNA 等物质[20]。植物病毒可利用植物维管组织这一高效的运输系统,在植物中进行长距离运动以实现系统侵染。在植物病毒系统侵染的模型中,植物病毒由薄壁细胞进入导管/筛管,随汁液流进行长距离运动,最终从导管/筛管卸载到邻近的薄壁细胞,建立新的侵染点[6]。植物病毒系统侵染研究中的难点在于解释病毒如何进入导管/筛管,又如何从导管/筛管卸载到近邻薄壁细胞。目前普遍性的推测是:病毒由韧皮部伴胞进入筛管的途径为PPU;病毒由木质部薄壁细胞进入导管的途径为纹孔膜[14-15]。本实验研究发现TSWV 病毒粒子存在于导管,说明TSWV 有可能利用导管进行长距离运动。TSWV 为大颗粒病毒,病毒粒子直径达80~120 nm,远大于植物胞间连丝的直径(烟草胞间连丝直径20~50 nm),那么TSWV 病毒粒子由叶肉细胞进入导管的机制仍然未知。之前的研究发现有一部分病毒,如PVY 属病毒TuMV以囊泡相关VRC 形式存在于导管和筛管内,由病毒膜蛋白6K2介导形成囊泡,包裹病毒dsRNA 及病毒RdRp、CP 进行长距离运动[14-15]。6K2为膜蛋白,能诱导ER 膜结构发生改变,产生6K2相关囊泡,介导TuMV 的胞间运动[12]。番茄斑萎病毒属病毒编码的运动蛋白NSm 与6K2功能类似,也为膜蛋白,已证实能够诱导ER 膜结构改变,产生病毒相关囊泡[21-22]。本实验研究发现伴细胞上有大量病理性的小囊泡,并且在与伴细胞相邻的筛管中也发现有小囊泡增多的现象,这些病理性小囊泡是否为TSWV 囊泡相关VRC,TSWV 是否以VRC 的形式通过筛管进行长距离运动还需要进一步的实验验证。
本研究应用TEM 技术观察了TSWV 在系统寄主普通烟草K326 维管组织中各类细胞中的分布特征,发现导管内有囊泡包裹的病毒粒体及病毒NCA 类似物,维管薄壁细胞内质网池中分布有病毒粒体、且囊泡在细胞质中有增多现象,伴细胞中存在成熟的病毒粒体和大量的直径120 nm 左右的囊泡小体、且伴随有线粒体基质膨大和线粒体崩解等现象。而在筛管中未发现有病毒粒体,但存在较多的直径80~200 nm 的囊泡小体。暗示TSWV 病毒粒体可能通过导管进行长距离运动,TSWV 囊泡相关复合体(VRC)可能通过筛管进行长距离运动。
猜你喜欢
当代水产(2022年1期)2022-04-26
汽车实用技术(2022年6期)2022-04-02
建材发展导向(2021年13期)2021-07-28
铁道建筑技术(2021年4期)2021-07-21
汽车工程(2021年12期)2021-03-08
中国瓜菜(2019年8期)2019-09-19
汽车零部件(2017年3期)2017-07-12
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
现代企业(2015年6期)2015-02-28
