
光谱对灰茶尺蠖成虫光反应行为的影响
2020-10-16 09:34乔利洪枫金银利耿书宝郭世保
茶叶科学 2020年5期
乔利,洪枫,金银利,耿书宝,郭世保*
光谱对灰茶尺蠖成虫光反应行为的影响
乔利1,2,洪枫1,2,金银利1,2,耿书宝1,2,郭世保1,2*
1. 信阳农林学院,河南 信阳 464000;2. 河南省豫南农作物有害生物绿色防控院士工作站,河南 信阳 464000
为获得灰茶尺蠖(Warren)的趋光/避光反应特征、光谱反应范围及性别差异,采用光行为学的方法研究了370~600 nm范围内的18种不同波长的单色光对成虫趋光、避光反应行为的影响。结果显示,各单色光均能引起灰茶尺蠖成虫一定的趋光或避光反应,雌虫在橙光(600~605 nm)、绿光(520~525 nm)、紫光(370~375 nm)范围内趋光性明显;雄虫在绿光(520~525 nm)、紫光(395~400 nm和390~395 nm)范围内趋光性明显;雌虫在冰蓝青光(490~495 nm)、蓝光(440~450 nm)和红光(660~665 nm)范围内避光性明显;雄虫在紫光(400~405 nm)、蓝光(440~450 nm)、冰蓝青光(490~495 nm)和自然白光(CK)范围内避光率较高。研究表明,灰茶尺蠖成虫对不同波长光的趋、避性存在差异,性别对其光谱行为反应有一定的影响。
灰茶尺蠖;成虫;光谱;趋光反应;避光反应
灰茶尺蠖(Warren)属鳞翅目(Lepidoptera)尺蛾科(Geometridae),是茶树上的主要食叶类害虫,在我国各产茶省份都有发生,以幼虫取食嫩叶危害,大发生时可将茶树取食成秃枝,对茶叶产量、品质造成严重影响[1-3]。目前除少量有机茶园外,对灰茶尺蠖的防治大多采用化学农药,导致灰茶尺蠖抗药性增加,农药残留问题也严重影响茶叶的质量和产量[4-5]。灰茶尺蠖危害给茶农造成巨大的经济损失,已成为制约茶叶优质、高产的重要因素之一,因此,研究茶园害虫防治的新方法对提升茶叶产量和品质具有重要的现实意义。
利用昆虫的趋/避光性对成虫进行灯光诱杀、驱赶或干扰其发育节律等措施,减少其在田间的落卵量以避免其大量发生的方法为害虫防治提供了新思路。已有研究表明,多数昆虫对短波光具有较强的趋性[6],但也有部分昆虫对长波光的趋性较强[7]。光照强度、昆虫的生理状态等因素对昆虫的趋光行为也有显著地影响。范凡等[8]研究表明,在340~605 nm波谱范围内,西花蓟马(Pergande)对380、440 nm和498~524 nm波长的光表现出趋光行为,且趋光率随光强增强而增大。在340~605 nm波谱范围内,二点委夜蛾(Moschler)成虫的趋光率较高,均在56%以上,且对360 nm紫外光趋光性最强[9]。对茶尺蠖趋色性的研究指出,其有趋蓝色的习性,可用诱虫板进行诱杀[10];当采用单波段诱杀时,茶尺蠖的趋光波段在365~410 nm[11];利用紫光和蓝光组成的LED双光谱太阳能杀虫灯诱杀时发现,380~400 nm的波段对茶尺蠖诱杀量最多[12];也有研究表明光谱范围295~490 nm、300~390 nm、400~505 nm对茶尺蠖的诱杀效果最好[13];关于灰茶尺蠖成虫更偏好哪种光源以及它们对不同单色波长的趋/避光行为目前尚无研究报道。
本试验采用行为测试的方法研究了灰茶尺蠖对18种不同波长单色光的行为反应及性别差异,为其趋光性和光视觉的深入研究提供理论依据,也为采用灯光物理防治措施对灰茶尺蠖综合治理提供科学依据。
1 材料与方法
1.1 虫源及饲养方法
灰茶尺蠖虫源来自于河南省信阳市浉河区浉河港镇白庙村茶园,于人工气候室内鲜叶继代饲养,试验用虫为饲养5代后的成虫,饲养环境为温度22~26℃、相对湿度60%~70%,光周期为12 L∶12 D。
1.2 试验设备
RTOP-310Y型人工气侯箱,浙江托普云农科技股份有限公司;PM6612数字照度计,深圳市华谊智测科技股份有限公司;养虫笼(50 cm×50 cm×60 cm),用于灰茶尺蠖羽化和产卵;自制光反应装置(图1)。
1.3 供试光源
供试光源根据光谱波长进行命名。光源颜色及波长分别为橙光(600~605 nm)、黄光(590~595 nm)、绿光(520~525 nm)、冰蓝青光(490~495 nm)、蓝光(460~465 nm)、紫光(370~375 nm)、紫光(410~415 nm)、宝蓝光(440~450 nm)、紫光(400~405 nm)、紫光(380~385 nm)、自然白光(CK)、红光(620~625 nm)、植物红光(660~665 nm)、琥珀光(595~600 nm)、紫光(390~395 nm)、紫光(385~390 nm)、紫光(395~400 nm)、紫光(420~425 nm)。
1.4 试验装置
趋光反应观察装置所用材料为纸板,包括可形成暗室的箱体,安装在箱体左侧面的光源(LED灯),自左至右安装在该箱体中的前隔板、挡光板和后隔板,前隔板和后隔板将箱体分隔成昆虫趋光室(30 cm×30 cm×20 cm)、昆虫栖息室(35 cm×30 cm×20 cm)和昆虫避光室(30 cm×30 cm×20 cm),前隔板上设置有趋光通孔,趋光通孔上安装可遮蔽的趋光通孔盖板;后隔板上设置避光通孔,避光通孔上安装可遮蔽的避光通孔盖板;挡光板固定在昆虫栖息室。
1.5 试验方法
试验于暗室中进行。为获得个体差异较小且接近自然条件的试验虫源,分别选取灰茶尺蠖2日龄雌、雄成虫置于趋光行为反应装置的栖息活动室内,每次单色光刺激前,将雌、雄成虫置于暗室暗适应1 h。每种单色光刺激时间为15 min,重复10次,每次重复间隔5 min,每次重复20头。为减少试验误差,每种单色光采用多组试虫,试虫不重复利用。经18种不同波长单色光刺激后,分别统计趋光室和避光室的试虫数,计算趋光率和避光率。
趋光率=趋光反应室虫数÷试虫数×100%;
避光率=避光反应室虫数÷试虫数×100%。
1.6 数据处理
试验数据采用Excel2010软件和SPSS 16.0(SPSS Inc.,Chicago,IL)进行处理。不同光源处理间差异显著性采用测验,多重分析采用Tukey’s测验。雌雄间比较采用独立样本检验。行为反应观察装置采用CAD绘图软件制作。

注:1:箱体;2:光源;3:趋光反应室;4:栖息室;5:避光反应室;11:左侧面;12:前隔板;13:前侧面;14:挡光板;15:箱盖;16:后隔板;18:白纱布;32:趋光通孔;34:趋光通孔盖板;52:避光通孔;54:避光通孔盖板
2 结果与分析
2.1 灰茶尺蠖成虫的趋光行为反应
雌虫的趋光率曲线呈现5个突出的峰(图2)。主峰位于橙光600~605 nm,与其他处理差异达显著水平。次峰位于绿光520~525 nm,第3峰位于紫光370~375 nm,第4个峰位于琥珀光595~600 nm,第5峰为雌虫对自然白光的趋光率。次峰与第3、4、5峰差异显著性处于同一水平。2个较小的峰分别位于紫光420~425 nm和390~395 nm处。以冰蓝青光(490~495 nm)和黄光(590~595 nm)的趋光率最低,且两者无显著性差异,其中黄光处理和其他处理均达显著水平。
雄虫的趋光率曲线呈现3个突出的峰(图2)。主峰出现的位置与雌虫次峰波长区相同,位于绿光520~525 nm,与其他波长处理差异达到显著水平。次峰位于紫光395~400 nm,第3峰位于紫光390~395 nm。次峰与第3峰差异显著性处于同一水平,与其他处理达到显著水平。几个较小的峰,分别位于紫光370~375 nm、385~390 nm,红光660~665 nm和橙光600~605 nm处,差异显著性处于同一水平,同时与紫光380~385 nm和琥珀光595~600 nm也处于同一显著水平。灰茶尺蠖雄虫趋光率较低的光谱有紫光400~405 nm、410~415 nm、420~425 nm、蓝光440~450 nm、冰蓝青光490~495 nm、红光620~625 nm和自然白光(CK),除与蓝光的460~465 nm和黄光590~595 nm也处于同一显著水平外,与其他处理均达差异显著水平。
在紫光395~400 nm、420~425 nm,黄光590~595 nm,琥珀光595~600 nm,橙光600~605 nm,红光660~665 nm和自然白光(CK)处理下,雌、雄成虫趋光率差异显著,其他处理均无差异(图2)。
2.2 灰茶尺蠖成虫的避光行为反应
雌虫的避光率曲线呈多峰型(图3)。主要表现为3个峰,主峰位于冰蓝青光490~495 nm,次峰分别位于蓝光440~450 nm和红光 660~665 nm,主峰和次峰的差异显著性处于同一水平。第3峰位于蓝光460~465 nm。3个较小的峰,分别出现在紫光385~390 nm、395~400 nm和红光620~625 nm,与次峰和第3峰差异显著性处于同一水平。雌成虫的避光率在黄光590~595 nm处最低,与其他处理差异达显著水平;其次为紫光400~405 nm,与其他处理差异均达显著水平。
雄虫的避光率均在50%以上,各峰值较接近,主要表现为8个突出的峰(图3)。主峰分别位于紫光400~405 nm、蓝光440~450 nm、冰蓝青光490~495 nm和自然白光区(CK)。次峰位于紫光420~425 nm和红光620~625 nm,第3峰位于蓝光460~465 nm和黄光590~595 nm。8个峰值的差异显著性处于同一水平。避光行为反应最小的为绿光520~525 nm和橙光600~605 nm,与其他处理差异达显著水平。
在紫光395~400 nm、400~405 nm、420~ 425 nm,黄光590~595 nm,橙光600~605 nm,红光660~665 nm和自然白光(CK)处理下,雌、雄成虫趋光率差异显著(图3)。

注:不同小写字母表示不同波长处理雌虫或雄虫间差异显著(P<0.05,F测验)。同行数据后不同大写字母表示雌雄间差异显著(P<0.05,t检验)

注:不同小写字母表示不同波长处理雌虫或雄虫间差异显著(P<0.05,F测验)。同行数据后不同大写字母表示雌雄间差异显著(P<0.05,t检验)
2.3灰茶尺蠖光谱行为反应的差异性
从雌雄成虫的趋、避光率曲线可以看出,避光率均明显高于趋光率,雌、雄之间存在明显的交叉(图2,图3)。整体来看,趋光率弱的波段,避光率相对较强。差异显著性分析表明,雌虫的趋光与避光反应,除黄光590~595 nm和橙光600~605 nm无差异外,其他光谱处理均差异显著(表1)。雄虫则表现为除绿光520~525 nm无差异外,其他光谱处理均差异显著(表2)。
3 讨论
利用昆虫趋光特性,采用灯光诱杀农田害虫是一种便捷有效的防治措施,既节能环保又可直接把害虫消灭在大发生之前[14]。本研究发现,在370~665 nm单色光照射下,灰茶尺蠖雌、雄成虫均能产生一定的趋、避光行为反应,其趋、避光反应曲线均为多峰型,且避光率明显高于趋光率。雌、雄成虫趋光率较高的光谱为绿光、紫光和橙光,该研究结果与杨心月等[9]研究二点委夜蛾雌、雄成虫对紫外光、橙光、绿光的趋光性结果一致,但与其对蓝光的趋性较强存在差异,可能与昆虫对光的反应存在差异有关。
郭祖国等[10]认为茶尺蠖成虫具有趋蓝色的习性,本研究中雌雄成虫对蓝色光谱的趋性均较低,可能与尺蠖种间差异有关。据报道,涂海华等[12]采用紫光和蓝光组成的LED双光谱太阳能杀虫灯,在380~400 nm波段内对茶尺蠖的诱杀量较高,而本研究对灰茶尺蠖具有较强吸引力的光谱为520~525 nm,其次为390~400 nm、370~375 nm和385~390 nm,与其趋光性一致,这可能与单色光谱和双光谱的组合差异有关。涂海华等[11]采用单波段诱杀结果显示,茶尺蠖趋光性较强的波段为365~410 nm,波段范围较广,与本研究中390~400 nm、370~375 nm和385~390 nm波段的趋光性相一致,但与本研究中趋光性较强的600~605 nm和520~525 nm光区不同,可能与涂海华等[11]采用的波段较少有关,具体原因还需进一步研究。

表1 灰茶尺蠖雌虫的趋、避光率差异显著性分析
注:表中不同字母表示同一波长下处理间差异显著性(<0.05,检验)
Note: Different small letters following the data in a column indicate significant differences among treatments (<0.05,-test)
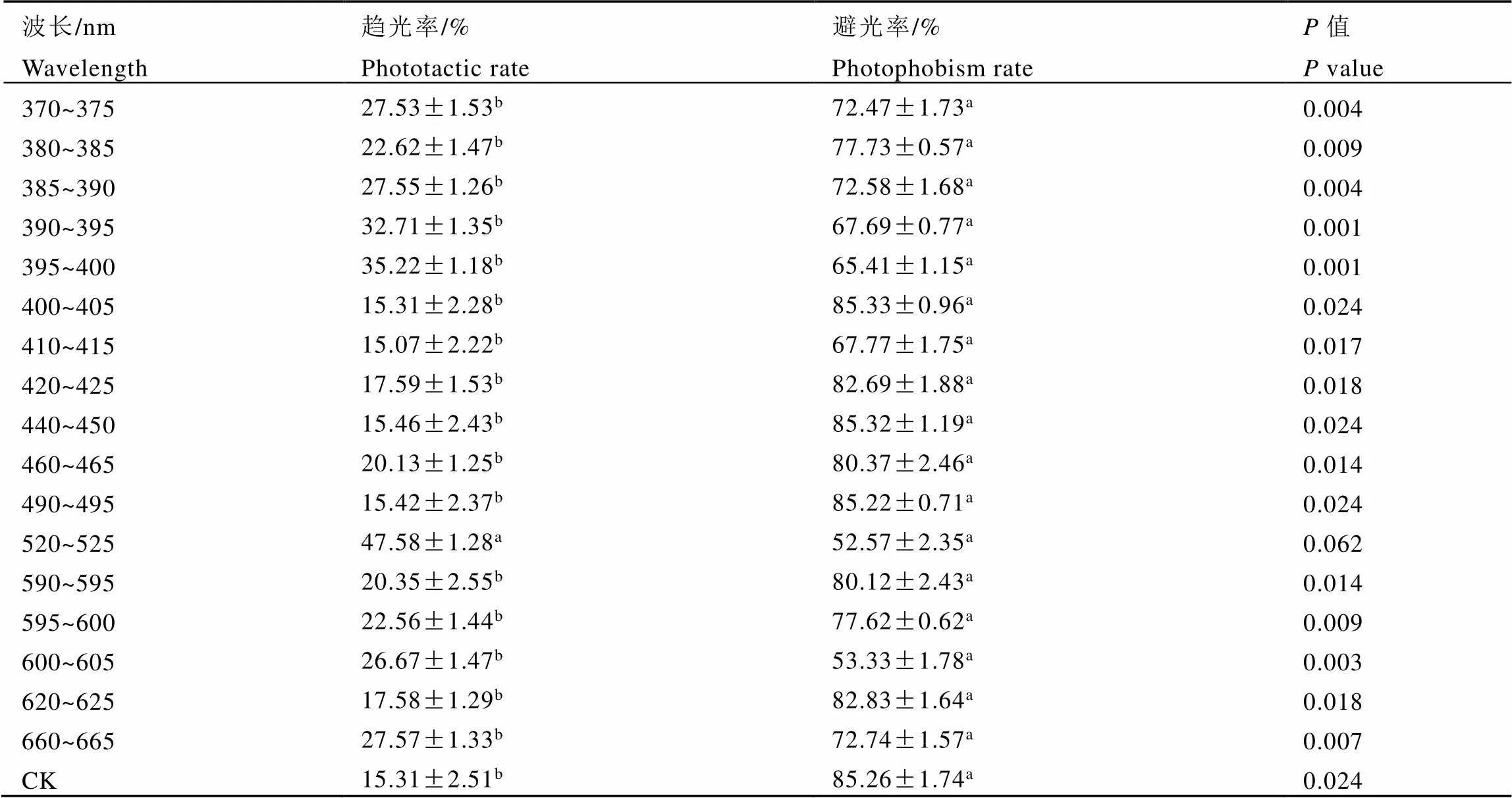
表2 灰茶尺蠖雄虫的趋、避光率差异显著性分析
注:表中不同字母表示同一波长下处理间差异显著(<0.05,检验)
Note: Different small letters following the data in a column indicate significant differences among treatments (<0.05,-test)
昆虫性别也影响其对光的行为反应。蒋月丽等[15]研究认为,雌虫的趋光率高于雄虫,且在某些波段具有显著性差异。本研究表明,灰茶尺蠖雌、雄成虫避光行为较高,趋向性相同的光区分别为紫光420~425 nm、蓝光440~ 450 nm和460~465 nm、冰蓝青光490~495 nm和红光620~625 nm,避光率均在80%以上。灰茶尺蠖雌、雄成虫的趋光率较高且重复的光区分别为紫光370~375 nm、390~395 nm,绿光520~525 nm和橙光600~605 nm。雄虫的趋光率低于雌虫。从总体的光反应率曲线可以看出,当趋光率大时,其避光率相对较小,原因可能是避光反应是由趋光行为的衍生或者随机活动所造成的[16]。
在茶叶生产中双波灯、频振灯、LED灯等诱杀方式多样[17],不仅可以监测害虫种群动态的发展趋势,还可以对害虫进行有效防控。目前生产上常用的诱虫灯大多是混合连续光源,虽然在害虫的监测和诱杀上能达到较好效果,但同时也对天敌等非靶标昆虫造成一定危害[9]。因此,根据害虫发生的特定敏感光谱设计研发专用的诱虫灯具有较大的现实意义。
[1] 张家侠, 孙钦玉, 葛超美, 等. 灰茶尺蠖雌成虫生殖器形态特征与组织结构的研究[J]. 茶叶科学, 2019, 39(1): 98-104. Zhang J X, Sun Q Y, Ge C M, et al. Morphological characteristics and histological structure of the reproductive organs in female adults ofWarren (Lepidoptera: Geometridae) [J]. Journal of Tea Science, 2019, 39(1): 98-104.
[2] Ma T, Xiao Q, Yu Y G, et al. Analysis of tea geometrid () pheromone gland extracts using GC-EAD and GC×GC/TOFMS [J]. Journal of Agricultural and Food Chemistry, 2016, 64(16): 3161-3166.
[3] Wang Z B, Ma T, Mao T F, et al. Application technology of the sex pheromone of the tea geometrid(Lepidoptera: Geometridae) [J]. International Journal of Pest Management, 2018, 64(4): 372-378.
[4] 张方梅, 刘书含, 陈利军, 等. 土荆芥精油对灰茶尺蠖幼虫的熏蒸活性及对其体内两种酶活力的影响[J]. 中国植保导刊, 2019, 39(4): 13-16, 43. Zhang F M, Liu S H, Chen L J, et al. Fumigant activity ofoil and its effects on acetylcholinesterase and carboxylesterase in[J]. China Plant Protection, 2019, 39(4): 13-16, 43.
[5] 罗宗秀, 苏亮, 李兆群, 等. 灰茶尺蠖性信息素田间应用技术研究[J]. 茶叶科学, 2018, 38(2): 140-145. Luo Z X, Su L, Li Z Q, et al. Field application technology of sex pheromoneon[J]. Journal of Tea Science, 2018, 38(2): 140-145.
[6] 林闽, 姚白云, 张艳红, 等. 太阳能LED杀虫灯的研究[J]. 可再生能源, 2007, 25(3): 79-80. Lin M, Yao B Y, Zhang Y H, et al. The studies of solar LED lights for killing worms [J]. Renewable Energy Resources, 2007, 25(3): 79-80.
[7] 刘晓英, 焦学磊, 郭世荣, 等. 基于LED诱虫灯的果蝇趋光性实验[J]. 农业机械学报, 2009, 40(9): 178-180. Liu X Y, Jiao X L, Guo S R, et al. Experiment of light emitting diode catching insect lamp on phototaxis of drosophila [J]. Transactions of the Chinese Society for Agricultural Machinery, 2009, 40(9): 178-180.
[8] 范凡, 任红敏, 吕利华, 等. 光谱和光强度对西花蓟马雌虫趋光行为的影响[J]. 生态学报, 2012, 32(6): 1790-1795. Fan F, Ren H M, Lv L H, et al. Effect of spectral sensitivity and intensity response on the phototaxis of(Pergande) [J]. Acta Ecologica Sinica, 2012, 32(6): 1790-1795.
[9] 杨心月, 范凡, 陈洁, 等. 光谱对二点委夜蛾成虫趋光行为的影响[J]. 植物保护学报, 2015, 42(6): 1009-1013. Yang X Y, Fan F, Chen J. et al. Effects of spectral sensitivity on the phototaxis ofadults [J]. Journal of Plant Protection, 2015, 42(6): 1009-1013.
[10] 郭祖国, 王梦馨, 崔林, 等. 昆虫趋色性及诱虫色板的研究和应用进展[J]. 应用生态学报, 2019, 30(10): 3615-3626. Guo Z G, Wang M X, Cui L, et al. Advance in insect phototaxis and the development and application of colored sticky boards [J]. Chinese Journal of Applied Ecology, 2019, 30(10): 3615-3626.
[11] 涂海华, 邝先飞, 毛宇, 等. 双波段LED太阳能杀虫灯对茶园害虫的控制作用研究[J]. 中国植保导刊, 2018, 38(10): 53-57. Tu H H, Kuang X F, Mao Y, et al. Effect of solar insecticidal lamp with dual-band LED on insect pests in tea plantation [J]. China Plant Protection, 2018, 38(10): 53-57.
[12] 涂海华, 胡秀霞, 毛宇, 等. 双光谱LED茶园专用杀虫灯的研制与应用[J]. 安徽农业科学, 2019, 47(3): 114-117. Tu H H, Hu X X, Mao Y, et al. Development and application of the dual spectrum LED insecticidal lamp for tea garden dedicated [J]. Journal of Anhui Agricultural Sciences, 2019, 47(3): 114-117.
[13] 王朝伟, 杨云秋, 钱六九, 等. 不同频振光源对皖南茶园主要昆虫的诱集效果[J]. 安徽农业大学学报, 2018, 45(5): 790-795. Wang C W, Yang Y Q, Qian L J, et al. Trapping effect of different light sources from frequency trembler grid lampon major insects in tea fields of southern Anhui [J]. Journal of Anhui Agricultural University, 2018, 45(5): 790-795.
[14] 周向平, 蒋笃忠, 沈力, 等. 太阳能诱虫灯诱杀烟草害虫的效果研究[J]. 湖南农业科学, 2012(19): 90-92. Zhou X P, Jiang D Z, Shen L, et al. Controlling effect of solar trapping lamp on tobacco pests [J]. Hunan Agricultural Sciences, 2012(19): 90-92.
[15] 蒋月丽, 武予清, 李彤, 等. 铜绿丽金龟对不同光谱的行为反应[J]. 昆虫学报, 2015, 58(10): 1146-1150. Jiang Y L, Wu Y Q, Li T, et al. Behavioural responses ofMotschulsky (Coleoptera: Scarabaeoidea) to different spectral light [J]. Acta Entomologica Sinica, 2015, 58(10): 1146-1150.
[16] 陈晓霞, 闫海燕, 魏玮, 等. 光谱和光强度对龟纹瓢虫成虫趋光行为的影响[J]. 生态学报, 2009, 29(5): 2349-2355. Chen X X, Yan H Y, Wei W, et al. Effect of spectral sensitivity and intensity response on the phototaxis of(Thunberg) [J]. Acta Ecologica Sinica, 2009, 29(5): 2349-2355.
[17] 边磊, 苏亮, 蔡顶晓.天敌友好型LED杀虫灯应用技术[J]. 中国茶叶, 2018, 40(2): 5-8. Bian L, Su L, Cai D X. Application technology of natural enemy friendly LED insecticide lamp [J]. China Tea, 2018, 40(2): 5-8.
Effects on the PhotoreactionBehavior ofWarren to DifferentLight Spectrum
QIAO Li1,2, HONG Feng1,2, JIN Yinli1,2, GENG Shubao1,2, GUO Shibao1,2*
1. Xinyang Agriculture and Forestry University, Xinyang 464000, China; 2. Henan Provincial South Henan Crop Pest Green Prevention and Control Academician Workstation, Xinyang 464000, China
The study aims to get the sensitive spectral range ofWarren to different light spectrum. The phototaxis or photophobism behavior ofWarren in response to spectral sensitivity was tested under 18 monochromatic lights ranging from 370 nm to 600 nm in the laboratory. The results show that phototaxis or photophobism reaction were observed under all the tested monochromatic wavelengths between 370 nm and 600 nm spectrum. The female adults showed positive phototaxis under orange light (600-605 nm), green light (520-525 nm) and ultraviolet light (370-375 nm). Meanwhile the male adults showed phototaxis under green light (520-525 nm), ultraviolet light (395-400 nm) and ultraviolet light (390-395 nm). The female adults showed photophobia under ice indigo light (490-495 nm), blue light (440-450 nm) and red light (660-665 nm). Meanwhile the male adults showed photophobia under ultraviolet light (400-405 nm), blue light (440-450 nm), ice indigo light (490-495 nm) and CK. There are differences inphototaxis ofWarren to light at different wavelengths, and gender is an important factor affecting the spectral behavior response. This study not only provided a necessary theoretical foundation for further research on light vision ofWarren, but also provided a scientific basis for the use of phototaxis in the integrated management ofWarren.
Warren, adult, light spectrum, phototaxis, photophobism
S571.1;S435.711
A
1000-369X(2020)05-617-08
2019-12-30
2020-02-12
国家重点研发计划“茶园化肥农药减施增效技术集成研究与示范”(2016YFD0200900)、信阳农林学院青年教师科研基金资助项目(2018LG003、201701007)
乔利,女,副教授,主要从事茶园害虫绿色防控方面的研究,qiaoli@xyafu.edu.cn。*通信作者:sbguo510@163.com
猜你喜欢
世界热带农业信息(2022年8期)2022-07-19
植物保护(2021年4期)2021-11-12
茶叶科学(2021年4期)2021-08-12
通信产业报(2019年31期)2019-10-21
中国知识产权(2018年11期)2018-11-29
爱你(2017年13期)2017-11-15
农民致富之友·下半月(2015年6期)2015-07-09
百科知识(2015年7期)2015-04-08
中国计算机报(2014年46期)2014-12-29
江苏农业科学(2014年3期)2014-07-16
