
外源5-氨基乙酰丙酸对低温胁迫下茶树叶片光合及生理特性的影响
2020-10-16 09:34:04燕飞蒋文华曲东付静赵璇
茶叶科学 2020年5期
燕飞,蒋文华,曲东,2,3,付静,赵璇
外源5-氨基乙酰丙酸对低温胁迫下茶树叶片光合及生理特性的影响
燕飞1,2,3,4,蒋文华1,曲东1,2,3,付静1,2,3,4,赵璇4
1. 陕西理工大学生物科学与工程学院,陕西 汉中 723000;2. 陕西省资源生物重点实验室,陕西 汉中 723000;3. 陕南秦巴山区生物资源综合开发协同创新中心,陕西 汉中 723000;4. 陕西理工大学秦巴红茶研究所,陕西 汉中 723000
通过对陕茶1号(耐低温型)和金牡丹(低温敏感型)2个茶树品种在冬季自然低温胁迫下喷施不同浓度(0、10、30、50 mg·L-1)的外源5-氨基乙酰丙酸(5-aminolevulinic acid,ALA),探究外源ALA对低温胁迫下茶树叶片光合荧光特性及生理特性的调控作用。结果表明,适宜浓度的外源ALA对低温胁迫下茶树叶片净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、PSⅡ最大光化学效率及PSⅡ潜在活性具有促进作用,能够提高水浸出物、咖啡碱、游离氨基酸、儿茶素类物质、茶氨酸、可溶性糖、可溶性蛋白等生理活性物质含量的累积;外源ALA能够提高低温胁迫下茶树光合作用的能力并改善茶叶品质,其中50 mg·L-1的ALA处理能有效提高陕茶1号应对低温胁迫的能力,10 mg·L-1和30 mg·L-1的外源ALA对金牡丹应对低温胁迫具有较好的缓解效用。
5-氨基乙酰丙酸;茶树;低温胁迫;光合特性;生理特性
陕南属于我国江北茶区,冬季气温偏低、持续时间长(图1),冬季低温和早春“倒春寒”对该地区茶树越冬和春季名优茶生产造成很大的负面影响。低温冻害会使茶树叶片变红,出现黑色斑点、焦枯、卷缩、易脱落等现象,甚至导致茶树死亡,严重影响茶叶的产量和品质,造成重大的经济损失[1]。因此,如何提高茶树抵御低温的能力,减轻低温胁迫对茶叶的产量及品质的影响逐渐成为了茶叶越冬栽培的研究热点。
5-氨基乙酰丙酸是四吡咯类化合物的前体物质,是一种广泛存在于微生物、植物和动物细胞中的非蛋白类氨基酸[2],参与调节叶绿素的合成与代谢[3]。低浓度的ALA具有促进植物光合作用、提高PSⅡ反应活性、促进物质累积等作用,可以作为植物生长调节剂促进作物生长发育,缓解非生物胁迫对植物造成的伤害[4-8]。现有研究表明,外源ALA能够有效缓减干旱[9]、盐渍[10]、低温[11]等非生物胁迫对植物生长所产生的抑制效应,然而在提高茶树抗寒方面的研究尚未见报道。因此,本试验以陕茶1号(耐低温型)和金牡丹(低温敏感型)为试验对象,喷施不同浓度的外源ALA,研究外源ALA对低温胁迫下茶树叶片光合及其生理特性的影响,探讨外源ALA对低温胁迫下茶树光合调控及品质积累的作用机制,为茶树抗寒研究提供理论基础和技术依据。
1 材料和方法
1.1 试验材料
试验场地位于陕西省汉中市南郑区牟家坝镇汉中市农业科学研究所茶树种质资源圃(海拔650 m,冬季常年平均气温低于4℃,极端天气可达–14℃),供试材料为3年生的陕茶1号(耐低温型)和金牡丹(低温敏感型)。试验小区长10 m,宽1.5 m,共栽培60株茶树,采用随机区组设计,每个处理3个小区。
1.2 试验方法
1.2.1 材料处理方法
试验设4个处理,分别为0(喷施等量清水为对照)、10、30 mg·L-1和50 mg·L-1的ALA溶液(基于预试验结果设置)。将不同浓度的ALA溶液每隔7 d喷施于茶树正反叶面上,喷施标准以有水滴滴下为准,喷施时间为2019年2月11日至3月18日,共喷施6次。于3月25日(春茶采收生产前期,且天气晴朗)早上8:30—11:30,每个小区随机选取10株茶树测定光合气体交换参数,测定叶片均为顶芽下第三、四片无损伤、无病害、大小相近的健康叶片,随后采集第四叶位以上健康新梢叶片固样后粉碎,密封保存至–20℃冰箱,用于测定生理活性物质。

图1 试验场地2019年1月至3月最低温度
1.2.2 指标检测方法
使用美国LI-COR公司生产的Li-6400XT便携式光合仪对光合参数净光合速率()、胞间CO2浓度()、气孔导度()、蒸腾速率()等进行测定,并根据公式=/和=1-/,计算瞬间水分利用率()和气孔限制值(),其中,为叶外空气CO2浓度。叶室温度设定为28℃,照射光强为1 000 μmol·m-2·s-1,流速为500 μmol·mol-1。
固样:将采集的叶片大约每100 g鲜叶用蒸锅蒸青90 s,冷却至常温后于80℃烘箱烘干2 h,之后于50℃烘至茶叶可捏碎状态,用粉碎机粉碎后过40目筛,密封保存置于–20℃冰箱备用[12]。
干物质含量测定采用GB/T 8303—2013[13],水浸出物含量测定采用GB/T 8305—2013[13],游离氨基酸测定采用GB/T 8314—2013[13],茶氨酸测定采用GB/T 23193—2017[13],可溶性糖含量测定采用蒽酮–硫酸法[14],可溶性蛋白含量测定采用考马斯亮蓝-G250染色法[14],参考薛金金等[15]方法测定儿茶素类物质和咖啡碱含量。
将叶片暗适应20 min后使用OS-30P+植物胁迫快速测量仪测定PSⅡ最大光化学效率(/)和PSⅡ潜在活性(/)。
1.3 数据处理
所测数据采用Microsoft Excel 2010进行处理,使用IBM SPSS Statistics 20软件进行Duncan’s多重比较分析(<0.05),使用GraphPad Prism 5进行作图。
2 结果与分析
2.1 外源ALA对茶树叶片光合气体交换参数的影响
光合作用是植物将太阳能转化为化学能并将无机物转化为有机物的过程。光合速率的测量是研究植物光合性能、光合机构运转及环境因素对光合作用影响的重要方法。由表1可知,陕茶1号光合气体交换参数的、、、等指标随外源喷施ALA浓度升高呈上升趋势,当外源ALA浓度为50 mg·L-1时均达到最大值,与对照相比,分别提高了30.99%、46.67%、5.08%和31.47%,差异显著。随着ALA浓度升高呈先下降后上升趋势,浓度为30 mg·L-1时下降幅度最大,与对照相比降低了17.39%,而随外源ALA浓度升高并未出现显著差异。金牡丹的、、和变化趋势随外源ALA浓度升高呈先上升后下降趋势,浓度为10 mg·L-1时达到最大值,分别比对照提高了26.10%、23.53%、8.31%和61.25%,差异显著。随外源ALA浓度升高呈先下降后上升趋势,浓度为30 mg·L-1时下降幅度最大,与对照相比显著下降了19.23%。10 mg·L-1外源ALA处理下,金牡丹与其他处理存在显著性差异,相比对照显著降低了21.93%。
2.2 外源ALA对茶树叶片叶绿素荧光参数的影响
叶绿素荧光参数的变化在一定程度上可以反映植物在遇到环境胁迫时做出的响应,/和/值是反映茶树叶片的最大光能转化效率和光系统Ⅱ的潜在活性。如图2所示,外源ALA处理均显著增加了陕茶1号PSⅡ最大光化学效率,且PSⅡ最大光化学效率随着浓度升高呈上升趋势,当浓度为50 mg·L-1时达到最大值,比对照组显著增加了6.1%。随着外源ALA浓度升高,金牡丹的PSⅡ最大光化学效率呈先上升后下降趋势,浓度为10 mg·L-1达到最大值,较对照组提高5.4%,差异显著。
由图3可知,外源ALA浓度为30 mg·L-1和50 mg·L-1时,陕茶1号PSⅡ潜在活性与对照组存在显著性差异,分别比照组高13.4%和9.1%。与陕茶1号不同,3个浓度外源ALA处理均显著提高了金牡丹的PSⅡ潜在活性,10、30 mg·L-1和50 mg·L-1的外源ALA分别比对照组高21.1%、12.6%、13.4%,而ALA各浓度处理之间并无显著性差异。
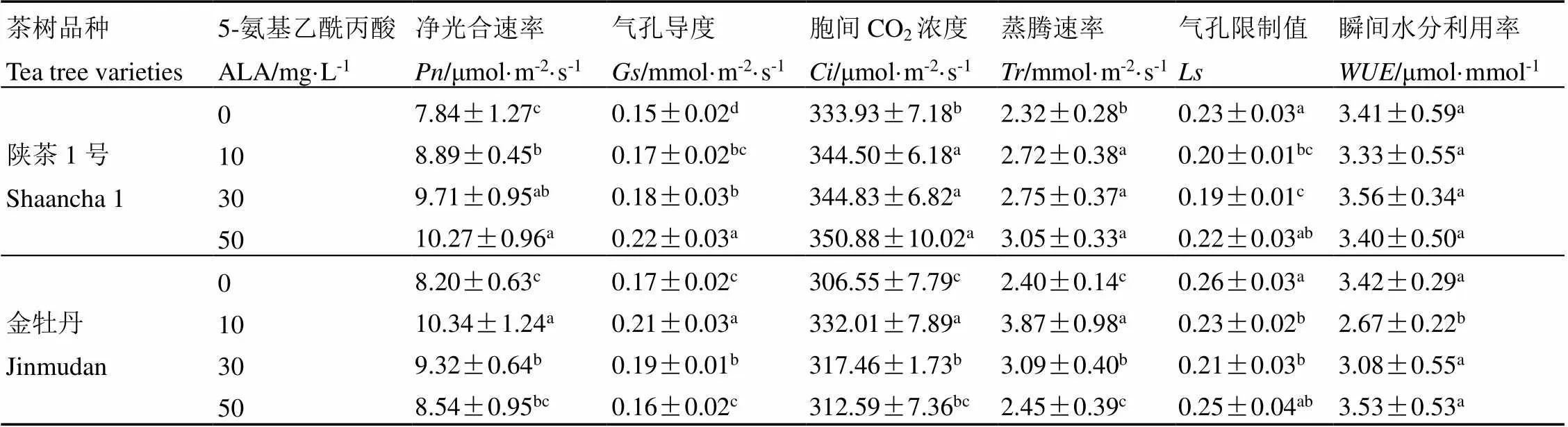
表1 外源5-氨基乙酰丙酸对茶树叶片光合气体交换参数的影响
注:2个品种每列不同的小写字母表示在0.05水平有显著差异。下同
Note: Different lowercase letters in each column of the two cultivars indicate significant difference at the level of 0.05. The same below
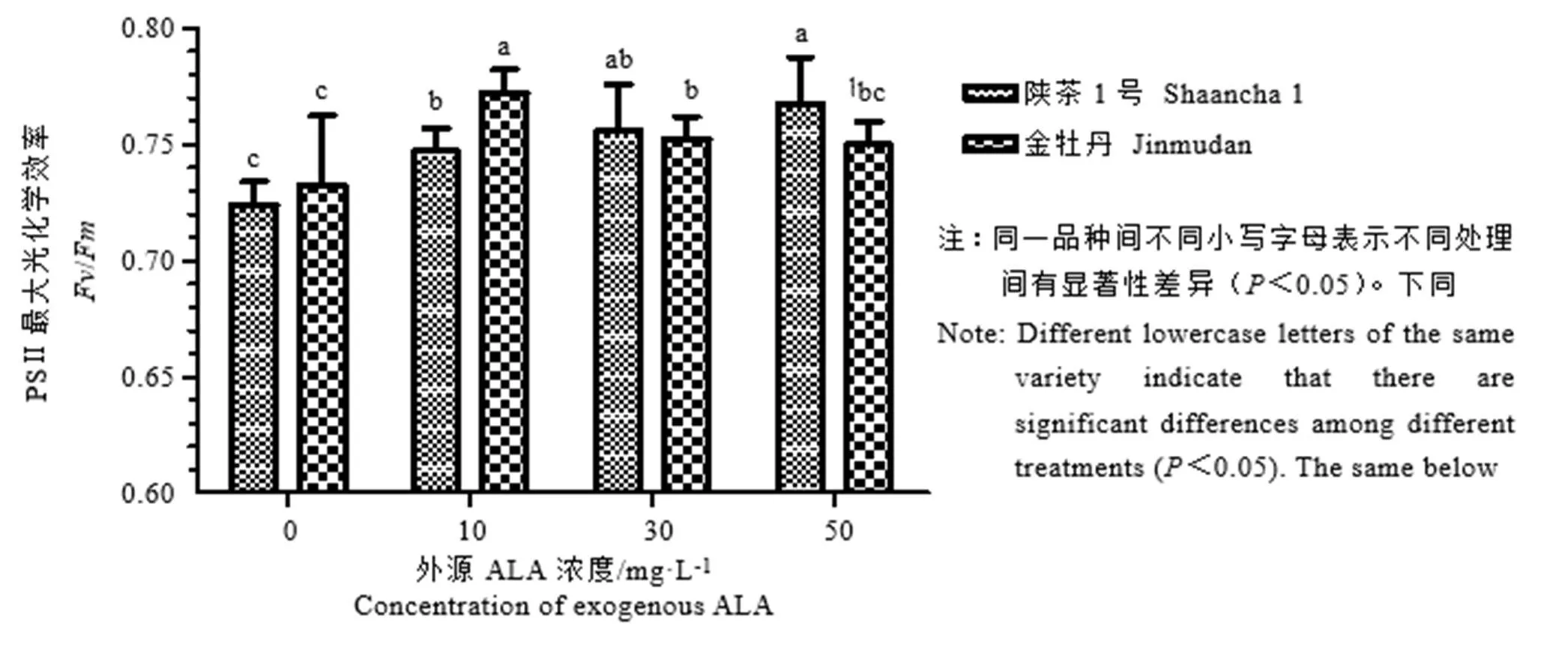
图2 外源5-氨基乙酰丙酸茶树叶片PSⅡ最大光化学效率的影响
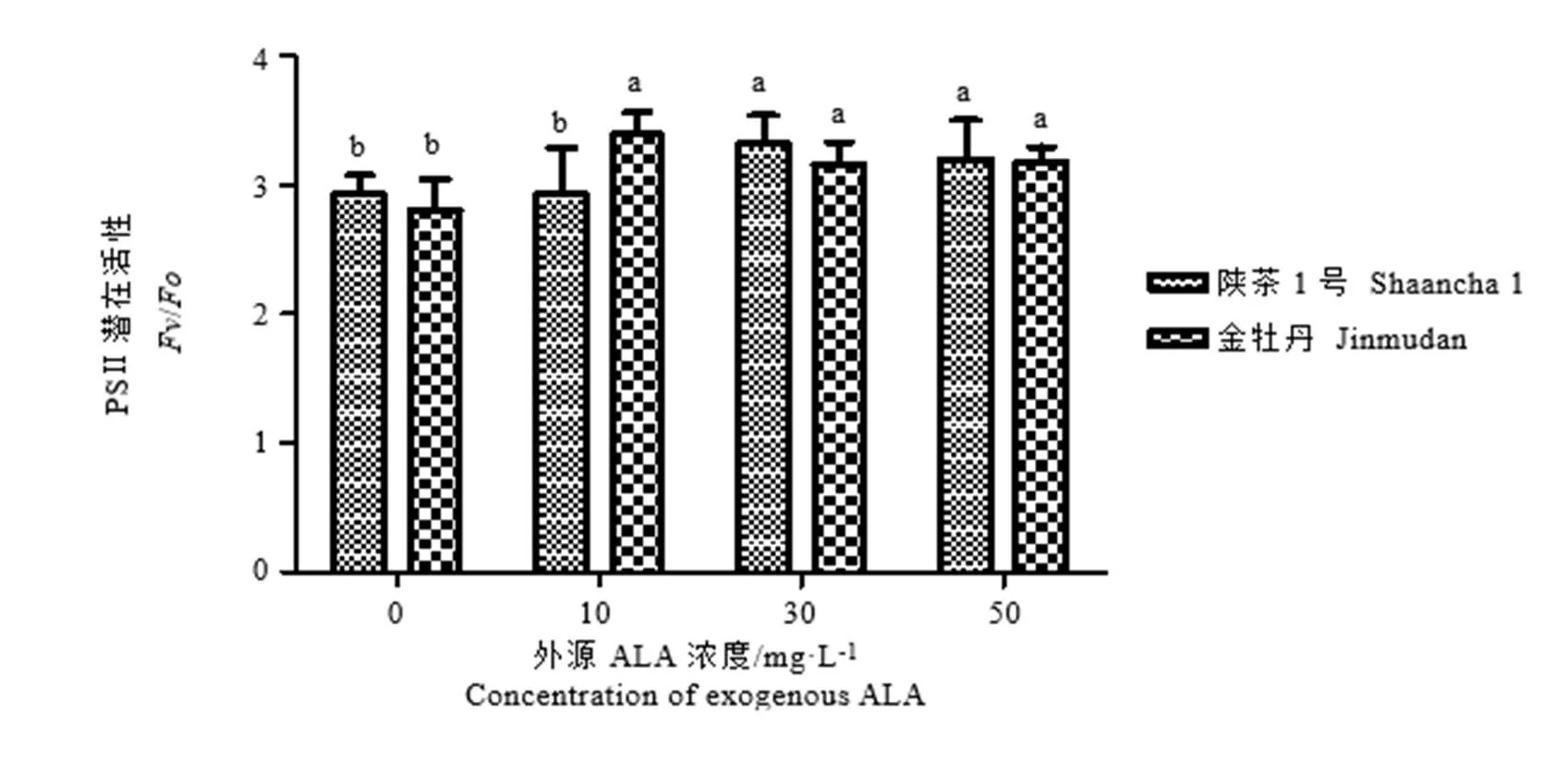
图3 外源5-氨基乙酰丙酸对低温胁迫下茶树叶片PSⅡ潜在活性的影响
2.3 外源ALA对茶树叶片生理特性的影响
从表2可以看出,30 mg·L-1和50 mg·L-1的外源ALA处理均显著提高了陕茶1号干物质含量,其中50 mg·L-1的ALA效果最佳;3个外源ALA浓度处理对水浸出物的影响差异不显著;咖啡碱、游离氨基酸、儿茶素类物质、茶氨酸等含量随着外源ALA浓度升高呈上升趋势,浓度为50 mg·L-1时达到最大值,分别比对照高出17.87%、4.35%、27.79%和12.26%,且差异显著。外源ALA处理提高了金牡丹干物质、水浸出物、咖啡碱等内含物的含量,随着处理浓度的升高呈先上升后下降趋势,30 mg·L-1的外源ALA最大限度的提高了金牡丹茶树叶片的生理活性成分含量,其中水浸出物、咖啡碱、游离氨基酸、儿茶素类物质和茶氨酸含量分别较对照组高20.23%、27.80%、24.06%、17.86%和68.78%,且存在显著差异。
可溶性糖与可溶性蛋白不仅能为茶叶提供甜味,以及保持茶汤清亮,而且参与植物渗透调节,维持细胞内渗透势,具有维持细胞稳定的功能。从图4可以看出,经ALA处理后,陕茶1号和金牡丹的可溶性糖含量随外源ALA浓度升高呈先上升后下降趋势,ALA浓度为30 mg·L-1时上升幅度最大,与对照相比分别上升了26.94%和47.49%,差异显著。不同浓度对两个不同耐低温型茶树品种可溶性糖无显著影响。

表2 外源5-氨基乙酰丙酸对茶树叶片生理活性成分的影响

图4 外源5-氨基乙酰丙酸对茶树叶片可溶性糖的影响
如图5所示,喷施不同浓度外源ALA对陕茶1号和金牡丹可溶性蛋白含量的影响存在差异。喷施不同浓度外源ALA后,陕茶1号的可溶性蛋白含量明显高于对照组,且随着ALA处理浓度的升高呈上升趋势,浓度为50 mg·L-1时达到最大值,较对照显著提高了113.70%。金牡丹可溶性蛋白含量随着外源ALA浓度升高呈先上升后下降趋势,10 mg·L-1和30 mg·L-1的外源ALA处理使可溶性蛋白含量较对照分别提高33.82%和51.56%,当浓度为50 mg·L-1时其可溶性蛋白含量下降至对照水平。
3 讨论与结论
植物光合作用是有机物的主要来源,光合作用可为茶树碳、氮代谢提供碳源和能量[16-17],温度是影响茶树进行光合作用的主要生态因子之一[18]。大量研究表明,低温降低了植物的、、、和/,从而抑制其光合作用,进一步影响植物的生长[19-23]。本试验结果显示,与对照相比,喷施一定浓度的外源ALA可以提高陕茶1号和金牡丹茶树的、、、,降低其和,这与低温胁迫下茶树[23]、玉米幼苗[5]和辣椒[24]植株的研究结果一致。低温胁迫可直接损伤茶树叶片的PSⅡ反应中心,导致茶树光合作用能力减弱。ALA作为叶绿素合成的前体物质,能够促进低温胁迫下茶树光合作用,提高PSⅡ反应活性,提高植物应对低温胁迫的能力。光合速率下降原因主要由和的变化方向来判断,降低和升高,表明气孔导度降低是光合速率降低的主要因素,增高和降低表明光合速率降低的原因是非气孔因素[25]。本试验中外源ALA处理使陕茶1号升高,和与变化趋势一致,降低;金牡丹随外源ALA处理浓度增大表现为先增高后降低,和与变化趋势一致,表明气孔导度是影响茶树光合速率的主要因素,外源ALA促进了茶树气孔开放,叶片通过气孔吸收环境中的CO2,加快CO2和H2O的交换速率,从而提高光合速率。Wang等[26]发现外源ALA能够提高冷胁迫下大甜瓜叶片的气孔导度,进而提高光合速率;张治平等[27]发现转基因烟草中过量合成ALA可以诱导叶片气孔开放,促进CO2进入叶肉细胞,从而提高植物叶片净光合速率;陈令会等[28]进一步验证了ALA能够促进光、暗条件下苹果叶片气孔开放,并阻止外源ABA和H2O2诱导的气孔关闭,逆转Ca2+诱导的气孔关闭。而另一方面,ALA作为叶绿体的前体物质,参与叶绿素的合成代谢,叶绿素是植物进行光合作用的主要色素,其含量高低可以反映植物光合作用能力。有研究表明,外源ALA能够提高春茶叶绿素含量[29],因此推测,外源ALA能够促进低温胁迫下茶树叶绿素合成,从而缓解低温胁迫对光合作用的抑制,提高茶树应对低温胁迫的能力。而外源ALA提高茶树光合速率可能受气孔因素和非气孔因素共同作用,此猜测仍需进一步验证。此外,本研究中外源ALA处理后陕茶1号和金牡丹/和/显著高于对照组,这与康琅等[30]对西瓜叶片的研究结果一致,表明外源ALA可以提高茶树叶片PSⅡ原初光能转化效率,减轻低温对PSⅡ反应中心损伤程度,促进叶片PSⅡ捕获光能,有利于光合电子传递效率。

图5 外源5-氨基乙酰丙酸对茶树叶片可溶性蛋白的影响
咖啡碱、氨基酸、儿茶素类物质、糖类、蛋白质等是评价茶叶风味品质成分的主要指标[31],这些风味品质成分共同决定着茶叶的滋味与口感,其含量决定着茶叶品质的优劣[32-34]。郭湘等[35]研究发现,与正常茶样相比,受冻茶样的水浸出物、游离氨基酸、儿茶素、咖啡碱均有不同程度的下降。本研究通过对2个品种茶树外源喷施一定浓度ALA的试验发现,与低温胁迫对照相比,3个浓度的外源ALA均提高了2个茶树品种的水浸出物、咖啡碱、游离氨基酸、儿茶素类物质、茶氨酸等含量的积累,这与安玉艳等[29]研究外源ALA对春茶生长与品质影响的试验结果一致,可能是外源ALA缓减了低温胁迫对茶树的生长抑制,维持了细胞稳定,缓减了低温对细胞膜的伤害,促进了光合作用能力,提高光合作用转化水平,有利于有机物质的转化。茶鲜叶中蛋白质的含量占干物质总量的20%左右,它对保持茶汤清亮和茶汤胶体溶液的稳定性起着重要作用。有研究发现,茶树叶片中可溶性糖含量随低温胁迫时间延长呈上升趋势,可溶性蛋白含量随着低温胁迫的时间延长呈现先降低后升高的趋势[36],本试验中,茶树通过外源ALA处理后,可溶性糖和可溶性蛋白含量均显著高于对照组,表明外源ALA能够提高逆境环境下植物可溶性糖和可溶性蛋白含量,这与外源ALA缓解盐胁迫对颠茄生长抑制[37]和促进百合生长发育[38]的研究结果一致,究其原因,可能是外源ALA促进了水解酶类的活性,水解酶使碳水化合物大量水解形成可溶性糖,协调茶汤滋味,单宁酶将茶叶中的多酚类没食子酸水解成咖啡碱和没食子多酚类物质。此外,低温条件下外源ALA可能促进诱导产生新的蛋白使可溶性蛋白的含量增加,蛋白质在蛋白酶的催化下水解成游离氨基酸,改善茶叶的鲜爽度。龙志荣等[39]研究发现,水解酶类与茶叶中可溶性糖、氨基酸、酚类物质等密切相关,如水解酶中的纤维素酶浓度增加,单丛茶的水浸出物、氨基酸、可溶性糖、多酚类、咖啡碱的含量均显著增加,水浸出物、氨基酸、多酚类含量随蛋白酶浓度增加而增加。本研究通过对2个不同温度敏感型的茶树品种喷施3个浓度的外源ALA发现,不同品种对于不同浓度的外源ALA响应不同。50 mg·L-1的外源ALA处理对陕茶1号光合作用促进效果最佳,而金牡丹则在浓度为10~30 mg·L-1范围内响应强烈,产生这一差异可能与植物自身的形态、内含物含量以及对外界低温环境的响应不同等有关。植物的光合作用和光合活性会随温度变化而变化,而光合作用对温度的敏感性与植物物种有密切关系,冷敏感的马铃薯在温度从30℃降低到4℃时光饱和下的CO2吸收速率下降75%,最大量子效率降低50%,暴露于低温下超过1 h则会发生不可逆的光合抑制现象,而抗冷性强的菠菜几乎未受到影响[40],可见,低温敏感型物种其光合作用对温度的响应更为敏感。低温敏感型植物冰点较低,低温胁迫会首先对细胞造成损伤,而在同等温度下耐寒性物种未出现结冰现象[41]。植物遭受胁迫伤害时,细胞膜会受到损伤,产生脱水现象,此时植物会主动形成渗透调节物质,调整水分吸收,因此渗透调节物质含量增加,低温敏感型植物会在低温条件下产生更多的渗透调节物质来应对低温环境,这也是低温条件下金牡丹可溶性糖和可溶性蛋白含量略高于陕茶1号的原因。
综上所述,适宜浓度的外源ALA有利于低温胁迫条件下茶树进行光合气体交换,促进光合作用,减轻低温对PSⅡ反应中心损伤程度,从而提高茶树应对低温胁迫的能力,并提高茶树叶片对光能的吸收及利用,茶叶中咖啡碱、游离氨基酸、儿茶素类物质、可溶性糖、可溶性蛋白含量等生理活性成分含量增加,改善了茶叶品质。然而不同品种的茶树,由于对外界环境信号的应答不同,导致外源ALA对茶树的作用效果产生浓度差异,具体响应机理,有待进一步深入分析和研究。
致谢:感谢汉中市农业科学研究所席彦军研究员为本试验提供场地和现场指导,感谢汉中市质量技术检验检测中心张亦凡工程师在茶叶活性物质测定方面给予的帮助和指导。
[1] 韩文炎, 李鑫, 颜鹏, 等. 茶园“倒春寒”防控技术[J]. 中国茶叶, 2018, 40(2): 9-12. Han W Y, Li X, Yan P, et al. Prevention and control technology of ‘late spring cold’ in tea garden [J]. China Tea, 2018, 40(2): 9-12.
[2] 李智祥, 赵磊, 梁云龙, 等. 生物法合成5-氨基乙酰丙酸的研究进展[J]. 发酵科技通讯, 2017, 46(3): 178-182. Li Z X, Zhao L, Liang Y L, et al. Advance on biosynthesis of 5-aminolevulinic acid [J]. Bulletin of Fermentation Science and Technology, 2017, 46(3): 178-182.
[3] Wu Y, Liao B W, Mohammed M D, et al. 5-aminolevulinic acid (ALA) biosynthetic and metabolic pathways and its role in higher plants: a review [J]. Plant Growth Regulation, 2019, 87(2): 357-374.
[4] Hotta Y, Tananka T, Bingshan L, et al. Improvement of cold resistance in rice seedlings by 5-aminolevulinic acid [J]. Journal of Pesticide Science, 1998, 23(1): 29-33.
[5] 孙阳, 王燚, 孟瑶, 等. 外源5-氨基乙酰丙酸对低温胁迫下玉米幼苗生长及光合特性的影响[J]. 作物杂志, 2016(5): 87-93. Sun Y, Wang Y, Meng Y, et al. Effects of exogenous ALA on growth and photosynthetic characteristics of maize seedlings under low temperature stress [J]. Crops, 2016(5): 87-93.
[6] Fan H F, Du C X, Guo S R. Effect of nitric oxide on proline metabolism in cucumber seedlings under salinity stress [J]. Journal of the American Society for Horticultural Science, 2012, 137(3): 127-133.
[7] Fan H F, Du C X, Guo S R. Nitric oxide enhances salt tolerance in cucumber seedlings by regulating free polyamine content [J]. Environmental and Experimental Botany, 2013, 86: 52-59.
[8] Fu J J, Sun Y F, Chu X T, et al. Exogenous 5-aminolevulenic acid promotes seed germination inagainst oxidative damage induced by cold stress [J]. Plos One, 2014, 9(9): e107152. doi: 10.1371/journal.pone.0107152.
[9] 王宏信, 剧春晖, 李向林, 等. 外源5-氨基乙酰丙酸对干旱胁迫下降香黄檀幼苗生长、根系生理特性的影响[J]. 热带作物学报, 2017, 38(10): 1823-1829. Wang H X, Ju C H, Li X L, et al. Effects of exogenous 5-aminolevulinic acid on growth and root physiological characteristics ofT.chen under drought stress [J]. Chinese Journal of Tropical Crops, 2017, 38(10): 1823-1829.
[10] 孟长军, 杜喜春, 张旸. 5-氨基乙酰丙酸对喜树幼苗盐害缓解的生理机制研究[J]. 热带亚热带植物学报, 2019, 27(2): 164-170. Meng C J, Du X C, Zhang Y. Study on physiological mechanism of alleviating effect of 5-aminolevulinic acid onseedlings under salt stress [J]. Journal of Tropical and Subtropical Botany, 2019, 27(2): 164-170.
[11] 李海燕, 毕文双, 王燚, 等. 外源ALA对低温胁迫下玉米幼苗根系生长及生理特性的影响[J]. 东北农业大学学报, 2019, 50(2): 9-17. Li H Y, Bi W S, Wang Y, et al. Effect of exogenous ALA on root growth and physiological characteristics of maize seedlings under low temperature stress [J]. Journal of Northeast Agricultural University, 2019, 50(2): 9-17.
[12] 赵璇, 李新生, 邢丽红, 等. 高效液相色谱法测定汉中茶叶中茶氨酸含量[J]. 食品安全质量检测学报, 2015, 6(12): 4902-4907. Zhao X, Li X S, Xing L H, et al. Determination of theanine in Hanzhong tea plantation by high performance liquid chromatography [J]. Journal of Food Safety and Quality, 2015, 6(12): 4902-4907.
[13] 中国标准出版社第一编辑室. 茶叶标准汇编[M]. 5版. 北京: 中国标准出版社, 2014. The first editorial office of China Standards Press. Collection of tea standards [M]. 5th ed. Beijing: Standards Press of China, 2014.
[14] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006. Gao J F. Experimental guidance of plant physiology [M]. Beijing: Higher Education Press, 2006.
[15] 薛金金, 江和源, 龙丹, 等. HPLC法同时测定茶叶中聚酯型儿茶素和茶黄素[J]. 中国食品学报, 2014, 14(5): 237-243. Xue J J, Jiang H Y, Long D, et al. Simultaneous multiresidue determination of theasinensins and theaflavins in tea using high performance liquid chromatography [J]. Journal of Chinese Institute of Food Science and Technology, 2014, 14(5): 237-243.
[16] 田纪春, 王学臣, 刘广田. 植物的光合作用与光合氮、碳代谢的耦联及调节[J]. 生命科学, 2001, 13(4): 145-147. Tian J C, Wang X C, Liu G T. The coupling and regulation between photosynthesis and nitrogen, carbon metabolism in plant [J]. Chinese Bulletin of Life Sciences, 2001, 13(4): 145-147.
[17] 张兰, 魏吉鹏, 沈晨, 等. 秋茶光合作用与品质成分变化的分析[J]. 茶叶科学, 2018, 38(3): 271-280. Zhang L, Wei J P, Shen C, et al. Analysis of the photosynthesis and quality components changes in autumn tea [J]. Journal of Tea Science, 2018, 38(3): 271-280.
[18] 陈芳, 刘宇鹏, 谷晓平, 等. 低温对茶树光合特性及产量的影响[J]. 作物杂志, 2018(3): 155-161. Chen F, Liu Y P, Gu X P, et al. Effects of low temperature on photosynthetic characteristics and yield of tea (L.) [J]. Crops, 2018(3): 155-161.
[19] 崔波, 程邵丽, 袁秀云, 等. 低温胁迫对白及光合作用及叶绿素荧光参数的影响[J]. 热带作物学报, 2019, 40(5): 891-897. Cui B, Cheng S L, Yuan X Y, et al. Effects of low temperature stress on the photosynthetic characteristics and chlorophyll fluorescence parameters of[J]. Chinese Journal of Tropical Crops, 2019, 40(5): 891-897.
[20] 周超凡, 吴春涛, 李丹丹, 等. 外源H2S通过减轻低温光抑制增强黄瓜幼苗耐冷性[J]. 植物生理学报, 2018, 54(3): 411-420. Zhou C F, Wu C T, Li D D, et al. Hydrogen sulfide promotes chilling tolerance of cucumber seedlings by alleviating low-temperature photoinhibition [J]. Plant Physiology Journal, 2018, 54(3): 411-420.
[21] 吴雪霞, 朱宗文, 张爱冬, 等. 外源褪黑素对低温胁迫下茄子幼苗生长及其光合作用和抗氧化系统的影响[J]. 西北植物学报, 2017, 37(12): 2427-2434. Wu X X, Zhu Z W, Zhang A D, et al. Effects of exogenous melatonin on the growth, photosynthesis and antioxidant system in eggplant (L.) seedlings under low temperature stress [J]. Acta Botanica Boreali Occidentalia Sinica, 2017, 37(12): 2427-2434.
[22] 余丽玲, 何天友, 陈凌艳, 等. 水杨酸对低温胁迫下西洋杜鹃光合作用和叶绿素荧光的影响[J]. 江西农业大学学报, 2014, 36(3): 520-525. Yu L L, He T Y, Chen L Y, et al. Effects of salicylic acid on photosynthesis and chlorophyll fluorescence of rhododendron hybridum under low temperature stress [J]. Acta Agriculturae Universitatis Jiangxiensis, 2014, 36(3): 520-525.
[23] 李庆会, 徐辉, 周琳, 等. 低温胁迫对2个茶树品种叶片叶绿素荧光特性的影响[J]. 植物资源与环境学报, 2015, 24(2): 26-31. Li Q H, Xu H, Zhou L, et al. Effect of low temperature stress on chlorophyll fluorescence characteristics in leaf of[J]. Journal of Plant Resources and Environment, 2015, 24(2): 26-31.
[24] 徐刚, 刘涛, 高文瑞, 等. ALA对低温胁迫下辣椒植株生长及光合特性的影响[J]. 江苏农业学报, 2011, 27(3): 612-616. Xu G, Liu T, Gao W R, et al. Effects of 5-aminolevulinic acid (ALA) on growth and photosynthesis of pepper plants under low temperature stress [J]. Jiangsu Journal of Agricultural Sciences, 2011, 27(3): 612-616.
[25] 许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯, 1997(4): 241-244. Xu D Q. Some Problems in stomatal limitation analysis of photosynthesis [J]. Plant Physiology Journal, 1997(4): 241-244.
[26] Wang L J, Jing W B, Huang B J. Promotion of 5-aminolevulinic acid on photosynthesis of melon () seedlings under low light and chilling stress conditions [J]. Physiologia Plantarum, 2004, 121(2): 258-264.
[27] 张治平, 汪良驹, 姚泉洪. 过量合成ALA转基因烟草叶片光合与叶绿素荧光特性的研究[J]. 西北植物学报, 2008(6): 1196-1202. Zhang Z P, Wang L J, Yao Q H. Study on leaf photosynthesis and chlorophyll fluorescence of transgenic tobacco over-producing 5-aminolevulinic acid (ALA) [J]. Acta Botanica Boreali-Occidentalia Sinica, 2008(6): 1196-1202.
[28] 陈令会, 刘龙博, 安玉艳, 等. 外源5-氨基乙酰丙酸促进苹果叶片气孔开放机理的初探[J]. 园艺学报, 2014, 41(10): 1965-1974. Chen L H, Liu L B, An Y Y, et al. Preliminary studies on the possible mechanism underlying 5-aminolevulinic acid-induced stomatal opening in apple leaves [J]. Acta Horticulturae Sinica, 2014, 41(10): 1965-1974.
[29] 安玉艳, 冯新新, 丁恒毅, 等. 5-氨基乙酰丙酸(5-ALA)对春茶生长与品质成分的影响[J].江苏农业科学, 2016, 44(11): 224-228. An Y Y, Feng X X, Ding H Y, et al. Effects of 5-aminolevulinic acid on the growth and quality components of spring tea [J]. Jiangsu Agricultural Sciences, 2016, 44(11): 224-228.
[30] 康琅, 汪良驹. ALA对西瓜叶片叶绿素荧光光响应曲线的影响[J]. 南京农业大学学报, 2008, 31(1): 31-36. Kang L, Wang L J. Effects of ALA treatments on light response curves of chlorophyll fluorescence of watermelon leaves [J]. Journal of Nanjing Agricultural University, 2008, 31(1): 31-36.
[31] 叶乃兴. 茶叶品质性状的构成与评价[J]. 中国茶叶, 2010, 32(8): 10-11. Ye N X. Composition and evaluation of tea quality characters [J]. China Tea, 2010, 32(8): 10-11.
[32] 郭颖, 陈琦, 黄峻榕, 等. 茶叶滋味与其品质成分的关系[J].茶叶通讯, 2015, 42(3): 13-15, 28. Guo Y, Chen Q, Huang J R. et al. The tea flavor quality and its ingredients [J]. Journal of Tea Communication, 2015, 42(3): 13-15, 28.
[33] 宛晓春. 茶叶生物化学[M]. 3版. 北京: 中国农业出版社, 2003: 217-219. Wan X C. Tea biochemistry [M]. 3rd ed. Beijing: China Agriculture Press, 2003: 217-219.
[34] 王晓萍, 吴洵. 炒青绿茶的生化组分和微量元素综合评级[J]. 茶叶科学, 1991, 11(1): 45-49. Wang X P, Wu X. Comprehensive evaluation of biochemical components and trace elements in roasted green tea [J]. Journal of Tea Science, 1991, 11(1): 45-49.
[35] 郭湘, 唐茜, 许燕, 等. 早春霜冻对不同茶树品种芽叶的生化成分及制茶品质的影响[J]. 云南大学学报(自然科学版), 2015, 37(6): 930-938. Guo X, Tang Q, Xu Y, et al. Effect of frost in early spring on biochemical composition and sensory quality of new shoots in different tea cultivars [J]. Journal of Yunnan University (Natural Sciences Edition), 2015, 37(6): 930-938.
[36] 朱政, 蒋家月, 江昌俊, 等. 低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响[J]. 安徽农业大学学报, 2011, 38(1): 24-26. Zhu Z, Jiang J Y, Jiang C J, et al. Effects of low temperature stress on SOD activity, soluble protein content and soluble sugar content inleaves [J]. Journal of Anhui Agricultural University, 2011, 38(1): 24-26.
[37] 卢克欢, 郭双, 韦悦, 等. 外源性5-氨基乙酰丙酸对盐胁迫下颠茄生理特性及次生代谢产物含量的影响[J]. 西南大学学报(自然科学版), 2018, 40(9): 13-20. Lu K H, Guo S, Wei Y, et al. Effects of exogenous 5-aminolevulinic acid on physiological characteristics and secondary metabolite contents ofL. seedlings under NaCl stress [J]. Journal of Southwest University (Natural Science Edition), 2018, 40(9): 13-20.
[38] 胡云鹏, 康念, 李宇辉, 等. 5-氨基乙酰丙酸对百合生长发育的影响[J]. 仲恺农业工程学院学报, 2017, 30(2): 1-5. Hu Y P, Kang N, Li Y H, et al. Effects of 5-aminolevulinic acid on growth and development of lily [J]. Journal of Zhongkai University of Agriculture and Engineering, 2017, 30(2): 1-5.
[39] 龙志荣, 王登良, 邱瑞瑾, 等. 水解酶对乌龙茶品质形成的影响[J]. 广东茶业, 2007(1): 10-14. Long Z R, Wang D L, Qiu R J, et al. Effect of hydrolase on quality formation of oolong tea [J]. Guangdong Tea Industry, 2007(1): 10-14
[40] 曹慧明, 史作民, 周晓波, 等. 植物对低温环境的响应及其抗寒性研究综述[J]. 中国农业气象, 2010, 31(2): 310-314, 319. Cao H M, Shi Z M, Zhou X B, et al. A review on response of plant to low temperature and its cold resistance [J]. Chinese Journal of Agrometeorology, 2010, 31(2): 310-314, 319.
[41] Ashworth E N, Pearce R S. Extracellular freezing in leaves of freezing-sensitive species [J]. Planta, 2002, 214(5): 798-805.
Effects of Exogenous 5-aminolevulinic Acid on Photosynthetic and PhysiologicalCharacteristics of Tea Plants under Low Temperature Stress
YAN Fei1,2,3,4, JIANG Wenhua1, QU Dong1,2,3, FU Jing1,2,3,4, ZHAO Xuan4
1. College of Biological Science and Engineering, Shaanxi University of Technology, Hanzhong 723000, China; 2. Shaanxi Bio-resources Key Laboratory, Hanzhong 723000, China; 3. Coordination and Innovation Center for Comprehensive Development of Qinba Biological Resources, Hanzhong 723000, China; 4. Qinba Black Tea Research Institute, Shaanxi University of Technology, Hanzhong 723000, China
In order to explore the effects of 5-aminolevulinic acid (ALA) on photosynthetic and physiological characteristics of tea plants under natural low temperature stress in winter, we sprayed 0, 10, 30 mg·L-1and 50 mg·L-1ALA on ‘Shaancha 1’ (low temperature tolerant cultivar) and ‘Jinmudan’ (low temperature sensitive cultivar). The results show that ALA of suitable concentration could improve net photosynthetic rate, stomatal conductance, intercellular CO2concentration, transpiration rate, maximum photochemical efficiency of PSⅡ and potential activity of PSⅡ. Moreover, ALA treatment could promote the accumulations of water extract, caffeine, free amino acids, catechins, theanine, soluble sugar and protein contents. Exogenous ALA could improve the photosynthetic capacity and tea quality under low temperature stress. ALA treatment (50 mgL-1) could effectively improve the cold tolerance of ‘Shaancha 1’. While 10 mg·L-1and 30 mg·L-1ALA treatment had effects on the relief of low temperature stress in ‘Jinmudan’.
ALA, tea plants, low temperature stress, photosynthetic characteristics, physiological characteristics
S571.1
A
1000-369X(2020)05-597-10
2019-11-28
2020-02-13
陕西省教育厅项目(16JS019)、陕西理工大学博士启动基金(SLGQD15-14)、陕南秦巴山区生物资源综合开发协同创新中心项目[QBXT-Z(Z)-15-2]
燕飞,男,讲师,主要从事茶树栽培抗逆生理和分子生物学方面研究,yanfei3111@163.com
猜你喜欢
分子催化(2022年1期)2022-11-02 07:11:08
舰船科学技术(2022年11期)2022-07-15 07:51:56
西藏农业科技(2019年3期)2019-11-04 00:35:10
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
现代食品(2016年24期)2016-04-28 08:12:06
化工进展(2015年3期)2015-11-11 09:07:41
云南中医学院学报(2015年2期)2015-07-31 18:11:59
医学研究杂志(2015年5期)2015-06-10 06:43:26
化学反应工程与工艺(2015年1期)2015-04-16 03:06:16
