
细菌氧化应激反应的研究进展
2020-10-15 08:50:32李纾然谌翰林
化工管理 2020年28期
李纾然 谌翰林
(湖北大学 生命科学学院 生物催化和酶工程国家重点实验室 环境微生物技术中心,湖北 武汉 430062)
0 引言
地衣芽胞杆菌(Bacillus licheniformis)作为一种生物安全性菌株,其发酵产物(纳豆激酶、淀粉酶、聚-γ-谷氨酸、杆菌肽等)常应用于食品类、化妆品类等生物制品的生产中[1]。B. licheniformis属于典型的好氧菌,在其发酵过程中会产生大量的活性氧(reactive oxygen species, ROS),包括超氧阴离子(O2-)、过氧化氢(H2O2)和羟基自由基(·OH),而ROS的存在会对细胞造成巨大损伤,使得DNA断裂、蛋白羰基化、脂质过氧化,最终造成细胞衰老和死亡[1]。
菌体的氧化应激反应受到ROS诱导和生长时期的影响。芽胞杆菌中的转录调控因子PerR和OhrR均会受到ROS的攻击而使得结构发生改变,从而调控多种抗氧化剂的合成以抵抗ROS的氧化伤害[2]。同时,ROS通常在对数中后期大量积累,在稳定期合成最大,不同时期的细胞抵抗氧化应激的能力也不相同,一般稳定期细胞的抗氧化应激能力强于早期细胞[3]。
1 细菌的氧化应激
氧化应激指当细胞受到对自身有害刺激时,产生过多的高活性分子如活性氧自由基(ROS)与活性氮自由基(RNS),氧化物的生成速率大于氧化物的清除速率,导致氧化和抗氧化这两个系统处于失衡状态而造成损伤。真核生物在这一现象的研究比较多,真核细胞可以在氧化应激时通过调节tRNA来调控自身状态[3]。目前对细菌等原核生物的氧化应激调控机制研究相对较少。实验中发现,大部分的细菌受到氧化刺激之后,会通过基因调节的机制来缓解损伤[4-5]。
1.1 活性氧自由基与活性氮自由基
研究发现,ROS包括有超氧阴离子(O2-)、羟自由基(·OH)与过氧化氢(H2O2)等;RNS则是一氧化氮(NO)、二氧化氮(NO2)和过氧化硝酸盐(ONOO-)。而细菌中主要涉及ROS。
ROS中,O2-和H2O2是由呼吸脱氢酶的自氧化作用形成的。在正常周转期间,这些酶使用黄素辅因子从有机底物中接受氢化物阴离子。然后,还原的黄素一次次将电子转移到酶中的次级氧化还原部分上,该次级氧化还原部分是铁硫簇或结合的醌。但是,如果氧气在将电子传递到下一个载流子之前与还原的黄素发生碰撞,则电子会从FADH2跳到氧气上而生[1]。
分子氧本身在π反键轨道上有两个自旋排列的未配对电子(图1)。这种结构使得具有自旋对电子的有机分子不能一次将多个电子转移到氧。因为氧是一种相对较弱的单价电子受体,这种限制确保了氧不能有效地氧化氨基酸和核酸。然而,氧的未成对电子很容易与过渡金属和有机自由基的未成对电子相互作用。
相比而言,氧自由基、过氧化氢和羟基自由基的还原电位决定了它们在热力学上是比双氧强得多的单价氧化剂(图2)。O2-的阴离子电荷抑制了其作为富电子分子氧化剂的有效性,而H2O2的活性则因其氧-氧键的稳定性而降低。但这两个特征都不适用于羟基自由基,·OH与大多数生物分子的反应几乎是扩散受限的。

图1 分子氧、超氧化物、过氧化氢和单线态氧的分子轨道图
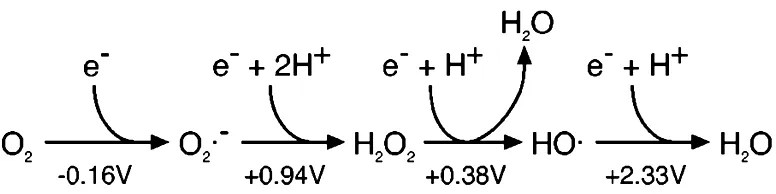
图2 活性氧还原电位(1M双氧水为标准态)
1.2 氧化应激危害
细胞中氧化应激的危害主要存在于三方面:脂类的氧化、DNA的氧化和蛋白质的损伤。
活性自由基直接进攻细胞膜上的不饱和脂肪酸,而导致膜结构、性质的改变以及蛋白质紊乱,产生的醛类则会进一步损伤细胞。
活性基团进攻核苷酸中碱基和糖的部位,导致DNA骨架的断裂;还能引入外来的碱基、糖基,并与其他分子链接来阻止DNA的复制。活性基团还会攻击蛋白质,氧化其氨基酸侧链残基变成硫醇基和羰基,从而导致了蛋白质性质、结构的改变。
2 细菌氧化应激的防御机制
氧化应激的防御机制主要分为两级:一级防御,酶抗氧化系统和非酶抗氧化系统,来使活性基团保持在合理水平;二级防御,用来修复氧化的损伤,如DNA修复、蛋白水解酶等。
最新研究表明大多数细菌中主要是SoxRS、OxyR这两个氧化还原应答转录因子来应对氧化应激[6]。
2.1 SoxRS调控机制
SoxRS的调控子是由两个编码独立转录激活子的基因SoxR、SoxS组成,能表达出SoxR、SoxS两种蛋白。
其中SoxR蛋白是MerR蛋白家族的成员之一,分子量有17 kD。其N端是一个特殊结构能与DNA特异性结合;其C端是4个半胱氨酸残基的序列,可以结合金属配体。
在溶液中以二聚体形式存在,单体中都含有[2Fe-2S]中心,是一个能感知氧化胁迫的分子开关。激活的SoxR能与SoxS启动子牢牢结合,具有高度亲和性。关于结合方式,在对SoxS启动子区的缺失实验结果表明,SoxS启动子-10区到-35区之间的间隔区对SoxR的结合与转录的激活起重要作用。推测可能是正常还原性SoxR结合在SoxS启动子上,RNA聚合酶结合在SoxR-DNA复合体上,此时为闭合结构,无转录激活活性。当氧化胁迫时,[2Fe-2S]中心被氧化,复合体开放,转录激活。
SoxRS调控系统中,调控顺序如下:SoxR由超氧化物刺激后的激活SoxS基因表达;SoxS蛋白含量增多而激活各个被调控基因表达,从而使细胞产生抗氧化。其中被调控基因包括:SodA、AcrAB、fpr等。可以作用于:氧化物清除(超氧化物歧化酶)、膜透性调节及外排、DNA修复(核酸内切酶)[6]。
2.2 脯氨酸代谢增强katG表达和氧化应激抗性
Lu Zhang等[7]研究发现研究表明,除了提供能量外,脯氨酸代谢还影响不同生物体的抗氧化应激能力。通过构建大肠杆菌野生型和putA突变株的应激实验发现putA突变株对氧化应激的敏感性明显高于亲本野生型菌株。用脯氨酸处理野生型细胞可显著提高过氧化氢酶I(由katG编码)的表达量和活性。同时katG菌株对脯氨酸没有反应,说明过氧化氢酶I在脯氨酸保护机制中起关键作用。脯氨酸氧化代谢可产生过氧化氢,这表明脯氨酸通过预适应效应提高大肠杆菌的氧化应激耐受性,包括内源性过氧化氢生成和过氧化氢酶过氧化物酶活性的增强。
2.3 OxyR调控机制
OxyR属于LysR蛋白家族,分子量是34kD。在100~1000μMH2O2的胁迫下,会在分子内形成二硫键,由还原态转为氧化态,从而结合启动子来调控基因。其调控基因有:katG(过氧化酶)、dps(DNA结合蛋白)等来清除细胞内的过氧化氢、保护DNA[6]。
MAYUREE FUANGTHONG,SOPAPAN ATICHARTPONGKUL[11]d等通过对枯草芽孢杆菌中OhrA和OhrB的突变分析发现OhrA和OhrB共同参与了枯草芽孢杆菌的抗氧化应激过程,其中OhrA蛋白在细胞生长中起着更重要的作用,且其表达受到OhrR的抑制。
3 结语
目前对多种调控因子调控地衣芽胞杆菌的抗氧化机制还不清楚,如何清除超氧阴离子(O2-)、过氧化氢(H2O2)与羟自由基(·OH)等活性自由基也还处于研究阶段。将来随着对其研究的深入会进一步阐明各类调控因子对抗氧化应激相关酶的调控机制,以及对半胱氨酸代谢、H2S代谢、Fe代谢等代谢途径的调节。还可以通过代谢组、转录组的研究,挖掘多种全局调控基因的作用,通过加深对地衣芽胞杆菌完整生理过程的认识,可以为以后的工业菌株改造提供依据。
猜你喜欢
中老年保健(2022年3期)2022-11-21 09:40:36
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:05:56
科学与财富(2019年15期)2019-10-21 13:35:48
发酵科技通讯(2018年2期)2018-07-06 11:38:46
湖南林业科技(2017年1期)2017-02-06 05:29:00
西南军医(2016年6期)2016-01-23 02:21:19
发明与创新(2015年37期)2015-02-27 10:40:35
西南军医(2015年2期)2015-01-22 09:09:37
机电信息(2014年5期)2014-02-27 15:51:48
机电信息(2014年2期)2014-02-27 15:51:39
