
人溶菌酶密码子优化及其在牛乳腺细胞中高效表达
2020-10-15 08:51田媛王力龙凤昝林森成功
中国农业科学 2020年18期
田媛,王力,龙凤,昝林森,2,成功,2
(1西北农林科技大学动物科技学院,陕西杨凌 712100;2国家肉牛改良中心,陕西杨凌 712100)
0 引言
【研究意义】抗生素滥用所引发的细菌耐药性、抗生素残留及食品安全等问题已被视为全球危害公众健康的问题,人们迫切需要开发新型广谱抗菌剂。溶菌酶又称胞壁质酶或N-乙酰胞壁质聚糖水解酶,在唾液、泪液、血清、人乳、牛乳和禽类蛋清中广泛分布,是一种能水解致病菌中黏多糖的碱性酶,具有广谱抗菌、消炎、抗病毒及免疫调节等多种功效,在畜牧、农产品、食品和医疗等行业广泛应用[1-3]。人溶菌酶和鸡蛋清溶菌酶氨基酸序列具有59%同源性,但抗菌活性远高于目前使用较广的鸡蛋清溶菌酶,具有更广阔的应用前景[4]。然而,通过传统方法从母乳、胎盘、唾液中提取分离或微生物表达系统生产重组人溶菌酶,获得量很少,稳定性低,无法满足研究和潜在市场应用的需求[5]。基因工程不断发展为大规模高效生产重组人溶菌酶提供了可能。因此,进一步优化表达系统,通过生物反应器实现重组人溶菌酶高效表达,将为今后重组人溶菌酶的工业化生产和应用奠定基础。【前人研究进展】2006年,MEGA首次利用山羊乳腺生物反应器生产了重组人溶菌酶[6],随后在牛[7-9]、猪[10]、山羊[11]等动物的研究中均有成功实现了重组人溶菌酶在乳腺中表达的报道。然而,不容忽视的是,异源重组蛋白表达量低是目前生物反应器中亟待解决的难题。研究发现,宿主细胞密码子使用偏好很大程度上影响了异源重组蛋白在宿主细胞的高效表达[12]。组成蛋白质的氨基酸有 20种,而编码氨基酸的密码子有64种(简并密码子)。不同生物,甚至同种生物不同的蛋白质编码基因,对简并密码子使用频率并不相同,具有一定的偏爱性,其中使用频率高密码子称为最佳密码子,而那些很少被利用的密码子称为稀有或低频密码子。研究发现,少量的 DNA密码子的改变,在很大程度上影响了蛋白翻译速率,即使替换单个简并密码子,其蛋白合成的速度会减缓到其正常速度的1/10甚至更慢[13],密码子使用偏好与蛋白翻译效率呈显著正相关[14]。通过蛋白质和mRNA表达数据库联合分析发现,密码子的选择决定了蛋白的表达量,生物体通过密码子的选择进而调控内源蛋白的表达丰度[15]。密码子的选择在生物反应器研究过程中十分重要,外源基因含有宿主细胞基因使用的低频密码子或者偏好密码子不一致情况下,往往会影响外源基因的mRNA的稳定性和翻译效率,造成重组蛋白表达量低[16]。前人研究结果比较发现,转基因山羊、牛乳腺中重组人乳铁蛋白表达量为 1.5—30 g·L-1,而重组人溶菌酶的表达量只有0.025—0.27 g·L-1[17],表达量低造成提纯成本高,难以达到乳腺生物反应器工业化生产要求。【本研究切入点】因此,人溶菌酶基因密码子使用偏好性是否与牛主要乳蛋白密码子使用偏好一致?这种差异是否是导致人溶菌酶在乳腺生物反应器中表达量低的主要原因?密码子优化能否进一步提高重组人溶菌酶表达量?这些问题仍待进一步研究。【拟解决的关键问题】本试验通过对牛乳腺主要乳蛋白密码子使用偏好性进行分析,并根据牛乳蛋白高频密码子和低频密码子对人溶菌酶基因密码子进行翻译起始区优化和全局优化设计,通过荧光素酶、实时荧光定量PCR及Western-blot等方法从mRNA和蛋白水平对密码子优化效果进行分析比较,实现人溶菌酶基因的高效表达,为开展重组人溶菌酶的工业化生产及应用奠定基础。
1 材料与方法
试验于 2019年在西北农林科技大学国家肉牛改良中心实验室完成。
1.1 材料
PGL3-Basic荧光素酶载体、海肾荧光素酶载体为实验室保存。E.coliDH 5a菌株和pMD19-T easy载体购自大连Takara公司。牛成纤维细胞通过耳组织块法分离获得,实验室冻存。牛乳腺上皮细胞系购自通派(上海)生物科技有限公司,小鼠乳腺上皮细胞系C127购自北京协和细胞库。主要试剂包括:高保真PCR酶、限制性内切酶、PrimeScript™ RT reagent Kit with gDNA Eraser反转录试剂盒和SYBR Premix EX Taq定量PCR试剂盒购自宝生物工程(大连)有限公司;T4 DNA连接酶、Dual-Glo双荧光素酶检测试剂盒购自普洛麦格(北京)生物技术有限公司;Endo-free Plasmid Kit、胶回收试剂盒购自Omega Biotek股份有限公司;Lipo3000转染试剂及 opti-MEM 购自Invitrogen公司;Western-blot细胞裂解液、BCA蛋白浓度测定试剂盒及SDS-PAGE蛋白上样缓冲液购自上海碧云天生物技术有限公司;蛋白酶抑制剂购自罗氏公司。兔源溶菌酶多抗、兔源GAPDH单抗和HRP标记的羊抗兔二抗分别购自Santa Cruz(sc-292849),Abcam(ab181603)和生工生物工程(上海)股份有限公司(D110058)。
1.2 牛主要乳蛋白基因和人溶菌酶基因密码子使用偏好分析
选取 αs1、αs2、β、κ-酪蛋白,β-乳球蛋白,α-乳清白蛋白,乳铁蛋白等7种牛乳中主要乳蛋白基因CDS序列,用于密码子使用偏好性分析。具体乳蛋白基因名称、GenBank登录号、CDS序列长度见表1。
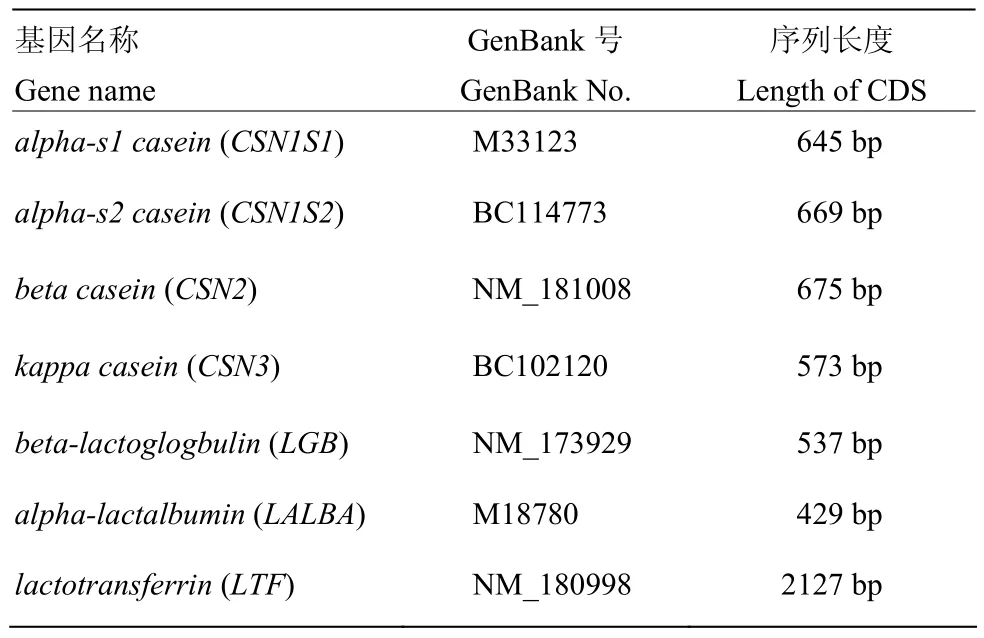
表1 7种牛乳蛋白基因GenBank登录号及序列信息Table 1 GenBank accession number and sequence information of cattle seven milk protein genes
利用 CodonW1.42和 EMBOSS(http://imed.med.ucm.es/EMBOSS/)中CUSP、CHIPs等工具对牛主要乳蛋白基因密码子及人溶菌酶密码子使用特性进行分析。分析的特性参数主要包括:A3s,G3s,C3s,同义密码子在第 3 位上相应碱基的出现频率(T3s)、基因的G+C含量(GC)、密码子第3位的G+C 含量(GC3s)、密码子适应性指数(CAI)、有效密码子数(ENc)等。同义密码子的相对使用度(RSCU)是对同义密码子的使用偏好性评估,该值等于同义密码子的实际观测值与同义密码子平均使用期望值的比值。如果密码子使用无偏好性,则RSCU值为1;如该密码子比其他同义密码子使用更频繁,则其 RSCU值大于1,反之亦然。将7个牛乳蛋白基因和人溶菌酶基因各自作为一个对象,以密码子的RSCU 值作为变量,对应每个编码氨基酸密码子利用TBtools 0.665绘制热图并进行聚类分析。
1.3 溶菌酶密码子优化及基因克隆
根据牛主要乳蛋白基因密码子使用偏好性,对人溶菌酶基因局部(翻译起始区前22位密码子)和全局优化设计。利用mfold在线软件对优化后的人溶菌酶基因序列进行RNA二级结构分析(http://unafold.rna.albany.edu/)。
采用Trizol 法提取人Hela细胞总RNA, 通过凝胶电泳和核酸定量检测 RNA质量和浓度。根据Takara反转录试剂盒操作说明对RNA进行反转录获得cDNA。以cDNA为模板,利用P-pGL3-cw引物(表 2)克隆获得人溶菌酶编码区序列(野生型溶菌酶序列,LYZcw)。反应条件为:98 ℃,30 s模板变性;98℃,5 s;57℃,15 s;72℃,15 s,30个循环。72℃,5 min延伸。以LYZcw cDNA为模板,利用P-pGL3-op22引物(表2)克隆获得翻译起始区密码子优化序列(LYZop22),PCR反应条件同上。对上述获得的LYZcw、LYZop22PCR产物经过胶回收纯化、序列末端加A反应、T-A克隆并DH5α转化后进行测序验证(LYZcw-T、LYZop22-T)。全局密码子优化的溶菌酶序列(LYZop),经生工生物工程(上海)股份有限公司全基因合成并克隆到 T载体中(LYZop-T)。

表2 LYZcw、LYZop22、LYZop序列克隆引物信息及基因合成Table 2 Primers for the amplication of LYZcw, LYZop22 and LYZop
1.4 pGL3-LYZcw/op22/op溶菌酶-荧光素酶融合表达构建
以Invitrogen公司pGL3-control载体为骨架载体,构建人溶菌酶基因与萤火虫荧光素酶基因融合表达载体。通过NcoI内切酶分别酶切LYZcw-T、LYZop22-T、LYZop- T和pGL3-control载体。1%琼脂糖凝胶电泳回收LYZcw、LYZop22、LYZop溶菌酶基因序列和线性化的pGL3-control载体。通过T4 DNA连接酶进行连接,分别构建pGL3-LYZcw/op22/op荧光素酶融合表达载体。测序筛选鉴定LYZcw/op22/op序列以正向插入并与萤火虫荧光素酶基因正确融合的质粒用于下一步研究。
1.5 细胞转染及荧光素酶检测密码子优化效果
分别将实验室冻存的牛乳腺上皮细胞系(BMEC)、牛成纤维细胞(BFFC)及C127小鼠乳腺上皮细胞系复苏,用完全培养基(DMEM高糖+10% FBS +1% PS)重悬,接种到T75培养瓶中,置于37 ℃,5% CO2培养箱中培养。待细胞汇合度达到 90%时进行传代接种于24孔培养板中(2×105/孔)。接种后的第二天,按照每孔 0.8 μg质粒(PGL-LYZcw/LYZop/LYZop22∶pRL-tk = 10∶1):2 μLlipo3000比例转染上述细胞,每组4个复孔,转染48 h后通过荧光素酶活性检测比较人溶菌酶密码子优化效果。
1.6 pcDNA3.1-LYZcw/op22/op溶菌酶过表达载体构建
分别以pGL3-LYZcw/op22/op质粒为模板,利用P-pcDNA-cw、P-pcDNA-op22、P-pcDNA-op引物(表2)克隆LYZcw、LYZop22、LYZop序列(PCR反应条件参照1.3)并连接到T载体。使用HindIII、XhoI内切酶分别酶切pcDNA3.1(+)、LYZcw-T、LYZop22-T vector、LYZop- T,连接构建 pcDNA-LYZcw/op22/op人溶菌酶过表达载体,测序做进一步鉴定。
1.7 细胞转染及实时荧光定量 PCR检测密码子优化效果
将培养的牛成纤维细胞(BFFC)、牛乳腺上皮细胞(BMEC)及C127细胞传代接种于12孔培养板中(2×105/孔)。接种后的第二天,按照lipo3000转染试剂说明书,每孔 1.6 μg质粒(pcDNA 空载、pcDNA-LYZcw/op22/op):4 μL lipo3000 比例转染上述细胞,每组4个复孔。24 h后按照1.3中方法提取细胞总 RNA并反转录获得 cDNA,用于下一步定量PCR检测。
对不同密码子优化方式对重组人溶菌酶 mRNA水平表达量进行定量分析。反应条件为:预变性95 ℃,30 s。95 ℃,5 s;60 ℃,34 s,40 个循环。循环结束后进行熔解曲线分析,确保扩增条带单一。
1.8 Western-blot检测密码子优化效果
将培养的牛乳腺细胞(BMEC)经胰酶消化后接种于6孔培养板中(4×105/ 孔)。接种后的第二天,按照 lipo3000转染试剂说明书,每孔 4.0 μg质粒(pcDNA 空载、pcDNA-LYZcw/op22/op):10 μL lipo3000比例转染上述细胞,每组4个复孔。转染48 h后,收集蛋白用于Western-blot检测。蛋白电泳采用分离胶浓度为12%,蛋白上样量40 μg。电泳条件为浓缩胶80 V,分离胶120 V,待溴酚蓝跑出胶后终止电泳。蛋白转膜按照伯乐半干转膜仪操作说明,200 mA, 1h进行转膜。转膜结束后,PVDF膜经5%脱脂奶粉封闭——一抗4 ℃孵育过夜(人溶菌酶、GAPDH 1∶1 000稀释)——TBST洗涤(3次,每次10 min)——二抗室温孵育2 h(1∶3 000稀释)——TBST洗涤(3次,每次10 min)——显影——ChemiDoc XRS+仪器成像。
1.9 数据分析
采用2-△△Ct方法对real-time qPCR 分析计算目的基因相对表达量。GraphPad Prism 6.0进行 One-way ANOVA分析和作图。数据均以平均数±标准误(Mean±SEM)表示。
2 结果
2.1 牛乳蛋白及人溶菌酶基因密码子组成分析
通过CodonW软件对牛乳蛋白及人溶菌酶基因密码子组成进行分析,结果表明,牛乳蛋白基因密码子 GC含量平均为 0.485 ± 0.026,第三位碱基GC3s含量平均为0.537 ± 0.062。人溶菌酶基因密码子GC含量为0.473,第三位碱基GC3s含量为0.407。上述结果表明,人溶菌酶基因整体 GC含量与牛乳蛋白基因GC含量相近,但第三位碱基GC含量低于牛乳蛋白基因 GC含量。牛乳蛋白基因密码子偏好以 GC结尾,而人溶菌酶基因密码子偏好以 AT结尾(表3)。
ENc值范围为20(每个氨基酸只使用一个密码子)到61(各个密码子被均衡使用),其值越低,偏好性越强。牛乳蛋白及人溶菌酶ENc值分别为49.814和50.27,表明牛乳蛋白和人溶菌酶基因密码子使用相对较平均。ENC vs GC3s线性回归分析结果表明,7种乳蛋白基因有效密码子数 ENc值与第三位碱基GC3s呈显著负相关(R= - 0.8968,P<0.01)(图1),表明密码子使用偏好性越强(ENc值越小)的基因,其GC3s值越高,主要偏好以GC碱基结尾的密码子。因此,牛乳蛋白基因密码子的使用偏好性受GC3s影响。
2.2 牛乳蛋白基因密码子使用偏好性分析
对编码氨基酸且为简并密码子的7种牛乳蛋白基因使用的共59个密码子(不含Met、Trp及3个终止密码子)进行同义密码子的相对使用度(RSCU)分析。结果表明,牛乳蛋白基因对CTG(2.71)、AGG(2.32)、GCC(1.73)、GTG(1.68)、ATC(1.64)等5个密码子具有明显的偏好性(RSCU>1.5),为高频密码子。而 TTA(0.12)、ATA(0.18)、TCG(0.19)、CTA(0.25)、CCG(0.26)、GCG(0.45)、ACG(0.48)等7个密码子在牛乳蛋白基因中使用较小(RSCU<0.5),为低频密码子(表4)。

表4 牛乳蛋白基因密码子使用偏好性分析Table 4 Codon preference analysis of bovine milk protein genes

图1 牛乳蛋白基因ENc vs GC3s相关性分析Fig. 1 Correlation analysis of ENc vs GC3s of bovine milk protein genes
2.3 基因密码子偏好性的聚类分析
以密码子的RSCU 值作为变量,对具有简并性的59个编码氨基酸的密码子进行聚类分析。依据RSCU值可以将7种牛乳蛋白较好的分成为两大类:酪蛋白类(CSN2、CSN1S2、CSN3和CSN1S1)和乳清蛋白类(LTF、LGB和 LALBA),而人溶菌酶基因密码子使用偏好与牛乳酪蛋白类更接近(图 2)。结果表明,即使在同一乳腺组织内,编码牛乳酪蛋白类基因密码子和乳清蛋白类基因密码子在使用偏好性上可能也存在一定差异。
2.4 人溶菌酶基因密码子优化及分析
根据表3结果中牛乳蛋白基因高频密码子及低频密码子使用情况,对人溶菌酶密码子进行了优化。其中LYZop对溶菌酶全局密码子进行了优化,而 LYZop22只对翻译起始区前22位密码子进行了优化(表5)。

表5 人溶菌酶基因密码子优化情况Table 5 Codon Optimization of human lysozyme gene

图2 牛乳蛋白及人溶菌酶基因密码子使用RSCU值聚类分析Fig. 2 Clustering analysis of bovine milk protein and human lysozyme genes based on RSCU value
CodonW比较优化前后序列,结果表明,LYZcw、LYZop22、LYZop第三位碱基即 GC3s含量分别为0.407、0.450、0.757。mRNA二级结构最小自由能MFE从-138.83 kcal/mol(LYZcw)分别降低到了-141.02 kcal/mol(LYZop22)和-178.12 kcal/mol(LYZop),mRNA二级结构稳定性明显提高。对mRNA翻译起始区100个碱基进行二级结构分析,发现相比于LYZcw(-29.7 kcal/mol),LYZop22(-26.4 kcal/mol)和 LYZop(-31.0 kcal/mol)密码子优化并没有明显改变翻译起始区二级结构最小自由能,相反在5′端引入了更多的茎环结构(图3),有利于核糖体与mRNA的结合并启动翻译。

图3 人溶菌酶密码子优化翻译起始区(1-100 bp)mRNA二级结构示意图Fig. 3 Schematic diagram of RNA second structure of codon optimized human lysozyme gene (1-100bp)
2.5 人溶菌酶-荧光素融合载体构建及溶菌酶表达分析
电泳和测序结果表明,成功克隆到了人溶菌酶野生型LYZcw、翻译起始区密码子优化的LYZop22 CDS区序列及全基因合成的全局密码子优化的 LYZop CDS区序列,且与预期序列信息一致(图4)。Nco1酶切和测序结果表明,成功构建了 pGL3-LYZcw/op22/op 3个人溶菌酶-荧光素酶融合表达载体,且LYZcw、LYZop22、LYZop编码区与萤火虫荧光素酶基因正确融合,没有移码突变(图5)。
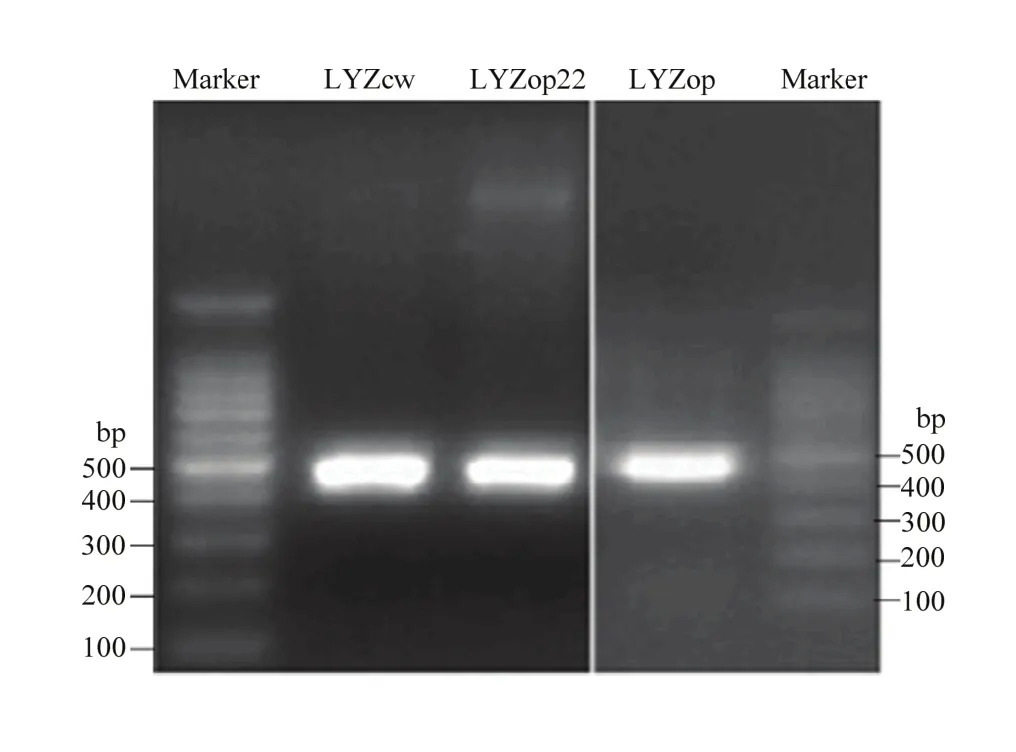
图4 LYZcw、LYZop22和LYZop电泳检测Fig. 4 LYZcw, LYZop22 and LYZop electrophoresis detection
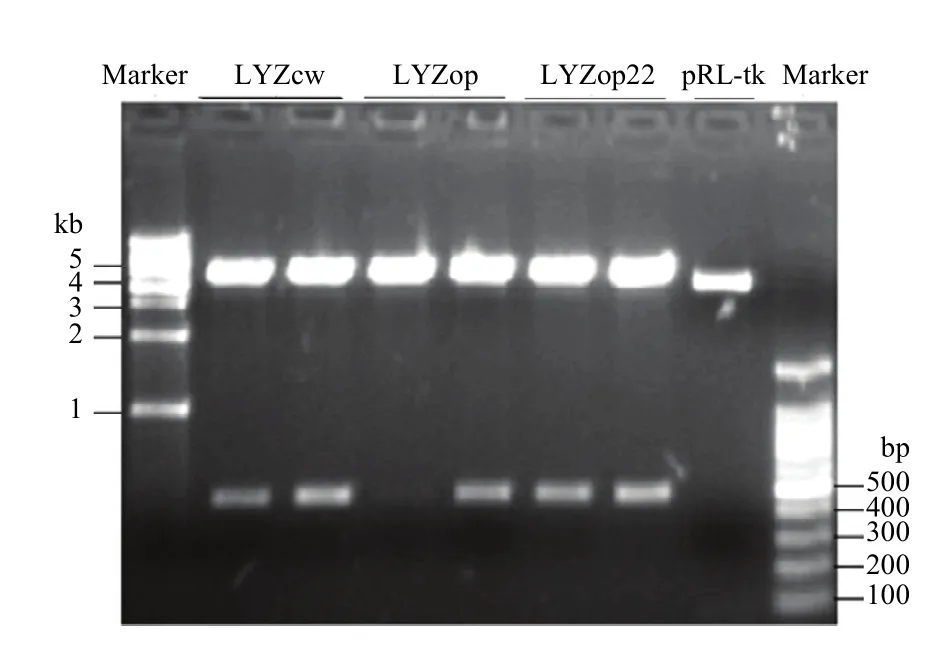
图5 pGL3-LYZcw/op22/op载体Nco1酶切鉴定Fig. 5 pGL3-LYZcw/op22/op vector identified by Nco1 enzyme digestion
通过lipo3000 将pGL-LYZcw+pRL-tk、pGL-LYZop+ pRL-tk、pGL-LYZop22 + pRL-tk分别转染BMEC、BFFC、C127细胞,48 h后检测荧光素酶表达量。荧光素酶检测结果表明,相比LYZcw,全局密码子优化的LYZop在BMEC和对照BFFC、C127细胞中分别提高了2.20倍(P<0.01)、2.44倍(P<0.01)和1.99倍(P<0.01)。翻译起始区密码子优化的 LYZop22在BMEC细胞中相比野生型提高了1.48倍(P<0.01),而在对照BFFC、C127细胞中差异不显著(P>0.05)(图6)。
2.6 mRNA、蛋白水平检测人溶菌酶基因密码子优化效果
构建 pcDNA-LYZcw/op22/op过表达载体,并分别转染BMEC、BFFC和C127细胞,检测密码子优化对人溶菌酶mRNA表达量的影响。实时荧光定量PCR结果表明,LYZop22相比LYZcw在BMEC和BFFC细胞中分别提高了2.08倍(P<0.05)和1.5倍(P>0.05)。而LYZop相比野生型LYZcw在上述两个细胞中分别提高了17.8倍(P<0.01)和22倍(P<0.01)(图 7)。上述结果表明,根据牛乳腺主要蛋白进行密码子优化后的溶菌酶能显著提高其在牛乳腺上皮细胞和成纤维细胞转录水平表达量。
进一步对 pcDNA-LYZcw/op22/op过表达载体转染的牛BMEC细胞进行人溶菌酶蛋白表达检测。相比野生型LYZcw,密码子优化的LYZop、LYZop22明显提高了人溶菌酶在牛BMEC细胞中的表达量,且全局密码子优化的 LYZop溶菌酶表达量高于翻译起始区密码子优化的LYZop22(图8-A)。上述结果表明,密码子优化能明显提高人溶菌酶在牛乳腺上皮细胞中的表达。然而,经全局密码子优化的人溶菌酶基因在牛乳腺上皮细胞中蛋白表达水平提高倍数低于mRNA水平提高倍数(图8-B)。
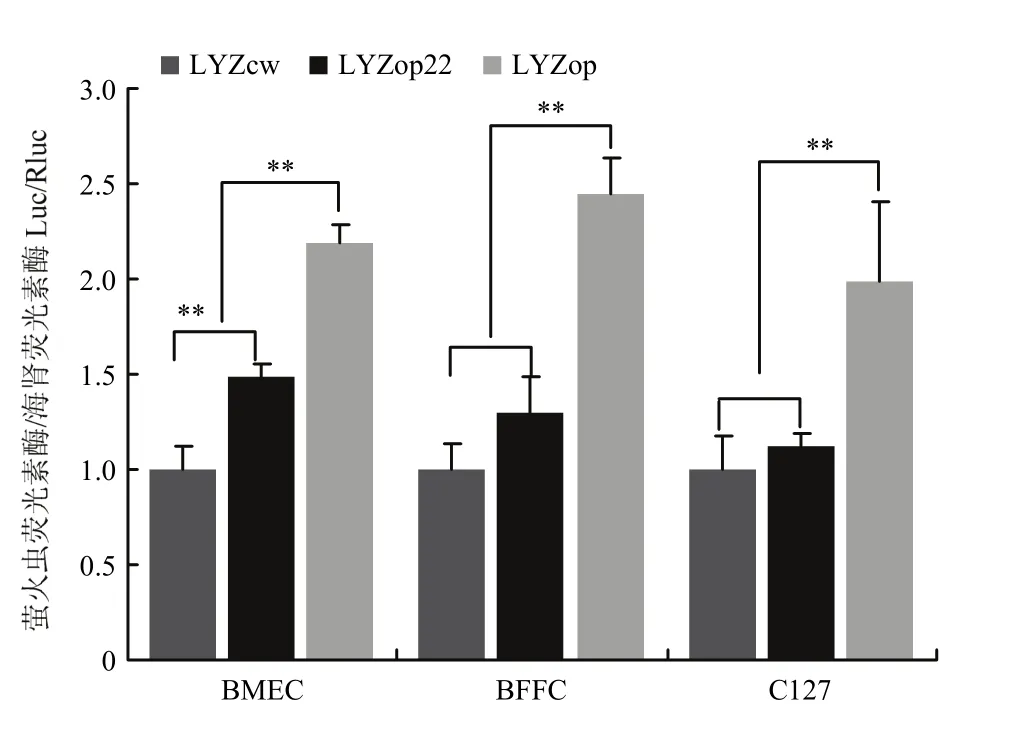
图6 荧光素酶检测人溶菌酶基因密码子优化效果Fig. 6 The effect of codon optimization analysis of human lysozyme gene by luciferase

图7 mRNA水平检测人溶菌酶基因密码子优化效果Fig. 7 The effect of codon optimization analysis on human mRNA level of human lysozyme gene
3 讨论
人溶菌酶作为非特异性免疫因子,对金黄色葡萄球菌、链球菌、大肠杆菌等均具有广谱的抗菌作用,在体内发挥着杀菌消炎、免疫调节、改善胃肠道菌群的作用。同时,在畜牧、农产品、食品、医疗和化妆品轻工等行业也具有广泛应用前景。利用现代基因工程和乳腺生物反应器生产重组人溶菌酶克服了传统提取方法蛋白稳定性差、成本高的不足,但如何进一步提高重组蛋白的表达量并用于下一步工业化生产仍是乳腺生物反应器首要考虑的问题。

图8 蛋白水平检测密码子优化对人溶菌酶表达量的影响Fig. 8 The effect of codon optimization analysis on protein level of human lysozyme
近年来研究发现,不同生物密码子的选择不仅对蛋白高水平表达发挥作用,同时对微量的调控蛋白表达也起到调节作用[13],这一发现对于提高外源重组蛋白的表达和生产具有重要的指导意义。研究发现,同一物种,不同组织甚至同一组织内不同蛋白其密码子使用偏好性均存在差异,正是这种差异可能进一步调节着蛋白的表达量[18]。本研究中根据牛乳中7种主要乳蛋白基因密码子偏好性进行聚类分析发现,牛乳中酪蛋白类(CSN2、CSN1S2、CSN3和CSN1S1)和乳清蛋白类(LTF、LGB和 LALBA)可以明显的分为两类。表明,同一组织内牛乳酪蛋白和乳清蛋白基因密码子使用偏好存在一定的差异,而这种密码子使用偏好性可能在一定程度上影响着牛乳酪蛋白和乳清蛋白表达量。密码子使用偏好性分析发现,牛乳蛋白基因中发现5个高频密码子和7个低频密码子,且牛乳蛋白基因密码子第三位碱基偏好以GC结尾,而人溶菌酶偏好以AT结尾。而较高的GC含量,特别是简并碱基GC含量在哺乳动物细胞中与mRNA丰度和蛋白表达量呈正相关[19-20]。表明,人溶菌酶密码子使用偏好性不利于在牛乳中高效表达,这也可能是造成牛乳腺生物反应器中重组人溶菌酶表达量低的主要原因[17, 21]。
密码子使用偏好性在调节mRNA稳定性、蛋白翻译起始、延伸等方面发挥了重要的作用[22-23]。本研究中局部和全局密码子优化后的人溶菌酶 mRNA二级结构自由能 MFE分别由-138.83 kcal/mol降低到了-141.02 kcal/mol和-178.12 kcal/mol。前人研究发现,mRNA折叠强度与 mRNA丰度和蛋白表达量呈正相关[24-25]。本研究发现,随着密码子优化后人溶菌酶mRNA二级结构稳定性的增加,其mRNA表达量也相应提高。在牛BMEC等3种细胞中,全局密码子优化的人溶菌酶 mRNA水平显著高于局部密码子优化的人溶菌酶,这一结果也与优化后 mRNA 稳定性呈正相关。表明,密码子优化可能通过增加mRNA二级结构稳定性和 mRNA转录效率进而提高了人溶菌酶mRNA表达水平。
研究发现,mRNA 二级结构5′端的茎环结构有利于蛋白翻译起始和延伸,进而提高蛋白表达量[26-27]。密码子优化后的溶菌酶尽管整体mRNA MFE明显降低,但进一步对 5′端翻译起始后 100 bp mRNA碱基进行二级结构分析发现,密码子优化并没有明显改变人溶菌酶基因前100bp 二级结构自由能,反而在翻译起始区引入了更多的茎环结构,而5′端茎环结构的引入,有利于提高蛋白翻译起始效率[28]。酿酒酵母中研究发现,蛋白表达丰度与 mRNA折叠强度存在正相关,而有效的 mRNA翻译起始效率和较高的 mRNA二级结构缩短了核糖体之间的距离,使 mRNA的翻译效率提高,进而提高了蛋白的表达丰度[25,29]。本研究中也发现,翻译区密码子优化后牛 BMEC和 BFFC细胞中人溶菌酶基因mRNA水平和蛋白水平相比野生型都有一定程度的提高。全局密码子优化的人溶菌酶基因,在mRNA翻译起始区引入了更多的茎环结构,同时,mRNA整体折叠也更稳定。因此,密码子优化后的人溶菌酶表达量在 mRNA 水平和蛋白水平相比野生型均得到明显的提高。
密码子优化是提高外源重组蛋白表达的重要途径之一。尽管目前已开发许多软件可根据宿主密码子使用偏好对目的基因密码子进行优化和设计,然而关于密码子优化的“金标准”仍未统一,密码子优化的效果也不尽如人意,归根结底是由于目前关于密码子优化是如何影响外源蛋白表达仍存在很大的争议。密码子优化在mRNA转录效率[30]、稳定性[31],蛋白翻译起始[32]、延伸效率、蛋白折叠及蛋白稳定性[33]等多个环节均发挥了调节作用。ZHOU等研究发现,密码子优化提高了目的基因蛋白和RNA表达水平,但主要是体现在mRNA转录水平的提高[30]。PRESNYAK 等[31]认为,密码子优化是决定 mRNA稳定性的主要因素。本研究中发现,经全局密码子优化的人溶菌酶基因mRNA二级结构稳定性得到明显提高,其在牛乳腺上皮细胞中转录水平提高了17.8倍,然而其蛋白水平提高倍数却低于mRNA水平提高倍数。表明,密码子优化大幅提高了重组人溶菌酶mRNA的表达水平,但mRNA表达水平与蛋白表达水平并不一定呈强相关[34],而mRNA转录后的修饰可能进一步影响了蛋白的表达。因此,通过密码子优化提高mRNA表达水平的同时如何进一步提高蛋白表达水平及其内在分子机制仍有待于进一步研究。
4 结论
本研究通过生物信息学分析了牛乳腺主要乳蛋白基因密码子使用偏好,获得了牛乳蛋白基因密码子使用偏好及使用的高低频密码子;依据牛乳蛋白基因密码子使用偏好性对人溶菌酶全局密码子优化能显著提高重组人溶菌酶mRNA水平和蛋白水平的表达量,为今后利用生物反应器高效生产重组人溶菌酶奠定基础。
猜你喜欢
畜牧兽医学报(2022年4期)2022-04-24
食品安全导刊(2021年21期)2021-08-30
福建农业学报(2021年6期)2021-08-18
消费导刊(2020年35期)2021-01-28
大连工业大学学报(2020年3期)2020-08-13
发明与创新·中学生(2019年6期)2019-06-26
食品界(2019年2期)2019-03-10
生物学教学(2018年2期)2018-08-07
安徽农业科学(2018年1期)2018-05-14
科技视界(2018年2期)2018-03-27
