
水稻产量性状一般配合力QTL定位
2020-10-09 08:34曹应江游书梅蒋开锋杨乾华郑家奎
核农学报 2020年9期
曹应江 游书梅 蒋开锋 张 涛 杨 莉 杨乾华 秦 俭 郑家奎
(四川省农业科学院水稻高粱研究所/国家水稻改良中心泸州分中心/农业部西南水稻生物学与遗传育种重点实验室,四川 德阳 618000)
水稻是最重要的粮食作物之一,世界上一半以上的人口以稻米作为主食[1]。在我国,随着现代工业化和城市化的发展,可种植水稻耕地面积逐渐减少,因此通过提高水稻单位面积产量,保证稻谷总产量,是我国育种家普遍采取的主要技术途径。我国从1973年实现水稻三系配套成功,1976年开始大面积推广种植杂交稻以来,水稻单产大幅度提高[2],有效缓减了耕地减少与稻米需求增长之间的矛盾。高配合力亲本的创制,是培育高产杂交稻新品种的必要条件。因此,定位水稻产量性状高配合力数量性状座位(quantitative trait locus,QTL),对水稻高产育种具有重要意义。
配合力是指一个亲本(纯系、自交系或品种)材料在由它所产生的杂种一代的产量或其他性状表现中所起作用相对大小的能力。配合力分为一般配合力和特殊配合力。一般配合力(general combining ability,GCA)是指一个被测自交系与其他自交系组配的一系列杂交组合产量或其他数量性状的平均表现,由基因的加性效应决定,是可遗传的部分,因此GCA 的高低取决于自交系所含有利基因位点的数量,自交系所含有利位点越多,其GCA 越高。特殊配合力(specific combining ability,SCA)是指2 个亲本(系)所组配的杂交种产量或其他数量性状的水平。不完全双列杂交(NCⅡ)设计是配合力分析最经典的遗传试验设计。NCⅡ设计是指把供试亲本按试验要求分为2 组,只进行组间杂交,不进行组内杂交。当一组亲本数为n,另一组亲本为m 时,共有m×n 个组合。广大育种家已将这种设计应用于不同作物多种性状的配合力研究[3-8]。
GCA 作为水稻亲本最核心的性状之一,除了采用传统的广泛杂交方法筛选高配合力亲本外,研究者还主要开展了以下研究:一是优异配合力的简单重复序列标记(simple sequence repeats,SSR)鉴定或关联标记筛选研究[9-10];二是把配合力看作一个性状,用数量性状QTL 定位的方法开展研究。Qu 等[11]利用珍汕97与9311 的重组自交系群体,与两系不育系杂交构建双列杂交群体,定位水稻相关性状的一般配合力QTL;Xiang 等[12]定位了22 个主要产量性状一般配合力QTLs;Qi 等[13]开展了玉米一般配合力QTL 定位研究。此外,付新民[14]和张庆路[15]利用中国香稻与明恢63构建的重组自交系群体,开展了水稻一般配合力QTL定位研究。本研究利用泸恢8258 与扬恢34 构建的重组自交系(recombinant inbred lines,RIL)群体,分别与3 种不同胞质类型的不育系泸98A(K 型)、Ⅱ-32A(印尼水田谷型)、冈46A(冈型)杂交,构建双列杂交群体、定位水稻产量及相关性状一般配合力的QTL,以期为水稻产量性状高配合力基因的分子标记辅助选择,精细定位或克隆提供参考依据。
1 材料与方法
1.1 试验材料
泸恢8258 是四川省农业科学院水稻高粱研究所选育的水稻高配合力恢复系,是我国应用面积较大的水稻恢复系之一。目前,以泸恢8258 为亲本共组配了8 个品种,11 次通过省级或国家审定(表1)。本研究选用泸恢8258 与扬恢34 杂交,构建了包含193 个家系的重组自交系群体,按照NCⅡ设计的方法,分别与3 种不同类型胞质的不育系泸98A(K 型)、Ⅱ-32A(印尼水田谷型)、冈46A(冈型)杂交,包含双亲的杂交组合,共获得585 份杂交种子作为试验材料。
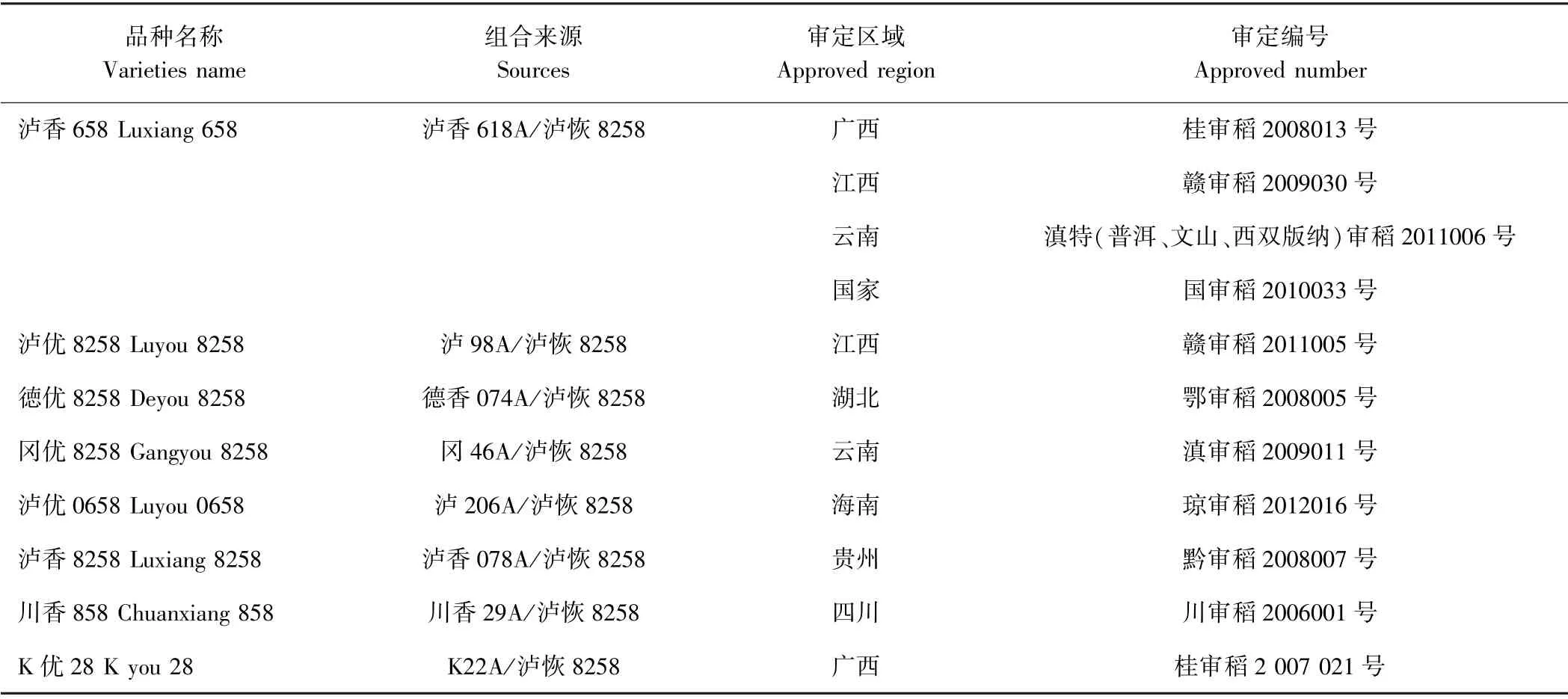
表1 以泸恢8258 为亲本之一组配的8 个杂交稻品种Table1 Eight hybrid rice varieties using Luhui 8258 as one of the parents
1.2 田间试验
田间试验在同一年份的3 种环境下进行,分别是四川省农业科学院水稻高粱研究所德阳基地(德阳,Deyang)、泸州基地(泸州,Luzhou)以及四川省遂宁市大英县象山镇(遂宁,Suining)。根据3 个试验点的不同气候环境,结合当地的大田生产,德阳基地3月25日播种,5月1日移栽;泸州基地3月10日播种,4月12日移栽;遂宁4月5日播种,5月10日移栽。每份材料种植3 行,每行6 株,行株距统一为33.3 cm×19.8 cm。第1 次重复按编号顺序栽插,第2、第3 次重复时每重复内随机排列。田间肥水管理同当地大田生产,及时防治病虫害。整个试验期间,未遭遇影响水稻生长的极端异常天气。
1.3 水稻性状调查与数据分析
成熟时,行区内去除边际效应,选取中间生长整齐一致的3 株水稻考种,包括单株生物量、收获指数、单株产量、有效穗数、每穗颖花数、每穗实粒数、结实率和千粒重8 个性状。泸州点未考察单株生物量和收获指数。
利用DPS 7.05 软件分析配合力及相关遗传参数,配合力分析参照文献[16-17]的方法。
1.4 连锁图谱构建及QTL 定位
连锁遗传图谱应用Mapmaker 3.0 构建,利用Kosambi 函数将重组率转化为遗传距离(cM)[18],并应用MapDraw V2.1 绘制遗传连锁图谱[19]。首先利用Temnykh 等[20]报道的524 个SSR 标记筛选到112 个标记在双亲间有多态性,然后结合群体扩增情况,去掉12 个图像不清晰和极度偏分离的标记后共有100 个标记,最后利用这100 个标记构建初步的遗传图谱。针对初步遗传图谱中遗传距离较远的区间,查询水稻基因组数据库(http:/ /www.gramene.org)公布的水稻基因组数据,重新设计了363 个SSR 标记、95 个插入缺失标记(insertion-deletion,InDel)和80 个序列标签位点标记(sequence-tagged site,STS),并对双亲进行多态性检测,筛选到84 个标记进入图谱。最终遗传图谱包含184 个标记,每条染色体的标记数在12 ~22 个之间,平均每条染色体标记数为15.33 个。InDel 和STS 标记名称及序列详见表2。所有的标记均由上海英骏生物技术有限公司合成。采用Sodium dodecyl sulfate(SDS)法提取DNA。

表2 图谱中用到的InDel 和STS 引物序列Table2 Primer sequences of InDel and STS used in this study
利用标记加密后的遗传图谱,结合表型农艺性状统计值,采用Win QTL Cartographer 2.5 软件进行QTL分析[21],应用复合区间作图法(composite interval mapping,CIM),以似然函数比值的对数(logarithm of odds,LOD)值≥2.5 作为QTL 存在阈值,检测QTL,估算QTL 的加性效应和可解释表型变异量。采用McCouch[22]的命名系统对定位到的QTL 进行命名。
2 结果与分析
2.1 水稻各性状方差分析及一般配合力方差分析
在德阳、遂宁和泸州3 种环境下,大多数性状的遗传型、恢复系、不育系、恢复系×不育系的F值均达到显著或极显著差异水平(表3),说明各处理间存在显著的遗传差异。但也有少数性状在个别环境下未达到显著差异水平,如收获指数在德阳和遂宁环境下的恢复系×不育系间等。多数性状在区组间未达到显著差异水平,说明试验整体比较均匀,但仍然有少数性状在不同环境的区组间达到极显著差异水平,这种现象可能是土壤肥力差异或其他因素造成的。

表3 水稻8 个农艺性状在3 种环境下的配合力的方差分析(F 值)Table3 Variance analysis of the combining ability of eight yield-associated traits under three environments(F value)
2.2 水稻各性状基因型方差、贡献率和遗传力
由表4可知,水稻各性状在3 种环境下一般配合力方差占比(Vg)均大于50.00%(德阳环境中单株产量,泸州环境单株产量和结实率除外),说明本试验中F1的表现主要受到亲本间基因加性效应的作用。VgR占总方差的比率在8.79%~84.27%之间,每穗颖花数在德阳、遂宁和泸州3 种环境下VgR 占Vg比均大于50.00%,说明该性状在3 种环境下恢复系发挥了主要作用。单株生物量、有效穗数、每穗实粒数等3 个性状在德阳和遂宁环境下VgR 占比均大于50%,说明这3个性状在德阳和遂宁环境下恢复系起了主要作用。不育系一般配合力方差占比在1.86%~59.92%之间,德阳环境中千粒重占比大于50.00%;遂宁环境中结实率占比大于50.00%;泸州环境中千粒重占比大于50.00%,表明这些性状在各自的环境中不育系发挥了主要作用。
由表4可知,各性状广义的遗传力在13.55%~84.77%之间,狭义的遗传力在10.62%~79.23%之间。千粒重在3 种环境下广义和狭义的遗传力均高于70.00%,说明该性状遗传给子代的能力很强,不易受环境变化的影响,而其他性状在3 种环境下的遗传力均较小,说明这些性状容易受环境的影响。
2.3 水稻各性状一般配合力的相关性
由表5可知,各性状在德阳、遂宁和泸州3 种环境下一般配合力相关系数在-0.68 ~0.96 之间,其中部分性状一般配合力相关性均达到显著或极显著差异水平,如有效穗数、每穗实粒数、结实率与单株产量,每穗颖花数、每穗实粒数与有效穗数,每穗实粒数、千粒重与每穗颖花数,结实率、千粒重与每穗实粒数。此外,部分性状间的相关性只在1 种环境下达到显著或极显著水平,如单株产量与千粒重仅在遂宁环境下达到显著相关;部分性状无相关性,如收获指数与千粒重,每穗颖花数与结实率等。上述结果说明水稻各性状的一般配合力间具有复杂的关系。

表4 水稻8 个性状的基因型方差、配合力方差的贡献率和遗传力Table4 Genotypic variance,contribution ratio of combining ability variance and heritability for eight yield-associated traits
2.4 一般配合力在水稻重组自交系中的分布
根据各性状一般配合力效应的大小及变异幅度,将193 个株系分为15 个组。由图1可知,水稻各性状在德阳、遂宁和泸州不同环境下表现为正态分布或偏态分布,有明显的峰值,表明各性状一般配合力效应在水稻重组自交系群体中呈连续分布,具有多基因控制数量性状的显著特征,适合进行QTL 分析。
2.5 水稻各性状一般配合力的QTL 定位
由图2和表6可知,德阳、遂宁和泸州3 种环境下共检测到50 个一般配合力QTL,分布在除第5 号染色体外的其他11 条染色体上。单个QTL 贡献率变幅在3.26%~34.26%之间,LOD 值变幅为2.52~16.55。检测到QTL 数量最多的性状是每穗颖花数和结实率,为8 个;最少的性状是收获指数,为2 个。27 个QTL 增效等位基因来自泸恢8258,占检测到QTL 总数的54.00%,除每穗颖花数和每穗实粒数一般配合力QTL多数增效等位基因来自扬恢34 外,其他性状的QTL增效等位基因大多来自泸恢8258。在所有QTL 中,有5 个QTL 在德阳、遂宁和泸州3 种环境中均被检测到,分别是控制有效穗数的qEP2-2,位于第2 染色体的RM526~RM599 区间,对表型的贡献率分别为8.94%、10.46%和16.53%,增效等位基因来自泸恢8258;控制每穗颖花数的qSP2-2,对表型的贡献率分别为27.83%、14.54%和34.26%,增效等位基因来自扬恢34;控制每穗实粒数的qFGP2-2,对表型的贡献率分别为24.00%、10.30%和17.75%,增效等位基因来自扬恢34;控制千粒重的qTGW1 和qTGW2,对表型的贡献率为5.98%、6.48%、18.66%和29.01%、24.84%、12.05%,增效等位基因来自泸恢8258。此外,还有qHI3、qEP7、qSP7、qSSR12-1、qTGW3-2 5 个QTL 在其中2 种环境下检测到。其他的QTL 仅在其中一种环境下检测到,具体分析如下:
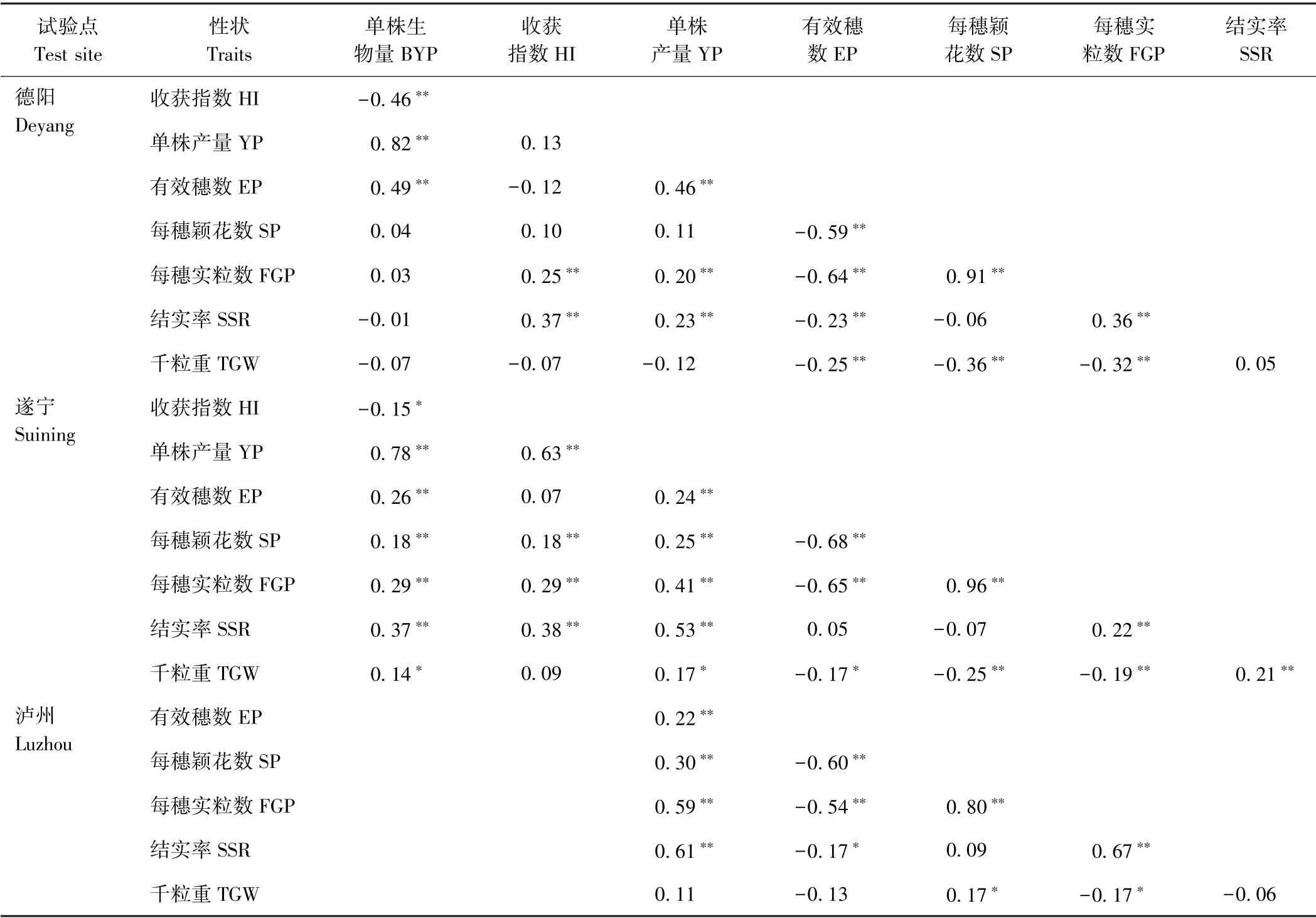
表5 水稻8 个产量性状一般配合力的相关性Table5 Correlation coefficients between eight rice yield-associated traits of general combining ability
1)单株生物量。在德阳和遂宁环境下检测到6个控制单株生物量的QTL,分布在第2、第3、第8、第9和第12 条染色体,单个QTL 对表型的贡献率在4.49%~9.84%之间,qBYP2-1 和qBYP12 QTL 增效等位基因来自泸恢8258,其他QTL 增效等位基因来自扬恢34。
2)收获指数。共检测到2 个控制收获指数的QTL,分布在第1 和第3 条染色体,单个QTL 对表型的贡献率在7.60%~17.42%之间。qHI3 在2 种环境下均被检测到,对表型的贡献率为17.42%和10.86%,增效等位基因来自泸恢8258。
3)单株产量。德阳、遂宁和泸州3 种环境下共检测到5 个单株产量一般配合力QTL,分布在第2、第3、第4、第6 和第9 条染色体,单个QTL 对表型的贡献率在5.67%~9.28%之间,5 个QTL 增效等位基因全部来自泸恢8258。
4)有效穗数。共有7 个有效穗数一般配合力QTL被检测到,分布在第2、第3、第7、第9 和第10 条染色体,单个QTL 对表型的贡献率在5.58%~19.20%之间。qEP2-2 在3 种环境下均被检测到,对表型的贡献率分别为8.94%、10.46%和16.53%,增效等位基因来自泸恢8258;qEP7 在德阳和遂宁环境下检测到,对表型的贡献率为5.58%和9.29%,增效等位基因来自扬恢34。此外,还有qEP2-1、qEP2-3、qEP9 和qEP10 4个QTL 增效等位基因来自泸恢8258。
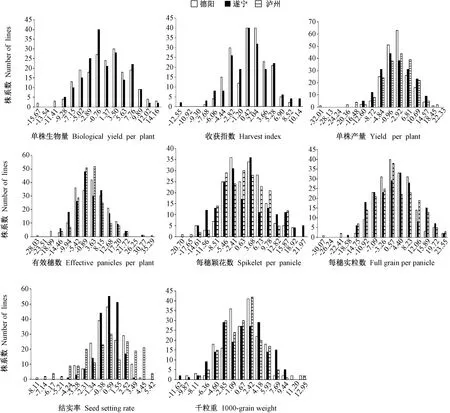
图1 8 个产量相关性状一般配合力在水稻重组自交系群体中的分布Fig.1 Distribution of general combining ability on eight yield-associated traits of rice in RIL population
5)每穗颖花数。德阳、遂宁和泸州3 种环境下共检测到8 个控制每穗颖花数一般配合力QTL,分布在第2、第7、第8、第9、第10 和第11 条染色体,单个QTL 对表型的贡献率为3.89%~34.26%之间。qSP2-2 在3 种环境下均被检测到,对表型的贡献率为27.83%、14.54%和34.26%,增效等位基因来自扬恢34。qSP7 在德阳和遂宁环境下检测到,对表型的贡献率为8.73%和5.35%,增效等位基因来自泸恢8258。qSP7、qSP8 和qSP11 3 个QTL 增效等位基因来自泸恢8258,其他QTL 增效等位基因来自扬恢34。
6)每穗实粒数。共检测到7 个控制每穗实粒数一般配合力的QTL,分布在第2、第3、第7、第9、第10条染色体,单个QTL 对表型的贡献率在4.05% ~24.00%之间。qFGP2-2 在德阳、遂宁和泸州下均被检测到,对表型的贡献率为24.00%、10.30%和17.75%,增效等位基因来自扬恢34。qFGP3、qFGP7 和qFGP10 3 个QTL 增效等位基因来自泸恢8258,其他QTL 增效等位基因来自扬恢34。
7)结实率。共有8 个结实率一般配合力QTL 被检测到。单个QTL 对表型的贡献率在4.25% ~20.64%之间,qSSR12-1 在德阳和遂宁2 种环境下同时被检测到,对表型的贡献率为7.92%和10.93%,增效等位基因来自泸恢8258。其他还有qSSR3-1、qSSR3-2 和qSSR9 3 个QTL 增效等位基因来自泸恢8258,其他QTL 增效等位基因来自扬恢34。
8)千粒重。在德阳、遂宁和泸州3 种环境下共检测到7 个控制千粒重一般配合力QTL,单个QTL 对表型的贡献率在3.26%~29.01%之间。有2 个QTL 在3种环境下均检测到,分别是qTGW1(表型贡献率为5.98%、6.48%和18.66%)、qTGW2(表型贡献率为29.01%、24.84%和12.05%),增效等位基因均来自泸恢8258。qTGW3-2 在德阳和遂宁均被检测到,对表型的贡献率为3.26%和5.88%,增效等位基因来自泸恢8258。此外,还有qTGW3-1 增效等位基因来自泸恢8258,其他QTL 增效等位基因来自扬恢34。
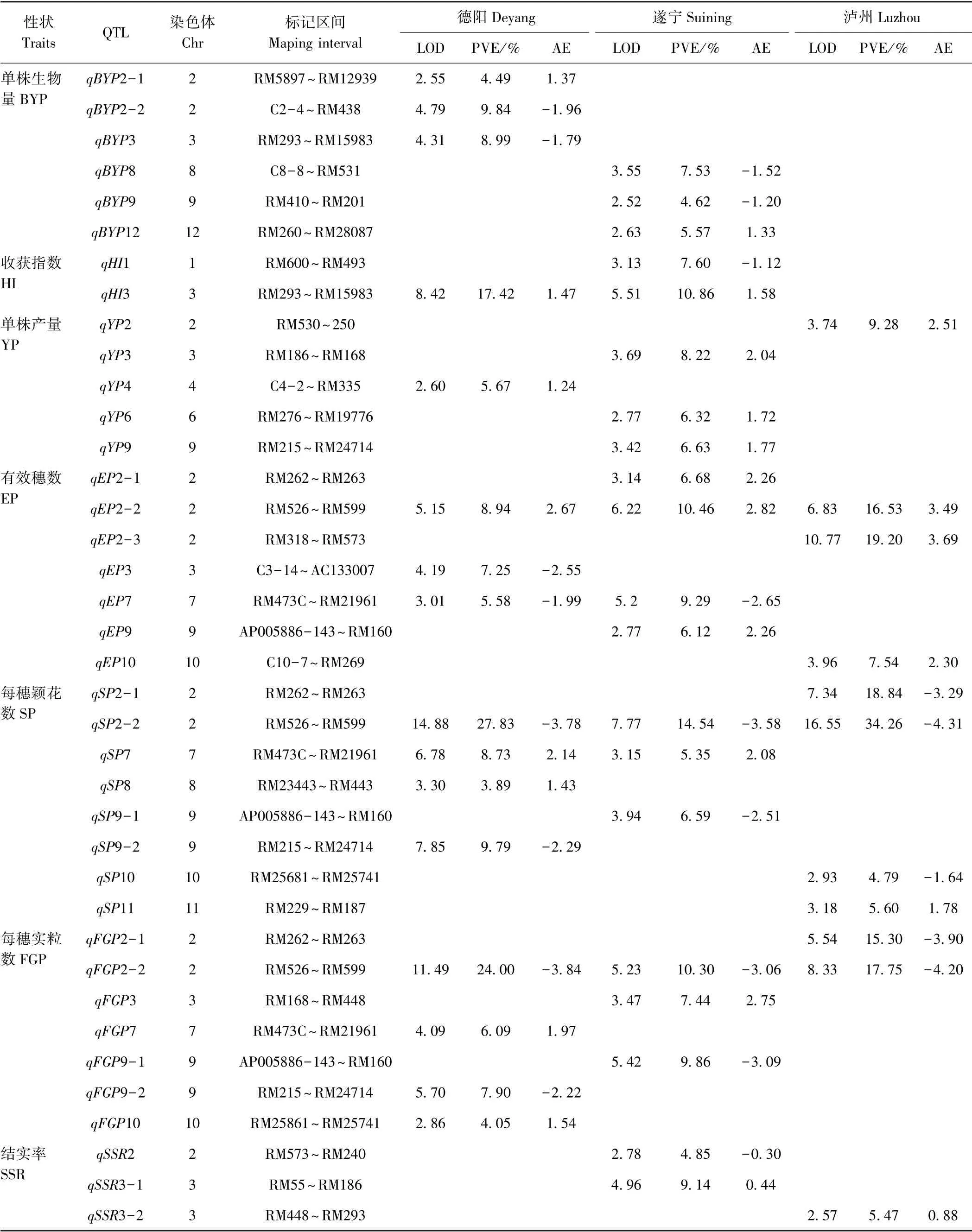
表6 德阳、遂宁和泸州3 种环境下检测到的各产量相关性状的QTLTable6 QTL for GCA of yield-associated traits identified in 3 environments such as Deyang,Suining and Luzhou
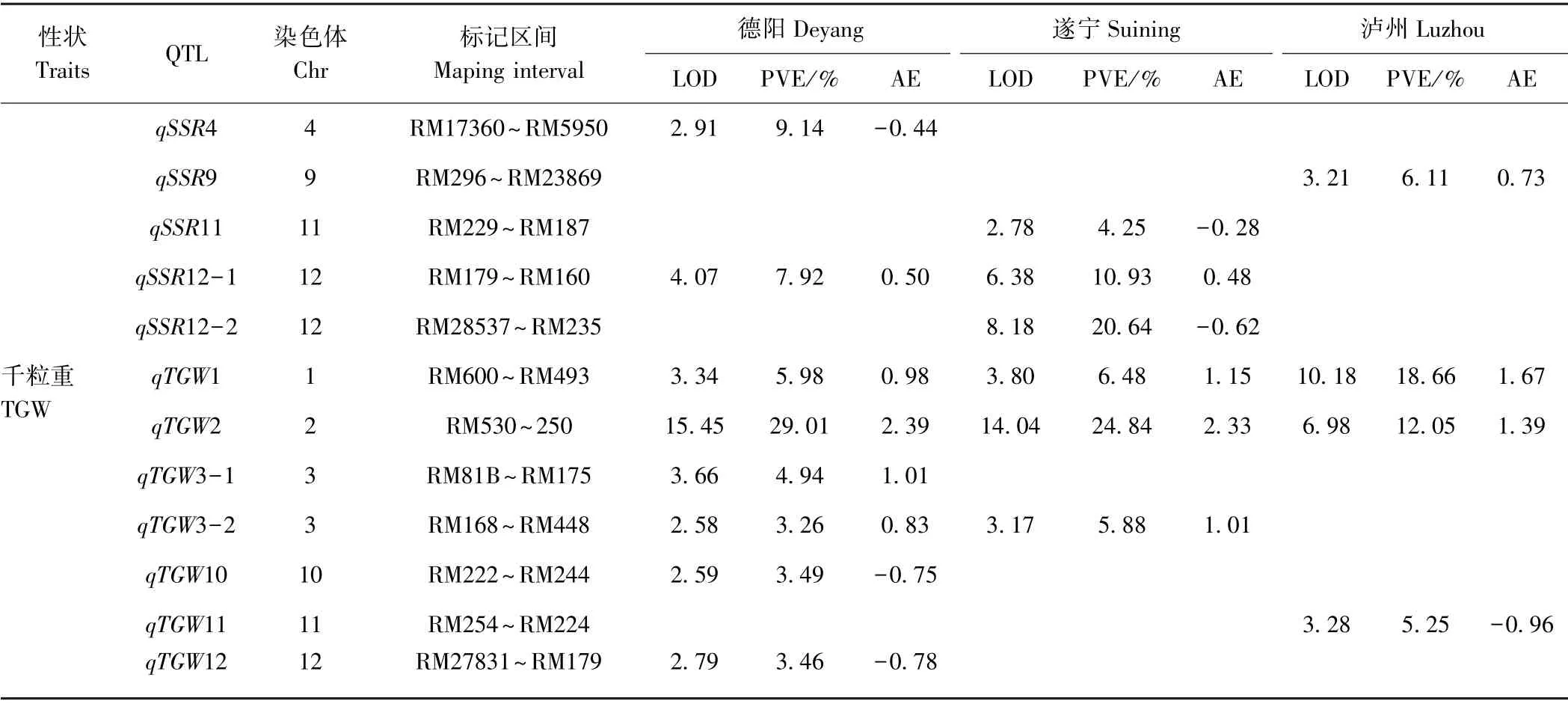
表6 (续)

图2 水稻各产量相关性状一般配合力QTL 在染色体上的分布Fig.2 Distribution of QTLs for rice yield associated traits of GCA on chromosomes
3 讨论
选育高配合力亲本材料是培育优异杂交品种的基础,然而在实际的育种实践中,育种专家通常是利用稳定的品系与不同的材料广泛的杂交,通过杂交F1组合表现的好坏,评价该品系配合力的高低,这种方法工作量大,耗时长。随着科学技术的进步,越来越多的性状已从原来单一表型的选择,转变为更加简便且有效的分子标记辅助选择,如水稻抗稻瘟病[23-24]、水稻部分品质基因[25]等。然而对于水稻高配合力基因的定位鲜有研究,目前高配合力基因分子标记辅助选择成功的报道较少。本研究共检测到50 个产量或相关性状高配合力的QTL,为产量性状一般配合的分子标记辅助选择提供了理论基础。
水稻产量性状的遗传不仅受到本身基因位点的影响,而且易受环境、遗传世代背景等因素的影响[26-28],因此,同一性状在不同的作图群体中检测到的基因也可能不同,同一群体在不同环境或不同年份检测到的基因也可能不同,本研究共检测到50 个产量相关性状一般配合力的QTL,但仅有qEP2-2、qSP2-2、qFGP2-2、qTGW1 和qTGW2 在德阳、遂宁和泸州3 种环境下被检测到,qHI3、qEP7、qSP7、qSSR12-1 和qTGW3-2 在其中2 种环境下被检测到,其他的QTL 仅在其中1 种环境下检测到,表明水稻一般配合力QTL 在不同环境下表达的差异化。
根据已有报道,科学家们对水稻QTL 进行了大量的研究,许多的性状检测到了大量的QTL,如产量性状、品质性状、农艺性状等,甚至有部分QTL 已经被克隆,如GS5[29]、Ghd7[30]等。但水稻产量性状一般配合力QTL 的报道相对较少,更没有QTL 被克隆。本研究共检测到50 个产量性状一般配合力QTL,与前人检测到的QTL 比较,多为不同位点,仅有控制单株产量一般配合力的qYYP4 与张庆路[15]检测到的qYD-4 在相同区间。本研究中检测到的其他一般配合力QTL 鲜见相关报道。就单一性状的QTL 与本研究检测到配合力QTL 有重叠的现象,如每穗实粒数的qFGP3 与沈希宏等[31]报道的qFGP3.2 和冯跃等[32]报道的qFGP3.1 位于相近区间。由此可知,虽然部分单一性状QTL 与配合力QTL 的区间有重叠,但其间是否存在必然关系,还需进一步深入研究。
4 结论
本研究共检测到50 个产量性状一般配合力QTL,单个QTL 对表型的贡献率变幅在3.36%~34.26%之间,有27 个增效等位基因来自泸恢8258,有5 个QTL在德阳、遂宁和泸州3 种环境下均被检测到,有5 个QTL 在其中2 种环境下检测到,其他QTL 在1 种环境下检测到。本研究结果为进一步开展相关基因的精细定位、克隆和分子标记辅助育种奠定了基础。
猜你喜欢
养殖与饲料(2021年11期)2021-11-15
园艺与种苗(2021年8期)2021-09-23
现代临床医学(2021年4期)2021-07-31
科教新报(2021年21期)2021-07-21
现代临床医学(2021年1期)2021-01-26
廉政瞭望(2020年14期)2020-11-17
廉政瞭望(2020年14期)2020-08-12
小说界(2016年3期)2016-06-06
园艺与种苗(2015年8期)2015-12-28
商界(2000年7期)2000-06-14
