
贵州特有植物黔中杜鹃种群结构及其动态分析
2020-09-28 08:15姜顺邦袁丛军余德会戴晓勇龙海燕
浙江林业科技 2020年4期
姜顺邦 ,袁丛军,余德会,戴晓勇,杨 冰,李 鹤,龙海燕
(1.贵州省林业学校,贵州 贵阳 550201;2.贵州省林业科学研究院,贵州 贵阳 550005;3.贵州雷公山国家级自然保护区管理局,贵州 雷山 557100)
种群是物种进化的基本单位,种群结构是种群最基本的特征,它既可反映种群内不同大小个体数量的分布和配置状况,也能反映种群的数量动态及其未来发展与演变趋势[1]。种群生命表与其构建的存活曲线以及生存分析理论不仅可以反映种群目前的存活状态,而且可以反映植物与环境之间的适合度[2],结合种群动态量化分析和时间序列预测还可以阐明种群的受干扰状况及未来的发展趋势。因此,研究植物种群结构与动态有助于了解种群的生活史和物种的更新动态,对野生植物的保护及开发利用具有重要意义。
黔中杜鹃Rhododendron feddei为杜鹃花科Ericaceae 杜鹃花属Rhododendron常绿灌木或小乔木,生长于海拔1 120~ 1 740 m 的丛林中,为贵州特有种[3],该种虽然发现较早,但在《贵州植物志》和《中国植物志》中都没有收录,而是在《FOC》中才有记载[4]。经过多年的寻找,我们在其模式产地的贵定县云雾山一带再次发现该种,在雷公山保护区也发现了该种,有关报道在贵州百里杜鹃(百里杜鹃国家级森林公园)发现的该种经过查证后并不存在[4]。由于杜鹃花科植物具有很高的利用价值和科研价值,近年来,关于杜鹃花科植物种群结构的研究比较多,如莫罗坚对东莞莲花山毛棉杜鹃花Rh.moulmainense种群结构进行了动态分析,认为毛棉杜鹃花种群径级分布呈倒“J”型,属于增长型种群[5];邓贤兰对井冈山杜鹃Rh.jinggangshanicum种群与群落特征进行了研究,认为井冈山杜鹃种群径级结构为金字塔型,生命期望值基本随径级增加而呈递减趋势,存活曲线为近直线型,种群属于增长型种群[6];蔡鑫对华顶杜鹃Rh.huadingense的种群结构和种间联结进行了研究,认为华顶杜鹃种群呈衰退趋势,幼苗和幼树储备不足[7]。这些学者的研究在很大程度上揭示了许多杜鹃花科植物的种群结构及其数量动态变化特征。黔中杜鹃作为贵州特有植物,种群数量少,具有一定的研究价值,然而,目前有关该物种的研究尚未见报道。本文以雷公山国家级自然保护区内仙女塘的黔中杜鹃种群为研究对象,对其种群结构及数量动态变化进行了分析,揭示该区黔中杜鹃种群数量变化动态特征、种群动态变化趋势,以期为该种的保护和进一步研究提供重要理论支撑。
1 材料与方法
1.1 研究区概况
研究区位于贵州雷公山国家级保护区仙女塘,地理坐标为108°11′37.98″ E,26°22′18.87″ N,属于中亚热带季风气候,年均温在13.9~ 15.2℃,极端最低气温为-6.2℃,最高气温为21.7℃,≥10℃年积温在4 098.0~ 4 531.2℃,年日照时数为1 513.2 h,无霜期在281~ 290 d,年均降水量在1 323.7~ 1 577.2 mm,雨期4-9 月降水量在1 065.1~ 1 249.7 mm,占全年降水的80.0%。
调查群落位置均在中坡,土壤类型主要为黄壤,群落为常绿落叶阔叶混交林,其中,乔木层树种主要有白辛树Pterostyrax psilophyllus,檫木Sassafras tzumu,枫香树Liquidambar formosana,云南桤叶树Clethra cavaleriei,花楸树Sorbus pohuashanensis,交让木Daphniphyllum macropodum,老鼠矢Symplocos stellaris,毛棉杜鹃花,青冈Cyclobalanopsis glauca,树参Dendropanax dentiger,水青冈Fagus longipetiolata,甜槠Castanopsis eyrei,银木荷Schima argentea,云贵鹅耳枥Carpinus pubescens,锥栗Castanea henryi等;灌木层植物有川桂Cinnamomum wilsonii,川杨桐Adinandra bockiana,吊钟花Enkianthus quinqueflorus,光叶山矾Symplocos sumuntia,黄丹木姜子Litsea elongata,凯里杜鹃Rh.kailiense,柃木Eurya japonica,毛果杜鹃Rh.seniavinii,毛棉杜鹃花,石木姜子Litsea elongatavar.faberi,茶荚蒾Viburnum setigerum,狭叶卫矛Euonymus stenophyllus,紫金牛Ardisia japonica等;草本植物分布较少。
1.2 研究方法
1.2.1 样地设置与调查方法 在野外实地踏查的基础上,选取黔中杜鹃天然群落为研究对象,于2019 年5 月进行调查,分别设置具代表性的样地3 个(表1),统一设在在中坡位置,每个样地大小为20 m×30 m,总样地面积为1 800 m2,样地间的距离大于100 m,每个样地以10 m 为间隔设置成6 个10 m×10 m 的样方,共18 个样方。详细记录每个样地的海拔、坡向、坡度等立地因子;对样地内乔木树种进行每木调查,记录各树种的株高、地径(胸径)、冠幅、枝下高等指标;同时,在每个样方的左下角设置1 个2 m×2 m 样方进行灌木层调查、1 个1 m×1 m 样方进行草本层调查。

表1 群落样地基本情况Table 1 Basic information of sample plots
1.2.2 径级划分 为避免破坏黔中杜鹃野生植物资源,研究采用“空间替代时间”的方法,即将林木依地径大小分级,以立木径级结构代替年龄结构分析种群动态,“Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ,Ⅵ,Ⅶ,Ⅷ径级”分别对应“1,2,3,4,5,6,7,8 龄级”。关于种群大小结构立木径级的划分标准,许多学者提出了不同的标准[8-12]。根据该种的生物学特性,本试验以灌丛为单株,采用植株地径(BD)作为个体大小的指标研究其种群大小结构。径级划分方法:Ⅰ级(BD<2.0 cm),Ⅱ级(2.0≤BD<4.0 cm),Ⅲ级(4.0≤BD<6.0 cm),Ⅳ级(6.0≤BD<8.0 cm),Ⅴ级(8.0≤BD<10.0 cm),Ⅵ级(10.0≤BD<12.0 cm),Ⅶ级(12.0≤BD<14.0 cm),Ⅷ级(14.0≤BD<16.0 cm)。
采用陈晓德[13]的数学模型推导种群年龄结构的动态指数对黔中杜鹃种群动态进行定量描述,其计算方法为:

式中,Vn为种群从n级到n+1 级的个体数量变化指数;Vpi为整个种群结构的数量变化动态指数;Sn为第n个龄级种群的个体数;Sn+1为第n+1 个龄级种群的个体数。当考虑未来的外部干扰时:

式中,K为种群龄级数量。Vpi与Vn的数值取正、负、零的意义分别反映了种群或相邻龄级个体数量的增长、衰退及稳定的动态关系。
1.2.3 生命表编制 静态生命表又称特定时间生命表,主要用于木本植物种群的统计研究,其主要参数如下:a0为种群开始时的个体数量,Ax为在x龄级内的现有个体数;ax为匀滑后x龄级内的现存个体数;lnlx为在x龄级开始时的标准化存活个体数;dx为从x到x+1 龄级间隔期内的标准化死亡数;qx为从x到x+1 龄级间隔期间的死亡率;lx为从x到x+1 龄级间隔期间还存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望或平均期望寿命;Kx为消失率(损失度)。表中各项都是相互关联的,可以通过实测值求得,其关系为:

由于静态生命表是反映了多个世代重叠的年龄动态历程中的一个特定时间,而不是对这一种群的全部生活史的追踪,并且调查中存在系统误差,在生命表中会出现死亡率为负的情况,因此,本研究采用匀滑技术对数据进行处理[14]。
1.2.4 生存函数 采用生存率函数(Si)、累积死亡率函数(Fi)、死亡密度函数(fi)和危险率函数(λi)[15-18]等指标对黔中杜鹃自然种群的生存规律及其种群动态进行分析,其计算公式如下:

式中,hi是第i间隔期的时间长度,pi为存活率,qi为死亡频率。
1.2.5 种群数量动态的时间序列预测 本研究采用时间序列分析中的一次移动平均法[19-20]对黔中杜鹃种群的年龄结构进行预测,其计算公式如下:

式中,n表示需要预测的时间(本研究为龄级时间),t为龄级,Xk为k龄级内的个体数量,Mt表示经过未来n个龄级时间后t龄级的种群大小。本文对未来经过2,4,6,8 和10 龄级时间后的种群各龄级的个体数量进行预测。
2 结果与分析
2.1 黔中杜鹃所在群落树种组成
调查结果表明,在研究区域的样地中,共有维管束植物23 科36 属43 种,其中,蕨类植物有2 科2 属2 种,被子植物有21 科34 属41 种。群落结构主要分为乔木层、灌木层和草本层三个层次。构成乔木层的树种共17种,其物种组成及重要值见表2;构成灌木层的树种共22 种,其物种组成及重要值见表3;黔中杜鹃在乔木层中的重要值为0.298,在灌木层中的重要值为0.099,虽然黔中杜鹃在乔木层和灌木层中的重要值均为最大,但其他物种的重要值占比也相对较大,所以,黔中杜鹃群落为混交林。草本层种类及数量均较少,其种类、频度等见表4。

表2 黔中杜鹃群落乔木层种类组成及重要值Table 2 Composition of species and importance values at tree layer in sample plots

表3 黔中杜鹃群落灌木层种类组成及重要值Table 3 Composition of species and importance values at shrub layer in sample plots

表4 黔中杜鹃群落草本层种类组成及重要值Table 4 Composition of species and importance values at herb layer in sample plots
2.2 黔中杜鹃种群分布现状及结构特征
黔中杜鹃种群结构呈现出不规则金字塔形(图1),种群个体数量主要集中分布在Ⅲ~Ⅴ径级,这三个径级的个体数量占种群总数量的69.23%,说明该黔中杜鹃种群的中幼龄个体数很充足,但幼龄个体数较少,仅为种群总数量的11.54%,严重缺乏幼龄个体,中老龄个体数量总体呈现出下降趋势。
采用种群动态量化方法对黔中杜鹃种群相邻大小级的结构动态变化进行分析,以便对其结构特征进行更准确的评价。根据黔中杜鹃种群结构(图1)计算得出,种群从Ⅰ级至Ⅷ级各径级间的Vn分别为-80.00%,-37.50%,-20%,-41.18%,58.82%,71.43%,50%;Vpi为14.92%,考虑种群外部干扰,Vpi为0.67%。由此可见,虽然黔中杜鹃种群结构存在较大波动变化过程,但目前仍处于增长型。
2.3 黔中杜鹃静态生命特征
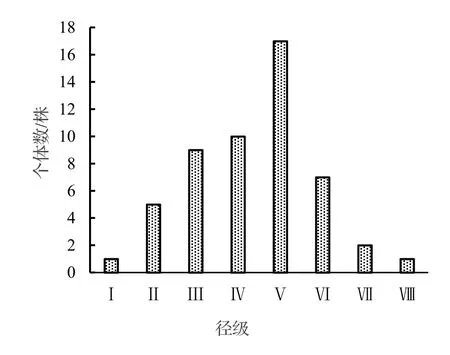
图1 黔中杜鹃种群径级结构Figure 1 Diameter class structure of Rh.feddei population
静态生命表不仅可以反映种群从出生到死亡的数量动态,还可用于预测种群未来发展的趋势[21]。由表5 可知,种群数量随着径级结构的增加呈现出先增大后减小的趋势,而种群个体存活数lx和标准化存活数lnlx随着径级的增加逐渐减小,从x到x+1 径级间隔期内标准化死亡数dx呈现出先下降后上升的趋势,其dx在Ⅵ径级时出现最大值,为170;种群死亡率qx从Ⅰ径级至Ⅷ径级随演替的进行呈现出先减小后增大的趋势,qx在Ⅵ和Ⅶ两个径级中出现“突变”,其qx明显大于其它径级,其次,qx较高的为Ⅰ径级,说明黔中杜鹃种群在演替过程中幼年个体和老年个体最容易死亡而被淘汰;从x到x+1 径级存活个体数Lx呈现出随龄级的增加而减小的趋势,个体期望寿命ex随着年龄的增加逐渐降低,这与其生物学特性相一致;损失度Tx整体表现为先急剧下降再急剧上升再下降最后剧烈上升的趋势,其损失度Tx在Ⅵ和Ⅶ径级相对较高,其次为Ⅰ径级和Ⅳ径级。

表5 黔中杜鹃种群静态生命表Table 5 Static life table of Rh.feddei population
2.4 黔中杜鹃种群存活曲线特征
存活曲线可以有效地反映种群个体在各年龄级的存活状况[22]。本研究以径级(相对龄级)为横坐标,以lnlx为纵坐标做出了黔中杜鹃种群存活曲线(图2),依据Hett 和Loucks 的数学模型得出的检验方程及参数见表6。由模型检验结果显示,两种模型的拟合结果均达到显著水平,但指数模型的R2值大于幂函数模型,且P值更小,说明黔中杜鹃的存活曲线趋近于Deevey-Ⅱ型。

表6 黔中杜鹃种群存活曲线检验模型Table 6 Test models for survival curve of Rh.feddei population

图2 黔中杜鹃种群存活曲线Figure 2 Survival curve of Rh.feddei population

图3 黔中杜鹃种群死亡率和亏损率曲线Figure 3 Mortality and vanish rate of Rh.feddei population
2.5 死亡率和亏损率曲线
以径级为横坐标,以各龄级的死亡率和亏损率为纵坐标做出的死亡率和亏损率曲线如图3 所示。黔中杜鹃死亡率qx和亏损率Kx曲线变化趋势一致,均呈现出先降低后升高再降低最后急剧增长的趋势(图3),认为黔中杜鹃种群数量具有前期短暂增加后逐渐减少,中期存在短暂的增长,后期急剧减少的特点。
2.6 黔中杜鹃种群生存分析
以径级为横坐标,函数值为纵坐标,绘制了黔中杜鹃野生种群生存率Si、累计死亡率Fi(图4),死亡密度fi和危险率λi函数(图5)。从图4 可以看出,黔中杜鹃种群生存率和累计死亡率呈互补状态,即随着径级的增加,黔中杜鹃种群的生存率下降,累计死亡率逐渐上升;种群在Ⅳ径级达到平衡,即种群生存率和累计死亡率持平。黔中杜鹃死亡密度曲线总体呈现出先缓慢上升后缓慢下降的趋势,整体趋势较为平缓,而危险率曲线总体表现为先缓慢上升后有一个较小的下降趋势,之后急剧上升再急剧下降,波动性比较大,表明黔中杜鹃种群数量受环境影响较大。

图4 黔中杜鹃种群生存率和累计死亡率曲线Figure 4 Survival rate and cumulative mortality rate of Rh.feddei population

图5 黔中杜鹃种群死亡密度和危险率曲线Figure 5 Mortality density and hazard rate of Rh.feddei population
2.7 黔中杜鹃种群数量的时间序列分析
以黔中杜鹃种群各龄级株数为原始数据,按照一次移动平均法则预测出各龄级在未来2,4,6 和8 个龄级时间后的个体数量(表7)。结果显示,经历2,4 个龄级时间,第2至第5 龄级的黔中杜鹃种群个体数均有不同程度的减少,而种群从第6 至第8 龄级开始之后均有不同程度的增加,经过6,8 个龄级时间,种群数量逐渐趋于稳定,说明一旦过了一定的生理年龄,种群就开始逐渐走向衰退,第2 至第3 龄级经过2 个龄级时间后个体数分别下降了40.0%和25.0%,第4至第5 龄级经过4 个龄级时间后个体数分别下降了40.0%和41.2%,这表明黔中杜鹃种群严重缺乏幼龄个体,综合其整个发展趋势来看,黔中杜鹃种群正常更新难以维持,未来会不同程度趋于衰退,甚至濒危。

表7 黔中杜鹃种群动态变化的时间序列分析Table 7 Time sequence analysis of age structure of Rh.feddei population
3 结论与讨论
植物种群结构特征不仅能够体现种群内部个体的发展过程,而且也是植物对环境适应性的反映[23],其径级结构能很好地反应种群动态变化[24]。本研究结果初步表明,黔中杜鹃种群结构整体呈现出不规则金字塔形;种群的中幼龄个体数充足,其个体数量占种群总数量的69.23%;严重缺乏幼龄个体数,仅为种群总数量的11.54%;中老龄个体数量总体呈现出下降趋势;种群目前呈增长型状态。说明黔中杜鹃种群结构总体呈相对稳定型,有向衰退转化的趋势,该种群虽然能够暂时适应当地的生存环境,但由于幼龄个体数的缺乏,随着时间的推移,种群整体的长期稳定性将难以维持。王立龙和易雪梅在裸果木Gymnocarpos przewalskii和水曲柳Fraxinus mandshurica的研究中也有相似的结论[25-26]。
黔中杜鹃种群静态生命表分析表明:黔中杜鹃种群数量随着龄级结构的增加呈现出先增大后减小的趋势,而种群个体存活数(lx)随着年龄的增加逐渐减小,个体期望寿命(ex)随着年龄的增加逐渐降低。这与其生物学特性相一致。黔中杜鹃死亡率(qx)和亏损率(Kx)曲线变化趋势一致,均呈现出先降低后升高再降低最后急剧增长的趋势,表明黔中杜鹃种群数量具有前期短暂增加后逐渐减少,中期存在短暂的增长,后期急剧减少的特点。这与红杉Larix potaninii种群的研究结论相似[27]。
黔中杜鹃种群生存率和累计死亡率呈互补状态,即随着龄级的增加,种群生存率下降,累计死亡率逐渐上升。黔中杜鹃种群在Ⅳ径级达到平衡,即种群生存率和累计死亡率持平;死亡密度曲线总体呈现出先缓慢上升后缓慢下降的趋势,整体趋势较为平缓,而危险率曲线总体表现为先缓慢上升后有一个较小的下降趋势,之后急剧上升再急剧下降,波动性比较大,表明黔中杜鹃种群数量受环境影响较大。姜在民在濒危植物羽叶丁香Syringa pinnatifolia种群的研究中也有相似的结论[28]。
采用时间序列分析中的“一次移动平均法”对黔中杜鹃种群的年龄结构进行预测,结果表明:种群一旦过了一定的生理年龄,就会开始逐渐走向衰退。从黔中杜鹃整个发展趋势来看,该物种种群正常更新难以维持,未来会不同程度趋于衰退,甚至濒危。因此,建议加强黔中杜鹃自生生物学特性研究,进一步摸清其生物学特性,开展种群的更新定位观测研究,包括不同群落枯落物对黔中杜鹃种子的自然萌发和苗木生长的影响,还有黔中杜鹃幼苗建成机制;同时,开展有性与无性繁殖方式相结合,进行回归引种试验,扩大种群数量,提升种群自身抵御种群衰退的风险。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
家教世界·创新阅读(2022年3期)2022-04-12
中老年保健(2021年4期)2021-08-22
心声歌刊(2021年3期)2021-08-05
园艺与种苗(2021年3期)2021-05-26
今日农业(2021年5期)2021-05-22
科学之谜(2020年6期)2020-08-11
小聪仔(幼儿版)(2020年12期)2020-02-01
当代水产(2019年8期)2019-10-12
