
杜仲无性系花芽的抗寒性测定与综合评价
2020-09-28 09:00朱利利刘攀峰杜庆鑫杜红岩
中南林业科技大学学报 2020年9期
何 凤,朱利利,刘攀峰,杜庆鑫,庆 军,王 璐,杜红岩
(1.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;2.南京林业大学,江苏 南京 210037;3.国家林业和草原局 泡桐研究开发中心,河南 郑州 450003;4.经济林种质创新与利用国家林业和草原局重点实验室,河南 郑州 450003)
杜仲Eucommia ulmoidesOliv.属杜仲科植物,该科仅有1属1种,是我国特有的古生树种,可作为优良的药用、用材、水土保持、绿化以及橡胶用树种,具有极大的经济价值[1]。杜仲是我国特有的国家战略储备资源,既是十分重要的优质天然橡胶资源,又是重要的木本油料、名贵中药和国家储备林树种。按照《全国杜仲产业发展规划(2016—2030)》,我国中西部生态脆弱区是今后杜仲资源及产业发展的重点地区,但是杜仲在北移及西部干旱区引种过程中,低温和干旱限制了杜仲栽培区域的扩展。良种选育是杜仲资源开发利用中一项十分重要的基础性工作,和所有栽培植物一样,除产量、品质等经济性状外,新品种的抗逆性是品种评价的重要指标。根据目前杜仲的栽培地域、生态条件和生产用途,客观上要求杜仲新品种应具有较强的抗性。低温作为最常见的制约植物生长发育的环境因素之一,限制了许多野生植物和作物的地理分布,降低了生产率。陈学森等[2]研究得出植物的不同品种遇到相同环境时所遭受到的冻害程度不同,因此栽培生产上选择抗寒性品种是减少冬季冻害的重要方法之一。选育抗寒杜仲品种是促进杜仲产业化发展的必由之路,研究杜仲无性系的抗寒能力对杜仲品种的选育以及在我国北方的推广具有重要的意义。
迄今,国内外学者对杜仲的抗寒性研究不多。杨吉安等[3]通过不同低温处理,用电导法研究了不同低温对杜仲优树无性系的枝条电解质渗出率的影响,并用模糊数学综合评判法分析了10个优树无性系的抗寒性。张博勇等[4]通过不同低温处理,研究了不同低温对杜仲优良无性系枝条电解质渗出率及染色度的影响,并用这2个指标分别对15个杜仲无性系的抗寒性进行了综合评估,结果基本一致。关于其它木本植物抗寒性的研究,大多数做的是枝条抗寒性的研究[5-6]。开花、结实都是由花芽正常发育才能进行的,所以花芽的抗寒性研究还是很有必要的,目前关于花芽抗寒性的研究有些报道[7]。在关于植物抗寒性研究的过程中,渗透调节物质、丙二醛、抗氧化酶等都被用作抗寒性的鉴定指标[8]。本试验研究了杜仲花芽在不同梯度低温胁迫下游离脯氨酸含量、可溶性蛋白含量及SOD、POD、CAT酶活性的变化,并利用隶属函数法、聚类分析对各测定指标进行分析,综合评价杜仲20个无性系花芽抗寒性。通过不同无性系的抗寒性比较,初步判断各个无性系的抗寒性,为杜仲抗寒性品种选育和引种栽培提供理论依据。
1 材料与方法
1.1 试验材料
在试材进入休眠期以后,于2018年12月11日采集种植于河南省新乡市原阳的国家林业和草原局经济林研究开发中心基地杜仲种质资源库20个6年生开花较晚的杜仲雌株无性系为供试树种,编号分别为‘10012C’‘10013C’‘10043C’‘10044C’‘10046C’‘10115C’‘10267C’‘10407C’‘10408C’‘10409C’‘10428C’‘10472C’‘10475C’‘10476C’‘10480C’‘10493C’‘10508C’‘10526C’‘10527C’‘10586C’。每个无性系6株,每株树在东南西北4个方向分别选择3~5个粗度基本一致、无病虫害和木质化良好的1年生枝条,用石蜡封枝条剪口。按照不同无性系编号贴标签,装入自封袋,带回实验室。
1.2 试验设计
各材料均用自来水清洗干净,再用去离子水多次冲洗,用滤纸吸干水分,平均分成6组,将分好的枝条置于0 ℃的低温恒温冰箱中预冷12 h后,取出一组作对照(0 ℃),其余的枝条继续置于低温恒温冰箱中进行低温处理,共5个温度梯度,分别为-10、-15、-25、-30和-40 ℃,降温速度为2 ℃/h,降到所需温度并维持24 h后分别取出1组枝条,将花芽从果枝上拨下来,经液氮速冻后,存于-80 ℃冰箱,用于各项生理指标的测定,各项指标3次重复。
1.3 理化指标的测定
参照李合生等[9]的方法测定生理指标。SOD活性采用NBT光还原法测定,POD活性采用愈创木酚法测定,CAT活性采用紫外吸收法测定,MDA含量采用硫代巴比妥酸(TBA)显色法测定,脯氨酸含量采用酸性茚三酮法测定,可溶性蛋白含量采用考马斯亮蓝G-250法测定。
1.4 数据处理及分析方法
实验数据采用Excel 2016与SPSS 21.0软件进行数据分析、显著性检验和聚类分析,表中所有数据均为平均值±标准差。抗寒性综合评价用隶属函数法综合各项指标进行抗寒性评价,其公式[10]为:Tij=(Xij-Xjmin)/(Xjmax-Xjmin)。式中:Tij表示i品种j指标的抗寒隶属函数值;Xij表示i种类j指标的测定值;Xjmin表示所有品种所有温度梯度下j指标的最小值;Xjmax表示所有品种所有温度梯度下j指标的最大值;i表示某个测试无性系;j表示某项生理指标。假如某项指标与抗寒性呈负相关,则1减去正相关的隶属函数值就是它的隶属函数值,平均每项指标的隶属函数值,确定隶属函数值大小进行排序,从大到小确定杜仲冬芽各个无性系抗寒性的强弱,值越大抗寒性越强,反之抗性越弱。
2 结果与分析
2.1 低温处理下不同无性系的杜仲花芽游离脯氨酸含量的变化
从表1和图1可知,在低温胁迫处理下杜仲花芽游离脯氨酸含量的变化与无性系有关,不同无性系的变化情况有所不同。其中‘10012C’‘10013C’‘10044C’‘10115C’‘10267C’‘10407C’‘10408C’‘10476C’‘10480C’和‘10526C’的游离脯氮酸含量整体上呈“升—降—升”的变化趋势,在-15 ℃和-40 ℃处理时各出现1个峰值,‘10044C’和‘10115C’的游离脯氨酸含量在-15 ℃处理时显著高于其它无性系,在不同温度处理下,‘10115C’的游离脯氨酸含量相对较高,脯氨酸在植物抗冻中具有重要作用,低温胁迫往往伴随着脯氨酸含量的增加,其含量高低与植物抗寒性密切相关,抗寒性强的品种增加的倍数越高[6],说明‘10115C’抗寒性较强;‘10043C’‘10046C’‘10409C’‘10472C’‘10475C’‘10493C’‘10508C’‘10527C’‘10586C’的游离脯氮酸含量均在-10 ℃处理时达到第一个小高峰,但-10~-40 ℃的低温处理时‘10046C’和‘10409C’的游离脯氨酸含量持续降低,‘10475C’在-25~-40 ℃时的游离脯氨酸含量缓慢增加,‘10472C’‘10527C’和‘10586C’在-30 ℃出现第二个高峰,其值大于-10 ℃的高峰,‘10493C’在0、-10和-40 ℃时的游离脯氨酸含量均相对较高,说明其抗寒性较强;‘10428C’的游离脯氨酸含量随着温度的降低变化较平缓。

表1 杜仲无性系花芽在不同低温胁迫下游离脯氨酸含量变化及无性系间差异性分析†Table 1 Changes in free proline content of flower bud of different Eucommia ulmoides clones under low temperature and difference analysis of these clones

图1 人工低温胁迫下不同无性系的杜仲花芽的游离脯氨酸含量变化Fig.1 Variation in free proline content of flower bud of different Eucommia ulmoides clones under artificial temperature stress
2.2 低温处理下不同无性系的杜仲花芽SOD酶活性的变化
从表2和图2可知,整体上,以0 ℃为对照,随着温度的降低,20个杜仲无性系花芽SOD酶活性的变化为“升—降—升”的趋势,在-10 ℃或-15 ℃有一个小高峰,到-40 ℃明显升高达到另一个高峰。‘10012C’‘10013C’‘10046C’‘10115C’‘10267C’‘10407C’‘10408C’‘10409C’‘10475C’‘10476C’和‘10527C’的SOD酶活性在-15 ℃处理时达到第一个小高峰,‘10476C’‘10475C’和‘10115C’的SOD酶活性相对较高(3 863.43、3 837.80、3 399.72 U/g),显著高于其他无性系,-40 ℃处理达到第二个高峰,‘10046C’‘10013C’‘10475’‘10267C’这4个无性系的SOD酶活性相对较高(5 253.68、4 647.79、3 954.24、3 382.31 U/g),通过比较发现‘10475C’在不同低温处理下SOD酶活力一直较高,说明其具有较强的抗寒性;‘10043C’‘10428C’‘10472C’‘10493C’‘10508C’‘10586C’在-10 ℃处理时达到第一个小高峰,‘10493C’和‘10472C’的SOD酶活性相对较高(2 172.66、2 015.56 U/g),-40 ℃达到第二个高峰,‘10428C’‘10586C’‘10508C’的SOD酶活性相对较高(4 357.22、3 356.76、3 088.90 U/g);‘10526C’的SOD酶活性在0~-25 ℃时平缓增加,-25~-40 ℃显著增加,但其值均较低;‘10044C’的SOD酶活性随温度变化出现下降—升高交替的变化趋势,‘10480C’在0~-10 ℃处理时SOD酶活性下降,-10~-40 ℃处理时SOD酶活性缓慢增加,其值较高,说明其抗寒性较高。

表2 杜仲无性系花芽在不同低温胁迫下SOD酶活性变化及无性系间差异性分析Table 2 Changes in SOD activity of flower bud of different Eucommia ulmoides clones under low temperature and difference analysis of these clones

图2 人工低温胁迫下不同无性系的杜仲花芽的SOD活性变化Fig.2 Variation in SOD activty of flower bud of different Eucommia ulmoides clones under artificial temperature stress
2.3 低温处理不同无性系的杜仲花芽POD酶活性变化
从表3和图3可知,在低温胁迫下POD酶活性的变化与无性系有关,不同无性系的变化情况有所不同。‘10012C’‘10043C’‘10046C’‘10407C’‘10480C’和‘10526C’的POD酶活性均在-10 ℃达到第一个峰值,‘10043C’‘10046C’和‘10407C’的POD酶活性随温度变化出现“升—降—升—降”的趋势,在-10 ℃和-25 ℃出现两个高峰,‘10012C’和‘10480C’的POD酶活性随温度变化出现“升—降—升—降”的趋势,在-10 ℃和-30 ℃出现两个高峰,‘10526C’在温度降到-25 ℃后POD酶活性维持在较低水平;‘10267C’‘10115C’‘10408C’‘10475C’‘10508C’的POD酶活性均在-10 ℃出现谷值,在-15 ℃出 现 峰 值;‘10493C’‘10472C’‘10476C’‘10586C’的POD酶活性随温度变化出现“升—降”的变化趋势,在-15 ℃时达到峰值;‘10428C’的POD酶活性随着温度的降低变化波动不大;‘10013C’‘10044C’‘10409C’‘10527C’的POD酶活性在0 ℃到-25 ℃均呈下降趋势,在-25 ℃出现谷值,‘10428C’的POD酶活性一直维持在较低的水平,‘10044C’的POD酶活性随温度的降低一直保持较高水平,说明其抗寒性较强。不同无性系对低温胁迫的反应不同,品种之间的差异很大。
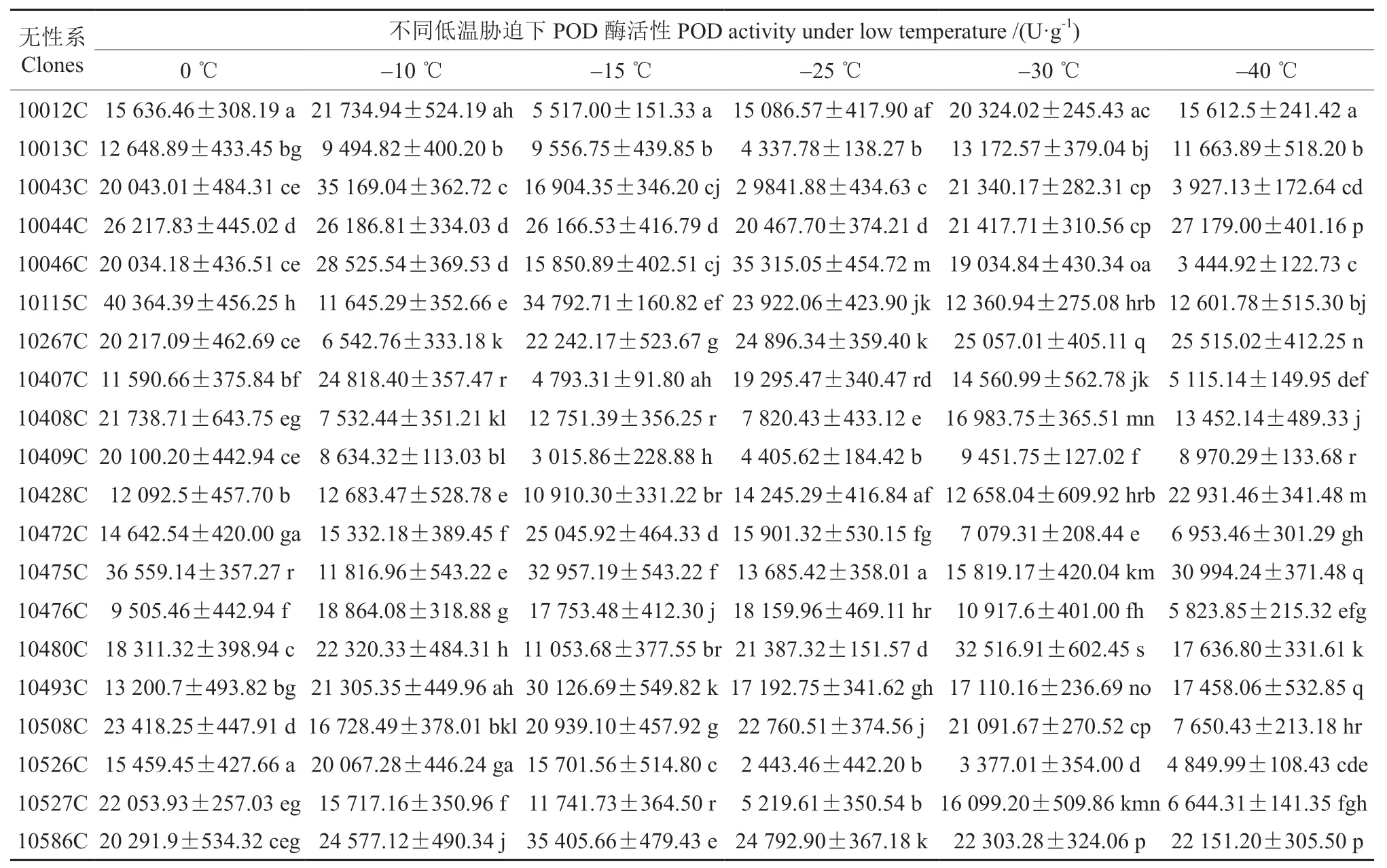
表3 杜仲无性系花芽在不同低温胁迫下POD酶活性变化及无性系间差异性分析Table 3 Changes in POD activity of flower bud of different Eucommia ulmoides clones under low temperature and difference analysis of these clones
2.4 低温处理下不同无性系的杜仲花芽CAT酶活性的变化
从表4和图4可知,在低温胁迫下花芽CAT酶活性的变化与无性系有关,不同无性系的变化情况有所不同。‘10115C’‘10267C’‘10407C’‘10409C’‘10046C’‘10428C’‘10475C’‘10527C’和‘10586C’的CAT酶活性随温度变化呈现“升—降—升—降”的趋势,均在-10 ℃时出现第一个峰值且峰值最大,第二个峰值出现的温度不一致,-25、-30、-40 ℃均有峰值出现;‘10508C’‘10526C’‘10493C’‘10012C’的CAT酶活性随温度变化出现“升—降—升—降”的波状变化,均在-10 ℃和-25 ℃出现两个峰值,后一个峰值高于前一个;‘10013C’和‘10476C’的CAT酶活性随温度变化出现“升—降—升”的变化,在-15 ℃和-40 ℃时出现两个峰值,后一个峰值高于前一个;‘10043C’和‘10044C’的CAT酶活性随温度变化出现“升—降—升—降”的波状变化,在-10 ℃和-30 ℃出现两个峰值,整体来说,0 ℃到-30 ℃,随着温度的下降,CAT酶活性变化比较平稳;‘10408C’‘10472C’和‘10480C’在0 ℃到-10 ℃处理下酶活性先下降,-10 ℃到-25 ℃处理下CAT酶活性变化平缓,‘10472C’和‘10480C’在-30 ℃时CAT酶活性升高,在-40 ℃时下降,‘10408C’在-30 ℃时CAT酶活性下降,在-40 ℃时升高。因此,不同无性系随着温度的变化反应不一致。
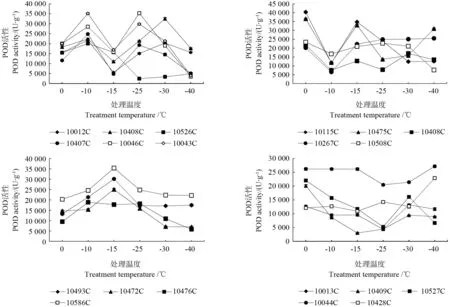
图3 人工低温胁迫下不同无性系的杜仲花芽的POD活性变化Fig.3 Variation in POD activty of flower bud of different Eucommia ulmoides clones under artificial temperature stress

表4 杜仲无性系花芽在不同低温胁迫下CAT酶活性变化及无性系间差异性分析Table 4 Changes in CAT activity of flower bud of different Eucommia ulmoides clones under low temperature and difference analysis of these clones
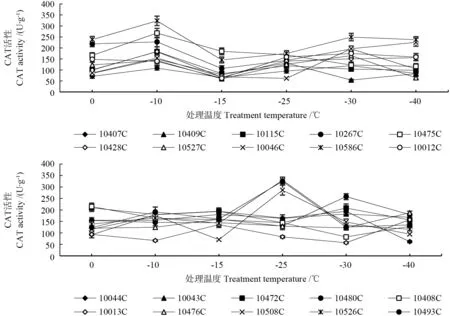
图4 人工低温胁迫下不同无性系的杜仲花芽的CAT活性变化Fig.4 Variation in CAT activty of flower bud of different Eucommia ulmoides clones under artificial temperature stress
2.5 低温处理下不同无性系的杜仲花芽可溶性蛋白含量变化
从表5和图5可知,随着温度的降低,‘10043C’‘10044C’‘10472C’‘10408C’‘10115C‘10267C’‘10428C’‘10475C’‘10493C’‘10508C’‘10526C’和‘10586C’的可溶性蛋白含量整体上出现“升—降—升”的变化趋势,在-10 ℃和-40 ℃时出现峰值,‘10493C’的可溶性蛋白含量整体水平上显著高于其他无性系,说明抗寒性较强;‘10013C’‘10046C’‘10409C’‘10476C’‘10480C’‘10508C’和‘10527C’的可溶性蛋白含量随温度变化出现“降—升—降—升”的变化趋势,在-15 ℃时出现峰值,‘10046C’的可溶性蛋白含量的峰值相对较高,随温度的降低可溶性蛋白含量整体水平上相对较高,说明抗寒性较强;‘10407C’和‘10012C’的可溶性蛋白含量随温度降低持续降低,说明其抗寒性较弱。
2.6 隶属函数法对抗寒性的综合评价
以游离脯氨酸、SOD酶活性、POD酶活性、CAT酶活性、可溶性蛋白含量作为依据,计算各指标的隶属函数值,获得隶属函数值累加起来并取平均值,再比较平均值进行排序,并进行综合评价,结果见表6。抗寒性由大到小排列顺序依次为‘10493C’‘10586C’‘10480C’‘10115C’‘10046C’‘10475C’‘10044C’‘10408C’‘10472C’‘10508C’‘10267C’‘10428C’‘10012C’‘10043C’‘10527C’‘10407C’‘10476C’‘10013C’‘10526C’‘10409C’。
2.7 20个杜仲无性系耐低温胁迫的聚类分析
对20个杜仲无性系耐低温胁迫能力的隶属函数平均值进行聚类分析,将在不同低温胁迫下耐受能力相近的杜仲无性系聚在一起,图6显示,使用Ward聚类法在欧式距离平方值约为11时,可将20种杜仲无性系耐受低温胁迫能力划分为3类,其中‘10493C’和‘10115C’耐受性较强,‘10586C’‘10475C’‘10044C’‘10267C’‘10043C’ 耐受性中等,其他13个杜仲无性系耐受性较弱。

表5 杜仲无性系花芽在不同低温胁迫下可溶性蛋白含量变化及无性系间差异性分析Table 5 Changes in soluble protein content of flower bud of different Eucommia ulmoides clones under low temperature and difference analysis of these clones

图5 人工低温胁迫下不同无性系的杜仲花芽的可溶性蛋白含量变化Fig.5 Variation in soluble protein contents of flower bud of different Eucommia ulmoides clones under artificial temperature stress

表6 不同杜仲无性系抗寒隶属函数值Table 6 Comparative study on cold resistance with membership function value of different Eucommia ulmoides’ clones

图6 20个杜仲无性系耐低温胁迫的聚类分析Fig.6 Cluster analysis of low temperature stress of 20 clones of Eucommia ulmoides
3 结论与讨论
目前国内外有关植物抗寒性的研究较多,主要研究低温胁迫对植物显微结构和超微结构的影响、生理生化指标的变化、抗寒相关基因的研究、外源生长调节剂对植物抗寒性的调控作用以及植物抗寒性鉴定方法[11]。植物对低温的生理反应是受多种因素影响的,其生理变化也很复杂,生理生化指标在反映植物抗寒性指标时的表现和敏感性不同,单独用某一个指标确定植物抗寒性的强弱不具有说服力,多个生理生化指标以及植物实际生长结果结合起来才能综合反映植物的抗寒性[12]。植物在低温胁迫下生存能力差别很大,抗氧化酶系统清除氧自由基和过氧化氢[13],SOD、POD、CAT是清除生物体内活性氧和其他过氧化物自由基的关键酶,在植物抵御低温胁迫过程中发挥着重要作用,其活性大小与植物抗寒性有一定相关性,活性越高植物的抗寒性越强[14-15]。在低温胁迫后期,植物体内抗氧化酶活性减弱,可能由于低温胁迫超过了植物承受极限,酶的结构发生改变或酶的表达受到抑制,从而导致酶活性减弱[11]。研究发现,植物体内抗氧化酶活性与抗寒性具有一定的相关性,抗寒性强的植物抗氧化酶活性较抗寒性弱的植物高[16]。有关研究表明,植物体内SOD活性随着低温胁迫的加剧呈先增强后减弱的趋势[17]。过氧化物酶(POD)是一种氧化还原酶,能够分解植物体内因逆境胁迫而产生的自由基,对逆境胁迫具有一定的防御作用,其活性越高,植物抗逆性越强[18]。脯氨酸和可溶性蛋白是细胞内的主要渗透调节物质,在低温胁迫下渗透调节物质的量增大可缓解或抵御低温伤害的发生,其含量的高低可反映植物抗寒能力的大小[19-20],脯氨酸的变化与植物的抗寒性呈正相关[21]。曲彦婷等[22]对低温胁迫下不同福禄考品种叶片的生理响应进行研究,结果表明随着温度的降低叶片可溶性蛋白含量呈“降—升—降—降”的变化趋势,脯氨酸的变化呈“升—降—降”的趋势,SOD活性的变化呈“降—升—降”的趋势;李嘉诚等[23]对低温胁迫下14个品种平欧杂种榛的1年生休眠枝生理生化的响应进行研究,结果表明不同低温处理下游离脯氨酸呈逐渐上升的趋势;黄永红等人[14]研究了低温胁迫下杏的一年生枝条的生理生化响应,结果表明随着温度的降低游离脯氨酸含量‘红丰’和‘骆驼黄’的变化趋势为“升—降—升”,‘凯特’的变化趋势为“降—升”,SOD比活力变化趋势为“升—降—升—降”,POD活性和CAT活性的总体变化趋势为“升—降—升”,可溶性蛋白含量的变化趋势为“降—升—降”,与本试验中部分无性系游离脯氨酸、SOD、POD、CAT、可溶性蛋白的变化趋势研究结果一致;欧欢等人[24]对低温胁迫下不同品种扁桃花蕾的生理指标的变化进行研究,结果表明在不同低温胁迫下不同品种扁桃花蕾可溶性蛋白的含量变化不同,浓帕烈、苦巴旦、小软壳花蕾可溶性蛋白质含量呈先上升后下降的趋势,桃巴旦和晚丰花蕾可溶性蛋白质含量呈“升—降—升”的趋势,与本试验中部分无性系可溶性蛋白含量的变化趋势一致。
抗寒性是由多因素联合控制的综合性数量性状,依据单一生理指标难以准确全面地反映植物抗寒性的强弱[25],在许多研究中均利用了隶属函数法进行多指标综合评价[26-29]。根据杜仲花芽抗寒隶属函数值,各无性系按照抗寒性由强到弱排 序 依 次 是‘10493C’‘10586C’‘10480C’‘10115C’‘10046C’‘10475C’‘10044C’‘10408C’‘10472C’‘10508C’‘10267C’‘10428C’‘10012C’‘10043C’‘10527C’‘10407C’‘10476C’‘10013C’‘10526C’‘10409C’。通过聚类分析发现‘10493C’和‘10115C’耐受性较强,‘10493C’和‘10115C’的接穗来源于北方,因此试验结果有一定的可信度,‘10493C’和‘10115C’可供将来在寒冷地区引种中参考使用。在自然条件下,植物受到低温胁迫时也会受到其他胁迫的影响。目前,主要采用人工模拟气候室法对植物离体叶片或枝条等进行低温胁迫处理,然后根据各项生理生化指标的变化综合评价植物抗寒性,但未必能够真实反映出个别植物在自然生长环境下的抗寒性。本研究试验材料采用的是离体果枝上的花芽,受到树龄、树体长势、果枝上花芽发育状况等的影响,且采用的室内人工低温恒温冰箱模拟外界环境的方法不可能完全正确地反映出自然环境条件下的冻害程度,均会造成试验上的误差[30]。因此,下一步可从这20个无性系中挑选几个抗寒性较强的无性系种植在寒冷地区,观察其成活率才能更好地分析其抗寒的能力;建议在以后的试验中对整株植物进行处理,同时还要结合野外试验和其他逆境环境进行综合研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
热带作物学报(2022年5期)2022-06-01
安徽化工(2021年3期)2021-05-29
山西农业科学(2020年10期)2020-10-16
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
陕西画报(2016年1期)2016-12-01
小雪花·小学生快乐作文(2015年9期)2015-09-29
小雪花·小学生快乐作文(2015年9期)2015-09-29
小雪花·小学生快乐作文(2015年9期)2015-09-29
