
新疆野生梯牧草种质资源分布与保护利用
2020-09-22 04:04:56张鲜花朱进忠李江艳
新疆农业科学 2020年8期
张鲜花,朱进忠,李江艳
(新疆农业大学草业与环境科学学院,新疆草地资源与生态重点实验室, 乌鲁木齐 830052)
0 引 言
【研究意义】梯牧草 (Phleum.pratenseL.),又名猫尾草,系禾本科梯牧草属植物。梯牧草属野生种全世界约有15种,主要分布在N40°~50°寒冷潮湿地区,我国有4种,新疆均有分布。与其近缘种有鬼蜡烛(P.paniculatumL.)、高山梯牧草(P.alpinumL.)和假梯牧草(P.phleoides(L.) Karst.),在天然草地中皆为优良牧草[1-3]。梯牧草饲用价值高、抗逆性强、适应性广,是奶牛和赛马的良好饲草[4]。在我国东北、华北和西北地区均有栽培。国外,在欧洲、北美洲一些国家广泛栽培,以美国和原苏联栽培面积最大[5]。据资料记载[1],梯牧草在我国的分布仅限于新疆天山山地,在集中分布地段的山地草甸草地中,常以伴生种存在,是天然草地的重要饲用植物。研究新疆野生梯牧草的种质地理资源分布及群落结构学,对牧草新品种选育、种质创新、种质保护与利用最有重要意义。【前人研究进展】于磊等[6]研究表明,在准噶尔西部山地只有梯牧草的近缘种假梯牧草的分布。 近年来,在梯牧草的引种选育也出了一些品种[7-11],如1934~1935年新疆就从原苏联引进梯牧草种子,分别在伊犁地区和乌鲁木齐试种,迄今还有少量逸生后代[10]。1941年中国甘肃从美国引进梯牧草在峨山县种畜场种植,选育出了眠山梯牧草。1998年中国四川草原总站从原西德引进了梯牧草,四川草原研究所从国内外引种了10多个梯牧草品种,经过20多年的自然选择,发现其残存植株及逸生后代对高寒牧区环境有很好的适应性,且存在着许多变异类型,其中有一些是很好的早熟、晚熟材料[11]。在自然界生物种群的分布可以大致分为3类,即均匀性(uniform)、随机型(random)和聚集型(clumped)[8]。【本研究切入点】有关新疆野生梯牧草种质资源分布的文献较少。研究野生梯牧草在新疆的地理分布及群落特征学。【拟解决的关键问题】以分布于新疆境内的野生梯牧草为对象,研究其种质资源的地理分布、种群分布的群落学特征以及开发利用保护,为该资源的开发利用及保护提供参考。
1 材料与方法
1.1 材 料
调查样地取自于新疆天山西段伊犁哈萨克自治州昭苏县和天山中段的乌鲁木齐县。天山是世界上最大呈东西走向的独立山系。东西延绵1 700 km。地处80°~96°E,42°10′~45°30′N,占地57×104km2,平均海拔约5 000 m,最高峰托木尔峰达7 435.3 m。天山在我国境内自西向东分为三段,即伊犁哈萨克自治州至乌苏县称为西段,平均海拔3 000~5 000 m;乌苏县至乌鲁木齐县称为天山中段,平均海拔高度为4 000~4 300 m;乌鲁木齐县至哈密地区为天山东段,平均海拔3 000~4 000 m。天山山地是新疆重要的山地草场分布区,草地垂直带谱完整,植物种类繁多,是重要的草牧业生产基地。表1
1.2 方 法
1.2.1 调查项目
野外调查采用区域控制与辅助路线调查相结合的方法进行。从梯牧草种群的地理分布、分布生境、个体分布空间格局、群落种类组成,分析梯牧草在野生状态下的植物地理学、种群与群落学特征。
种群地理分布:调查与查清梯牧草在新疆境内的分布。
群落种类组成与数量特征:调查梯牧草群落植物种类科、属、种组成,测定群落物种高度、盖度,分析梯牧草群落的数量特征变化与生境的关系。
种群个体分布空间格局:观察分析不同地段梯牧草种群空间分布特征,阐明组成种群个体在其生活空间中的位置状态与布局。
1.2.2 测定指标
植物种类组成:在调查样地内,统计群落植物科、属、种组成。
数量特征测定:测定群落植物的高度、盖度。
高度:测定群落内所有植物的自然生长高度,计算平均值。
盖度:用目测法进行。
2 结果与分析
2.1 新疆野生梯牧草种质资源的分布
2.1.1 种群的地理分布
研究表明,梯牧草在天然草地中的分布,具有严格的地理区域限制。《中国植物志》和《新疆植物志》记载,梯牧草在新疆分布仅限于天山与准噶尔西部山地特定的气候带和植被区域。2018~2019年连续两年对该区域进行考察,梯牧草在新疆只分布于天山山地。
梯牧草在新疆仅分布于天山山地的中、西地段,天山的东段山地从吉木萨尔至木垒一线,山地草甸广为发育,许多中生禾草繁衍茂盛,但草地群落中未曾发现有梯牧草的分布。
在天山中段山地,梯牧草只出现于位于乌鲁木齐县海拔为1 900~2 100 m的山地草甸植被中,多以伴生种出现在草地群落中,分布范围与面积也十分有限。研究表明,处于天山西段的伊犁地区的昭苏县是野生梯牧草的集中分布区,以位于县城北部的乌孙山分布最多,其次为沙尔套山,在海拔1 700~2 200 m的不同地形部位与水热条件下,均可觅到它们的踪迹,并以优势种和伴生种出现在不同的草地类型中。在属于天山山系的阿拉套山,在山地海拔2 200~2 400 m山地草甸植被中,梯牧草以伴生种常出现在草地群落中。
梯牧草的分布与其他中生禾草相比,对区域自然条件要求更高,决定了在草地中分布域的局限性。表2

表2 不同地段梯牧草集中分布区及海拔Table 2 Concentrated distribution areas and altitudes of Phleum pretense in different sections
2.1.2 种群分布的生境特征
研究表明,梯牧草在种的集中分布区,分布生境也十分局限。在天然草地中,多生长于局部地区曾有定期积水的低洼地和紧邻河谷阶地,空旷的山坡地虽有分布,但与山地的中小地形和聚集了最佳的水、热条件的生境有一定联系,如在坡地中经降水冲刷形成的小沟谷,修筑道路取土形成的低洼地和路基两侧,常形成种群的聚集分布,而在平直的坡面很少有生长。
梯牧草的分布对水、热条件有较高要求,尤其是对水分因子的耐受范围较窄,与同一分布地带内其他中生禾本科植物如鸭茅(Dactylisglomerata)、无芒雀麦(Bromusinermis)、早熟禾(Poasp)对水分条件响应的生态幅要窄的多,水分是梯牧草广域分布的主要限制因素,其窄水性是梯牧草种群繁衍与扩散对生境响应的最基本特征。
2.1.3 种群个体分布的空间格局
研究表明,梯牧草在天然草地中的分布,主要以聚集型和随机型2种形式存在。由于梯牧草在群落中数量上的劣势和对生境适应的局限性,在草地上中分布常带有一定的随机性,个体间的出现完全与机会符合,种群的个体依靠种子在母株周边的撒播而成随机分布。在开阔的山坡地,这种分布类型较为普遍,种群个体出现在很大程度上带有一定的偶然性,从而决定了梯牧草种群个体分布空间格局与生境的关系。
聚集型分布是调查区域内梯牧草种群分布格局最多的一种形式。其分布与局地或某一地段小生境以及梯牧草的繁殖特性有较大关系。该种分布类型在调查区域内仅出现在山地坡地和沟谷的一些低洼处,受降水汇集的作用,其生境较周边地区湿润,梯牧草以绝对优势的地位,形成局部聚集分布片段,但种群繁殖增长与扩散具有严格的生境边界,形成有特定的生态格局。
2.2 新疆野生梯牧草分布的群落类型及主要特征
虽然梯牧草在天然草地中的分布数量有限,但所处形成条件的差异和局部生境条件的变化呈现出一定的多样性。依据群落分布的地形部位,有坡地群落类型,山间谷地群落类型和河流阶地群落类型;分布类型不同,群落植物种类组成、数量特征,种间关系,种群个体空间分布格局以及在群落中的数量与地位,植株个体植物学性状均表现出一定的差异性。表3,表4
2.2.1 山坡地群落类型
是分布区域内群落分布的主导类型,占据山地半阴坡或半阳坡和台地,在草地群落中梯牧草多以伴生种与一些禾草、杂类草形成群落。
在伊犁昭苏县的乌孙山西段山地海2 000~2 200 m地段,梯牧草以伴生种与其他草类相伴存在形成的种类组成丰富、草群密度大、草层高、生物量积累大的群落。群落中常见禾草有鸭茅、无芒雀麦、长芒大穗鹅观草(Roegneriaaboliniisubsp.divaricans)、曲芒鹅观草(R.tschimganica)、圆柱披碱草(Elemus.Dahuricussubsp.cylindricus)、草地早熟禾(Poapratensis)、细叶早熟禾(P.angustifolia)、偃麦草(Elytrigiarepens)、新疆鹅观草(R.sinkiangensis)、细弱剪股颖(Agrostistenuis)等。其他草类有红花车轴草(Trifoliumpratense)、新疆鼠尾草(Salvia.deserta)、牛至(Origanumvulgare)、天山千里光(Seneciotianschanicus)、森林勿忘草(Myosotissilvatica)、穗花婆婆纳(Veronicaspicata)、黄花委陵菜(Potentilla.chrysantha)、蓬子菜(Galiumverum)、北方拉拉藤(G.boreale)、森林草莓(Fragaria.vesca)、直立老鹳草(Geraniumrectum)、丘陵老鹳草(G.collium)、新疆唐松草(Thalictrum.collinum)、千叶蓍(Achilleamillefolium)、线叶野豌豆(Viciatenuifolia)、聚花风铃草(Campanulaglomerata)等,草群平均高度40~140 cm,禾本科牧草生殖枝高度可达100~140 cm,梯牧草高度60.1~132.1 cm;群落覆盖度80%~90%,其中梯牧草的盖度差异极大,一般在5%~10%,局部水分聚集的低洼处可达到的60%,形成以梯牧草为建群种的群落。
在中段山地的乌鲁木齐南山,梯牧草主要分布在海拔1 900~2 100 m地段的坡地与台地,群落种类组成,鸭茅与无芒雀麦、披碱草、草地早熟禾、小糠草(Agrostisgigantea)、偃麦草形成群落的上层层片;红豆草(O.vicifolia)与千叶蓍、草原糙苏(Phlomispratensis)、森林草莓、牛蒡(Arctiumlappa)、黄花委陵菜(P.chrysantha)、和兰芹(Carumcarvi)、青兰(D.ruyschiana)、箭头唐松草(T.simplx)、直立老鹳草(G.rectum)、蓬子菜(Galiumverum)、聚花风铃草、白花车轴草(Trifoliumrepens)等杂类草形成下层层片,草群平草群平均高度40~120 cm,梯牧草高度89.1~115.2 cm;群落覆盖度85%~90%,其中梯牧草的盖度在10%~15%。
2.2.2 沟谷地群落类型
该类型是指分布于山地宽阔沟谷的群落,多占据梯牧草种群集中分布区的低海拔地段。因地形效应,在同海拔地段,土壤中较坡地具有较多的水分,土层深厚肥沃,是梯牧草生长的理想生境地。在草地中梯牧草以优势种出现在群落中,形成由梯牧草与其他禾草和杂类草组成的群落。梯牧草在群落中个体数量多,盖度大,生长茂盛,生物量大,对整个群落具有控制性影响,对草地群落与生态系统的稳定起着举足轻重的作用。群落种类组成,梯牧草与鸭茅、披碱草、小糠草、亚洲异燕麦(Helictotrichonhookeri)形成群落的上层层片;红车轴草(T.pratense)与千叶蓍、黄花委陵菜、、黄芪(Astragalussp)等杂类草形成下层层片,草群平均高度40~110 cm,梯牧草平均高度为55~70 cm,高者可达110~120 cm;群落盖度90%~100%,梯牧草在群落中的盖度可达到60%,占绝对优势。
2.2.3 近河流阶地群落类型
该类型是在具有河流地段与乔灌草混生的林间空地镶嵌分布的一种类型。生境荫蔽而湿润,在围栏条件下,草群高大,各种植物生长茂盛。由于受河流与小气候的影响,梯牧草的分布在海拔梯度上具有较宽的生态幅,但在草地中均以偶见种出现在群落中。草地群落由鸭茅、小糠草、披碱草(E.dahuricus)、偃麦草(Elytrigiarepens)、新疆鹅观草、千叶蓍、蒲公英(Taraxacumaltaicum)、唐松草、荷兰芹、红车轴草、车前(Plantagosp)、黄花萎陵菜、直立老鹳草等组成。草群高40~140 cm,梯牧草高度120~140 cm;群落盖度80%~90%,梯牧草盖度仅占1%左右。
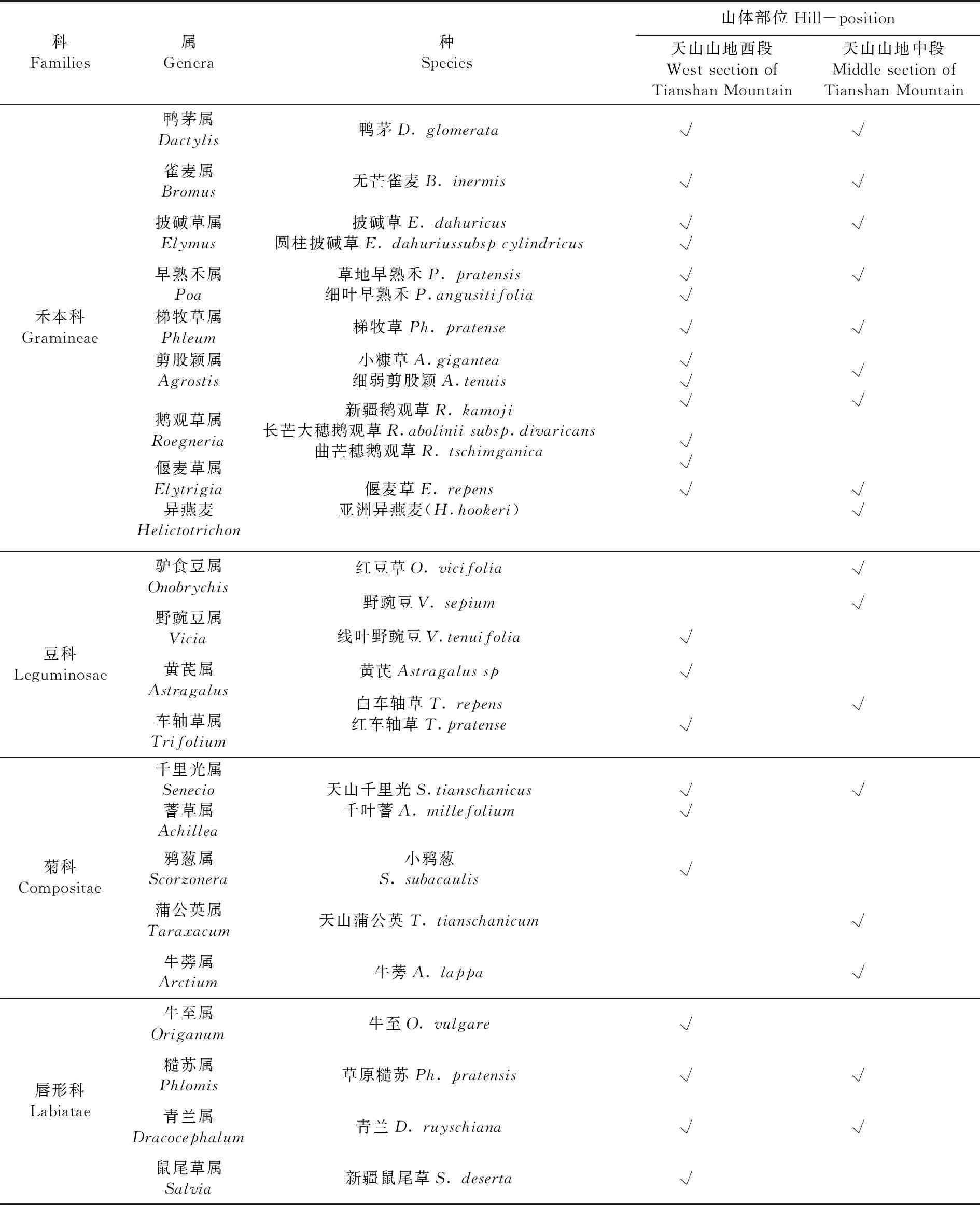
表3 不同地段梯牧草群落植物种类组成Table 3 The species composition of Phleum pretense in different locations

科Families属Genera种 Species山体部位Hill-position天山山地西段West section of Tianshan Mountain天山山地中段Middle section of Tianshan Mountain蔷薇科Rosaceae委陵菜属Potentilla黄花委陵菜P. chrysantha√√草莓属Fragaria森林草莓F. vesca√√地榆属Sanguisorbs地榆S. officinalis√√伞形科Umbelliferae和兰芹属Carum和兰芹C. carvi√√厚棱芹属Pachypleurum厚棱芹P.mucronatum√毛茛科Ranunculaceae唐松草属Thalictrum新疆唐松草T. collinum箭头唐松草T.simplx√√√翠雀属Delphinium伊犁翠雀花D. iliense√玄参科Scrophulariacea马先蒿属Pedicularis鼻花马先蒿P. rhinanthoides√婆婆纳属Veronica 穗花婆婆纳V. spicata√√牦牛儿苗科Geraniaceae老鹳草属Geranium直立老鹳草G. rectum丘陵老鹳草G. collium√√√茜草科Rubiaceae茜草属Rubia茜草R. cordifolia√拉拉藤属Galium蓬子菜G. verum北方拉拉藤G. boreale√√√车前科Plantaginaceae车前属Plantago平车前P. depressa√桔梗科Campanulaceae风铃草属Campanula聚花风铃草C. glomerata√√石竹科Caryophyllaceae蝇子草属Silene昭苏蝇子草S. holopetala√√紫草科Boraginaceae勿忘草属Myosotis森林勿忘草M. silvatica√√蓼科Polygonaceae酸模属Rumex皱叶酸模R. crispus√√合计:15科37属49种15科32属40种14科30属31种
3 讨 论
3.1 目前多数工作是从引进品种的研发做起,对国内野生资源的研究所做工作甚少,尤其是针对野生梯牧草种质资源的一些基础性研究尚处于空白[12-16]。一是对野生种质资源的地理分布开展研究,明确种间多样性的地理空间分布,阐明多样性形成与地理空间分布的关联性,探讨种质资源多样性及其形成机制;二是要对种质的植株形态学特征进行研究,通过对不同生育时期植株营养器官与繁殖器官的形态学特征分析,确定优良种质的表型特点;三是对所蕴藏的基因多样性与优良基因的发掘,种内和种间是否存在遗传变异、基因交流和居群分化,在表型特征、染色体数目及其结构、基因位点、DNA序列等存怎样的差异,通过研究揭示种质资源中的优异基因。
3.2 要加大种质资源的搜集与开发利用的工作力度,在对种质资源的地理分布和遗传多样性等研究基础上,着力发掘种质资源中的优异基因,开展优质牧草品种创新研究。通过广泛收集种质资
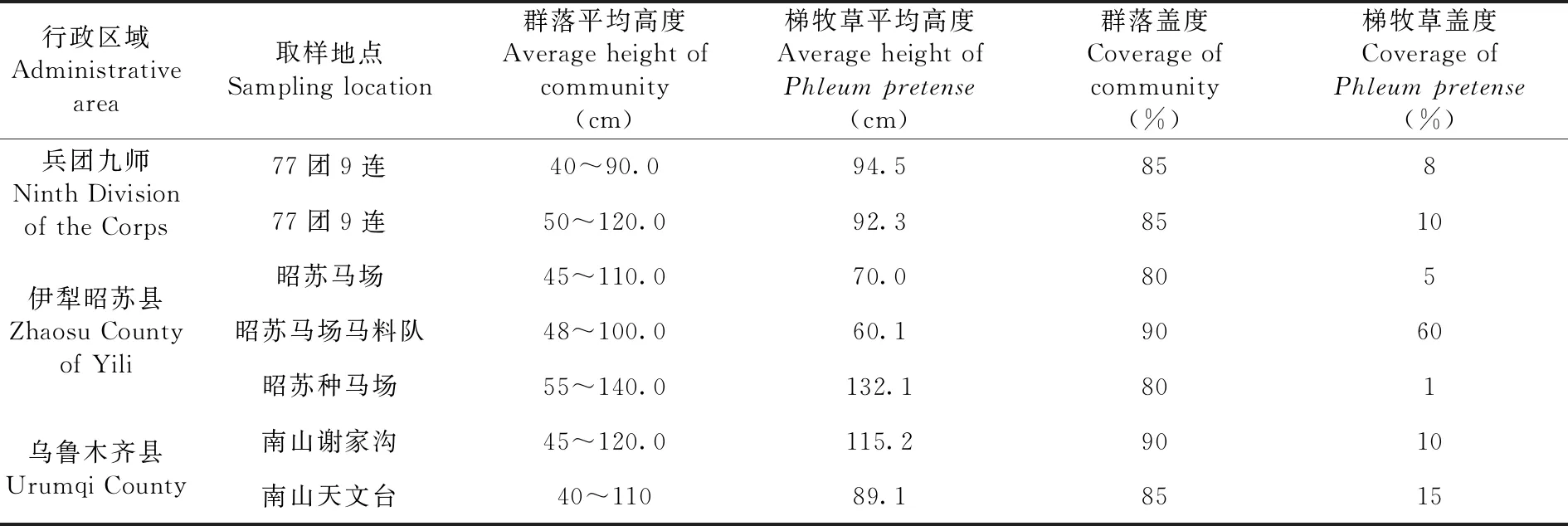
表4 不同地段梯牧草群落的高度、盖度Table 4 The height and coverage of Phleum pretense in different locations
源,围绕优异种质的评价与筛选从中筛选出具有开发利用价值的优良种质,明确优良性状的特点,揭示优良性状表达的形态学、生物学意义,探讨优异种资利用价值。研究农艺性状特性,主要围绕物候期、生长发育速度、再生性能、分蘖能力、牧草与种子产量及营养价值等进行观察与测定,揭示种质资源开发利用的方式与潜力,明确优异种质的表现及农艺价值。
3.3 调查表明,新疆野生梯牧草种质资源正在面临着濒临灭绝的边缘,目前种群分布的区域正在逐年缩小,据新疆植物志记载,梯牧草在乌鲁木齐、石河子、和布克赛尔、塔城、尼勒克、新源、昭苏与特克斯均有分布。通过近2年的调查,许多地段已难觅其踪迹,野生梯牧草的分布只局限在伊犁哈萨克自治州的昭苏县和乌鲁木齐县,然而在这一区域也仅出现在具有围栏的割草草地中,分布面积十分有限。
3.4 野生梯牧草种群的分布有严格的地理生态区域,新疆分布区只局限于天山东、西段山地的部分草甸植被中,形成为该地区的特有种。草地植物地理分布受到环境影响,环境对植物类群的选择发展进化与地理分布起有选择作用。
《新疆植物志》记载,梯牧草在该地区的新源县、尼勒克县也有分布,但此次研究结果在两县的草地中均未发现有梯牧草踪迹,这一结果是否受调查路线的局限影响,尚需进一步拓宽调查面加以证实。梯牧草种群的分布格局类型除在部分地段为随机分布外,大多情况下均呈聚集分布。此结果再一次证明了以往的观点,即自然界中大多数种群均呈聚集分布。有研究表明,在环境均匀一致的条件下,植物种群格局是随机的和均匀的,而不同的环境条件下,却往往是聚集型的。一般来说,种群空间分布格局不仅因种而异,甚至同一物种的分布格局在不同的年龄结构及不同的生境情况下也有差异,作为群落中的优势种,会决定着种群的分布格局,如刘青松等[17]、刘振国等[18]的研究结果也证明了这一点。在研究中,梯牧草种群的分布是严格受生境的制约,中小地形决定着种群的存在与分布类型。由此可见,种群在空间上的分布格局主要是与环境长期相互适应的结果。
4 结 论
4.1 梯牧草在天然草地中的分布,受严格的地理区域限制。在我国仅分布于新疆天山山地的中、西段海拔1 900~2 200 m山地草甸植被中。
4.2 梯牧草在草地群落中常以伴生种和偶见种出现,少有以优势种出现的种群分布。
4.3 梯牧草种群个体空间分布格局,多数地段呈聚集分布,随机分布形式较少。
4.4 新疆野生梯牧草具有重要的开发利用价值。
4.5 新疆野生梯牧草种质资源已处于濒危阶段,种群的分布仅限于面积十分有限的围栏草地内。
猜你喜欢
心声歌刊(2023年5期)2023-10-10 13:31:30
纺织学报(2022年8期)2022-08-26 02:29:32
落叶果树(2021年6期)2021-02-12 01:29:10
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
水土保持研究(2019年6期)2019-10-19 03:33:32
民族音乐(2016年4期)2016-12-06 02:50:38
体育科技(2016年2期)2016-02-28 17:06:07
防护林科技(2015年5期)2015-06-10 11:42:52
海南热带海洋学院学报(2015年5期)2015-03-14 09:51:07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
