
川西高原昆虫病原真菌的多样性*
2020-09-16 04:55:02裴晓亚MadukaNilakshiJayasekaraArachchige朱晨慧
林业科学 2020年8期
裴晓亚 Maduka Nilakshi Jayasekara Arachchige 朱晨慧 王 敦
(西北农林科技大学昆虫学研究所 杨凌 712100)
川西高原是四川省西部与青海、西藏交界的高海拔区,地势跌宕起伏,气候复杂多变,地域差异显著,孕育着丰富的生物种类,其中昆虫和真菌资源十分丰富。全球昆虫病原真菌种类丰富,迄今已记录800多种虫生真菌,我国已发现400余种(蒲蛰龙等,1996)。昆虫病原真菌是重要的生物防治资源,同时也可以稳定生态系统中昆虫种群。利用昆虫病原真菌防治害虫,对人、动物无毒害作用,对环境友好,因此在植物保护研究中受到广泛关注(王滨等, 2005; 刘春来, 2017; 李增智, 2015)。目前,物种多样性海拔分布格局及其形成机制是多样性研究的一个重要课题(徐翔等,2018),虫生真菌的分布格局与海拔区域也有密切关系(李亦菲等, 2018)。
本文利用Hiseq技术对川西高原13个海拔样地的土壤样品进行测序,探讨川西高原的昆虫病原真菌种类资源、分布以及虫生真菌多样性沿海拔梯度的变化规律,分析海拔变化对昆虫病原真菌多样性的影响,为昆虫病原真菌的开发利用以及川西高原物种多样性的研究提供科学依据。
1 材料与方法
1.1 试验材料
按照“S”形取样法,在雅安(多营镇、天全县、二郎山)、甘孜(雅江、新龙、沪定)和康定(折多山、新都桥)8个地域内共13个不同海拔区域进行随机取样。为避免土壤空间分布不均所导致的误差,在每个海拔分别选取5个取样点,每2个样点之间间隔大于50 km。土壤样品采集后立即封口,-10 ℃冻存于车载冰箱中,并用GPS记录坐标、贴标签。实验室内将每个海拔的5个样品混合,倒在灭菌后的纸上,用镊子去除样品中的植物根系、昆虫、石块等杂物,放于无菌台内风干、研磨,过1~2 mm筛后随机分为3份,保存于-80 ℃冰箱中,作为土壤微生物 DNA 提取的3个生物学重复。各土壤样品信息如表1所示。

表1 土壤样品信息
1.2 试验方法
土壤总DNA的提取参考并改进Bürgmann等(2001)的方法,经过物理研磨、化学试剂萃取获得总DNA。土壤总DNA的纯化参考购自BioFlux公司DNA纯化试剂盒(Gel DNA Purification Kit)的操作说明进行。
分别用13个土壤样品的DNA为模板,扩增引物参考White(1990)方法,分别为ITS1(5’TCCGTAGGTGAACCTGCGG3’),ITS4(5’TCCTCC GCTTATTGATATGC3’),扩增体系为25 μL(10×Buffer 2.5 μL、d NTP 2 μL、上下引物各1 μL、模板0.3 μL、dd H2O 18.7 μL),产物进行凝胶电泳处理、胶回收试剂盒对DNA进行回收,使用illumina Hiseq进行测序。
1.3 数据处理与分析
测序得到的原始数据,经过拼接、过滤、去除嵌合体成为有效数据,对其进行后续生物信息分析: 采用OTU(operational taxonomic units)即操作分类单元[为了便于进行分析,给某一个操作分类单元(品系、种、属、分组等)设置同一标志,即97%的相似度进行OTU聚类], 结合微生物目前最权威的数据库——Unite数据库进行物种注释和物种分类分析。 基于OTU的分析,获得样品的丰度信息,挖掘样品之间的群落结构和物种组成差异。
采用香侬指数(Shannon)Hshannon、辛普森指数(Simpson)D、Chao1指数(OTU估计数SChao1及覆盖率Coverage进行生物多样性分析。计算公式如下:
Schao1=Sobs+n1(n1-1)/[2(n2+1)],
coverage=[1-(ni/N)]×100%。
式中,Sobs为OTU实测数;ni为含有i条序列的OTU数目,N为全部序列数;Pi为ni/N。
2 结果与分析
2.1 测序数据预处理及OTU数统计
对13个海拔样品的测序结果表明,平均每个样品得到85 009条有效序列; 碱基质量值Q30比例均大于84.34%,满足后期分析的要求。按照序列的相似性高于97%定义为一个OTU,共获得51 515个OTU,其中13个样品共有的OTU个数为50个,样品DY、TQ、LD、ER、ZDS、Y1、Y2、Y3、XD、Y4、XL、Y5、Y6特有的OTU个数分别为228、1 179、863、682、1 346、1 466、1 556、316、1 540、724、790、1 880、1 127。
2.2 土壤中真菌的α多样性分析
13个土壤样品中真菌的α多样性如表2所示。所有样品的Coverage均超过94%,说明测序深度合理,可以反应样本的真实情况,其中,香侬指数在5.14~9.27之间, 辛普森指数在0.77~0.99之间, Chao1指数在2 172~5 608之间。

表2 土壤中真菌多样性指数统计结果
2.3 土壤中真菌等生物种类
对13个样品51 515个OTU的分析表明,在门水平上,相对丰度排名前十位的分别为子囊菌门(Ascomycota)、担子菌门(Bassidiomycota)、接合菌门(Zygomycota)、纤毛门(Ciliophora)、壶菌门(Chytridiomycota)、绿藻门(Chlorophyta)、罗兹菌门(Rozellomycota)、丝足虫类(Cercozoa)、球囊菌门(Glomeromycota)和芽枝霉门(Blastocladiomycota)(图1)。其中,子囊菌门与担子菌门在每个样品中的共同占比超过69%,占绝对优势。除样品Dy之外,剩余12个样本的优势门均为子囊菌门,占比均超过49%,次优势门为担子菌门,占比均超过6%。在样品Dy中,优势菌门为担子菌门,占比为62%,次优势菌门为子囊菌门,占比13.7%。排名前十位的门,除罗兹菌门与芽枝霉门外,有8个门在13个样品中均有分布。

图1 土壤真菌在门水平的分布
从属水平的分布来看(图2),13个样品间的差异比较显著,相对丰度排名前十位的分别是Echinoderma、丝膜菌属(Cortinarius)、青霉属(Penicillium)、被孢霉属(Mortierella)、棒束孢属(Isaria)、镰刀菌属(Fusarium)、假裸囊菌属(Pseudogymnoascus)、赤霉菌属(Gibberella)隐球菌属(Cryptococcus)、小菇属(Mycena)。13个样本中最优势属均有差异,在样品DY中,Echinoderma的占比为47.6%,占绝对优势; 样品Y3中的最优势属为丝膜菌属,占比24%; 其他样本中各最优势属均有差异。除Echinoderma、棒束孢属与假裸囊菌属之外,其他7个属在13个样本中均有分布。

图2 土壤真菌在属水平的分布
2.4 土壤中昆虫病原真菌的种类
13个不同海拔样本中的昆虫病原真菌主要分布于子囊菌门肉座菌目(Hypocreales)的麦角菌科(Clavicipitaceae)、虫草科(Cordycipitaceae)和线虫草科(Ophiocordycipitaceae)。这3个科的OTU约占总OTU的2.08%,而每个科的占比在各个采样点差距很大(图3)。对于麦角菌科,样品ZDS和DY的OTU占肉座菌目的6%,在13个样品中最高; Y4在13个样品中最低,只有0.08%; 13个样品平均占肉座菌目OTU的3%。对于虫草科,Y4的OTU在肉座菌目中的占比在13个样品最高,为 78%; 在XD中只占0.5%; 平均占肉座菌目的21%。 对于线虫草科,13个样品的OTU在这个科中的占比是3个科中最低的,平均值只有1%,其中Y3占比最低,只有0.05%; Dy和Y6最高。
在13个样品中检测到的昆虫病原真菌分属于子囊菌门肉座菌目3个科中的18个属。其中:麦角菌科7个属,虫草科6个属,线虫草科5个属;绿僵菌属(Metarhizium)、普可尼亚属(Pochonia)以及弯颈霉属(Tolypocladium)在全部样品中都被检测到; 棒束孢属、虫草属(Cordyceps)以及线虫草属(Ophiocordyceps)在12个样品中被检测到; 淡紫紫霉属(Purpureocillum)在9个样本中被检测到; 余下11个属都分别只在1~4个样本中被检测到。如表3所示,13个样品在属水平上差异很大,Y1中属的种类最多,共有11个,DY中属的种类最少,只有4个,分别为绿僵菌属、普可尼亚属、Rotiferophthora、弯颈霉属。
在麦角菌科中,绿僵菌属在13个样品中全都被检测到,除样品Y5之外,在余下样本中的占比均超过60%,占绝对优势; 在虫草科中,除样品DY之外,棒束孢属和虫草属在余下的12个样品中全部被检测到,且共同占比均超过68%,占绝对优势,但不同的样本中,最优势属存在差异; 线虫草科中弯颈霉属在全部13个样本中都被检测到,在样品DY、TQ、ER、ZDS、XD、Y5中都为优势属,淡紫紫霉属在9个样品中均为检测出,且在样品LD、Y1、Y4、Y5中均为优势属,在样品Y3中,弯颈霉属与线虫草属的占比均为50%,同为优势属。

图3 不同样品OTU数量占肉座菌目OTU数量百分比

表3 虫生真菌各属OTUs占同一样品本科所有OTUs的比例
2.5 虫生真菌多样性沿海拔梯度的变化
13个样品的采样地点均在川西高原,DY、TQ、LD、ER、ZDS、Y1、Y2、Y3、XD、Y4、XL、Y5、Y6海拔依次升高(500~4 300 m),跨度达3 800 m。通过OTU分析子囊菌门肉座菌目3个科中的优势属的OTU占比(图4)发现:麦角菌科的普可尼亚属随海拔的升高没有明显变化,绿僵菌属在500~1 000 m随海拔升高而降低,在2 100 m处达到峰值,然后随海拔的升高而下降; 在虫草科中,棒束孢属和虫草属随海拔高度的变化存在差异,但是这种变化并不是呈梯度递增或是递减,在500~2 100 m的范围内变化不大,基本在1%左右浮动,2 800 m之后数值开始大幅度变化,2 800~3 400 m依次递增,2个属分别在3 100 m与3 400 m处达到第1个顶峰,之后在3 471 m处达到一个低值,在3 700 m处又突然增高到达第2个顶峰,也是棒束孢属的最高值,3 700 m后逐渐减少,而虫草属在4 300 m处达到第3个峰值,且为最高值。由此推断,海拔会影响虫生真菌的多样性,特别是虫草科,其优势属OTU在肉座菌目OTU的占比与海拔相关,在4 300 m采样点虫草属占比最大,在3 700 m采样点棒束孢属的占比最大,而在2 100 m以下虫草科真菌种类极少。海拔3 471 m处虫草科的2个优势属均处于2个峰值(3 400、3 700 m)占比之间,其比值很低,可能与其他环境因子有关,也可能是测序样本误差导致,具体原因还需进一步研究。线虫草科的优势属弯颈属与淡紫紫霉属随海拔的变化其变化差异不大,基本在1%以下。
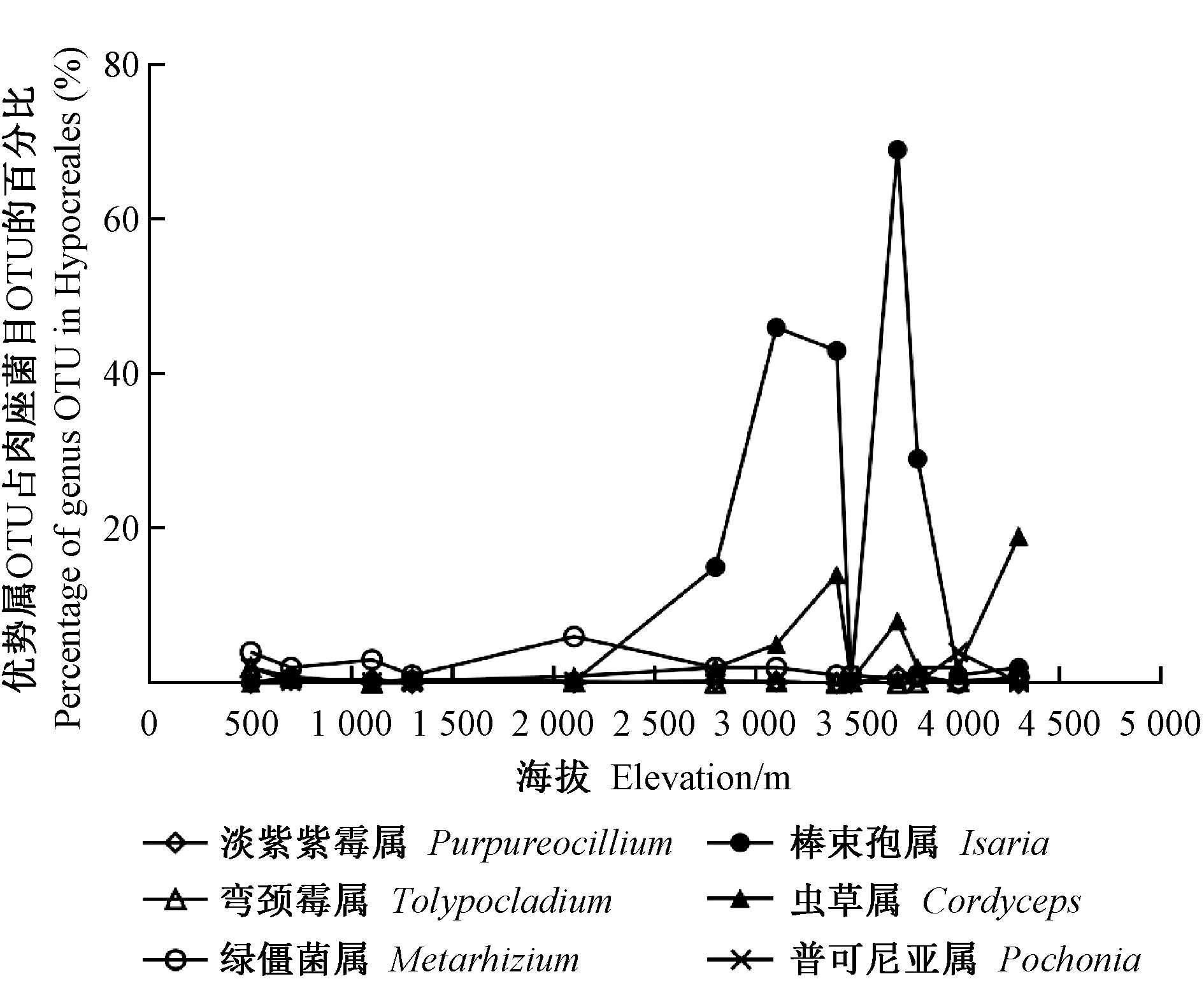
图4 真菌优势属与海拔的相关性
3 讨论
丰富的物种资源对维持生态系统平衡起着至关重要的作用,土壤功能的保持同样需要多种微生物资源来平衡(雷霆等, 2007)。本文通过对川西高原的土壤样品进行测序分析,发现昆虫病原真菌共有18个属,表明川西高原昆虫病原真菌群落丰富,其昆虫病原真菌类型数量高于安徽紫蓬山国家森林公园(刘玉军等, 2018),安徽琅琊山、鹞落坪、麻姑山(陈名君, 2008)和山西庞泉沟自然保护区(李文英等, 2003a)等等。
昆虫病原真菌存在明显地域分布特点,其种类丰富程度与环境有很大的关系(李文英等, 2003b),包括海拔、植被以及地理位置等都与昆虫病原真菌的种类数量与组成结构存在显著差异(陈名君, 2008)。本文的分析结果表明,13个样本的昆虫病原真菌种类及分布都存在差异,检测到的虫生真菌主要存在于肉座菌目的3个科中,其中各科的优势属的分布均有差异,棒束孢属和虫草属作为虫草科的优势属,在除样本DY以外都有分布,且在海拔超过2 100 m时相对丰度升高,表明其适合在高海拔地区生长; 白僵菌作为1种重要的虫生真菌,在此次取样点中鲜有分布,而在其他地区昆虫病原真菌的调查中常常为优势属种(周家喜等, 2017; 王四宝等, 2004),但这些地区的海拔均低于2 000 m,而川西高原海拔远远高于2 000 m。绿僵菌属的相对丰度虽在2 100 m处有峰值,但总体表现为随海拔的升高而下降,与Masoudi等(2018)的研究结果一致; 虫草科在2 100 m以上地区相对丰度较大,适合在高海拔地区生长,可能是因为气候条件适合且高海拔地区人为因素的影响相对较小,具体原因有待进一步研究。
对川西高原的昆虫病原真菌多样性的研究有助于昆虫病原真菌的开发与利用,也可为川西高原物种多样性的研究提供基础数据。
4 结论
本研究对川西高原的13个不同地点的土壤真菌样品进行测序,分析其病原微生物的种类及分布。门水平上,子囊菌门与担子菌门在每个样品中的共同占比超过69%,占据绝对优势。从属水平分布来看,不同样品的优势属存在差异,说明13个样品的虫生真菌种类存在差异。昆虫病原真菌主要分布于子囊菌门肉座菌目的3个科。虫草属与棒束孢属的相对丰度随海拔的升高,分别在4 300与3 700 m达到峰值; 而绿僵菌属随海拔的升高其相对丰度降低,说明相对于虫草菌科真菌,低海拔地区绿僵菌属真菌丰富度较高。
猜你喜欢
中老年保健(2022年2期)2022-08-24 03:20:44
散文诗(2021年22期)2022-01-12 06:14:04
中老年保健(2021年12期)2021-11-30 02:58:01
环球时报(2019-05-23)2019-05-23 06:17:36
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
成都信息工程大学学报(2017年6期)2017-03-16 03:04:39
高原山地气象研究(2016年1期)2016-11-10 06:06:15
中央民族大学学报(自然科学版)(2016年3期)2016-06-27 07:55:28
西南农业学报(2016年5期)2016-05-17 05:42:22
河北遥感(2014年4期)2014-07-10 13:54:58
